
黄芪甲苷对宫颈癌细胞株放射敏感性的影响及相关机制
2018-12-17 11:56周艳艳宋晓婕邵艳社
中国中医基础医学杂志 2018年11期
李 潇,周艳艳,宋晓婕,李 岩,邵艳社
(河南中医药大学第二附属医院, 郑州 450002)
宫颈癌在所有女性肿瘤发病病例数中排名第三位,死亡率位居女性肿瘤的第四位,约有超过85%的患者在发展中国家[1]。大量临床资料表明,早婚早育、性生活紊乱、宫颈创伤以及激素失调等均可增加宫颈癌的患病率[2]。近年来,宫颈癌患者具有年轻化趋势。放射治疗是宫颈癌临床治疗的主要手段之一,NCCN指南2012版推荐首选同步放化疗[3]。由于放疗的副反应会使患者承受较大痛苦,甚至会引起正常组织并发症,为避免此等损伤需降低肿瘤照射剂量,降低肿瘤的有效治疗剂量。
黄芪甲苷(Astragaloside IV)是天然药物黄芪的主要活性成分。已有多项研究表明,黄芪甲苷具有多种药理活性、免疫调节作用、器官保护作用、降糖作用、抗细胞凋亡作用以及抗炎抗病毒作用等[4]。尚未有文献关于黄芪甲苷对宫颈癌细胞的放射敏感性作用的研究。本文采用体外实验,观察黄芪甲苷对Hela细胞株放射敏感性的影响,并探讨其可能的作用机制。
1 材料与方法
1.1 实验材料
宫颈癌细胞株Hela购自上海科学院细胞中心;胎牛血清、胰蛋白酶和DMEM培养基购自Hyclone公司;MTT试剂购自美国Amresco公司;黄芪甲苷(Astragaloside IV)购自中国药品生物制品鉴定所;青霉素、链霉素购自华北制药股份有限公司;MTT试剂、碘化丙啶(PI)购自上海生工生物工程有限公司;p-EGFR、EGFR与p-Akt、Akt抗体购自美国Santa Cruz公司;其他试剂购自国产分析纯。
1.2 细胞培养与照射方法
将Hela细胞株培养于含10%胎牛血清中,青霉素与链霉素各100 U/ml的DMEM完全培养基中,置于37 ℃ 5% CO2的培养箱中培养。收集处于对数生长期的细胞0.25%胰酶消化,调整细胞密度为5×106个/mL,置于96孔培养板中。
1.3 MTT法检测黄芪甲苷对Hela细胞的生长抑制作用
收集处于对数生长期的Hela细胞,计数后调整细胞浓度为1×105个/mL,接种于96孔培养板中,培养贴壁后更换为加入含以下浓度的黄芪甲苷培养液:5、10、20、40、60、80和100 μg/mL,置于37 ℃5%CO2的培养箱中培养24 h,加入50 μL MTT溶液,继续培养4 h,每孔加入DMSO 150 μL,溶解结晶物,于570 nm处检测吸收值(OD值),并计算细胞生长抑制率,每个浓度设3复孔,同时设空白对照组。细胞生长抑制率=1-(药物组OD值-空白对照组OD值)/(对照组OD值-空白对照组OD值)×100%。
1.4 克隆形成实验
实验分为2组,放射组(6 Gy)仅给予照射处理,联合组(Ast-IV+ 6 Gy)给予60 μg/mL黄芪甲苷和不同照射处理,用剂量为0、2、4、6、8 Gy射线照射细胞,收集处于对数生长期的Hela细胞,并制成单细胞悬液。照射方法:将培养板置于照射野(20 cm×20 cm)内,距射野边缘2.5 cm,用直线加速器6MV-X线照射,源皮距(SSD)100 cm,剂量率300 cGy/min。联合组在照射前先给予黄芪甲苷培养24 h后再接受X射线照射,24 h后更换10%胎牛血清的PRMI-1640培养基,继续培养14 d。PBS液洗涤,用甲醛固定、吉姆萨液染色,自然晾干后克隆计数。多靶单击型模型公式:SF=l-(l-e-D/D0)N勾画细胞存活曲线,其中SF为细胞存活分数,D为放射剂量(Gy),D0代表平均致死剂量,Dq代表准域剂量,N为外推数,lnN=Dq/D0。并计算黄芪甲苷的放射增敏比(SER):SER=D0放射/D0放射+药物。
1.5 流式细胞仪检测细胞周期
收集处于对数生长期的Hela细胞,并制成细胞悬液,调整细胞浓度为2×104个/mL。实验分空白对照组(control组)、单药组(Ast-IV组)、放射组(6Gy)和联合组(Ast-IV+ 6Gy)(给予黄芪甲苷培养24 h后再进行照射处理),单药组给予60 μg/ml 黄芪甲苷,照射组与联合组的照射剂量为6 Gy。置于37 ℃5%CO2的培养箱中培养24 h,收集细胞并调整浓度为1×106个/mL。离心1000 r/pm,10 min,PBS洗涤、重悬,加入4 μL碘化丙啶(40 μg/mL),流式细胞仪观察细胞周期变化。
1.6 Tunel检测细胞凋亡
将细胞接种于已灭菌的培养皿中,1×105个/孔,待细胞贴壁后加入黄芪甲苷使最终浓度为60 μg/mL,培养24 h。4%多聚甲醛 4 ℃固定 30 min,0.1%Triton X-100冰浴2 min,分别加入50 μL TUNEL检测液,继续置于37 ℃避光孵育 60 min,PBS洗涤,加入DAPI染液(10 mg/L)孵育15 min,抗荧光淬灭剂封片,采用共聚焦激光扫描显微镜观察细胞的凋亡情况,并统计细胞凋亡率。
1.7 免疫印迹法
收集经相应处理的细胞,提取总蛋白并测定蛋白浓度,每道加入50 μg蛋白进行SDS-PAGE电泳,电转至PVDF膜,加入脱脂牛奶封闭1 h,加入一抗孵育1 h或者过夜,4 ℃,TBST液洗涤3次,加入相对应的二抗室温孵育1 h,TBST液洗涤3次,化学发光法显色,成像扫描分析系统保存图像。
1.8 统计学方法
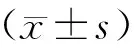
2 结果
2.1 黄芪甲苷抑制Hela细胞的生长作用
表1显示,采用MTT法检测Hela细胞在不同浓度黄芪甲苷处理下存活率的变化。 Hela细胞的存活率随着黄芪甲苷浓度增加而下降(P<0.05),当黄芪甲苷浓度为60 μg/ml时,随之黄芪甲苷的浓度增大,Hela细胞的存活率变化不明显(P>0.05),因此后续实验采用黄芪甲苷浓度为60 μg/ml进行试验。
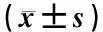
表1 不同浓度黄芪甲苷对Hela细胞生长的抑制作用
注:随着黄芪甲苷浓度增加,细胞存活率逐渐下降,当黄芪甲苷浓度为60 μg/ml时,继续增加黄芪甲苷浓度,细胞存活率无显著变化,与60 μg/mL组比较:#P>0.05;与0 μg/mL组比较:*P<0.05
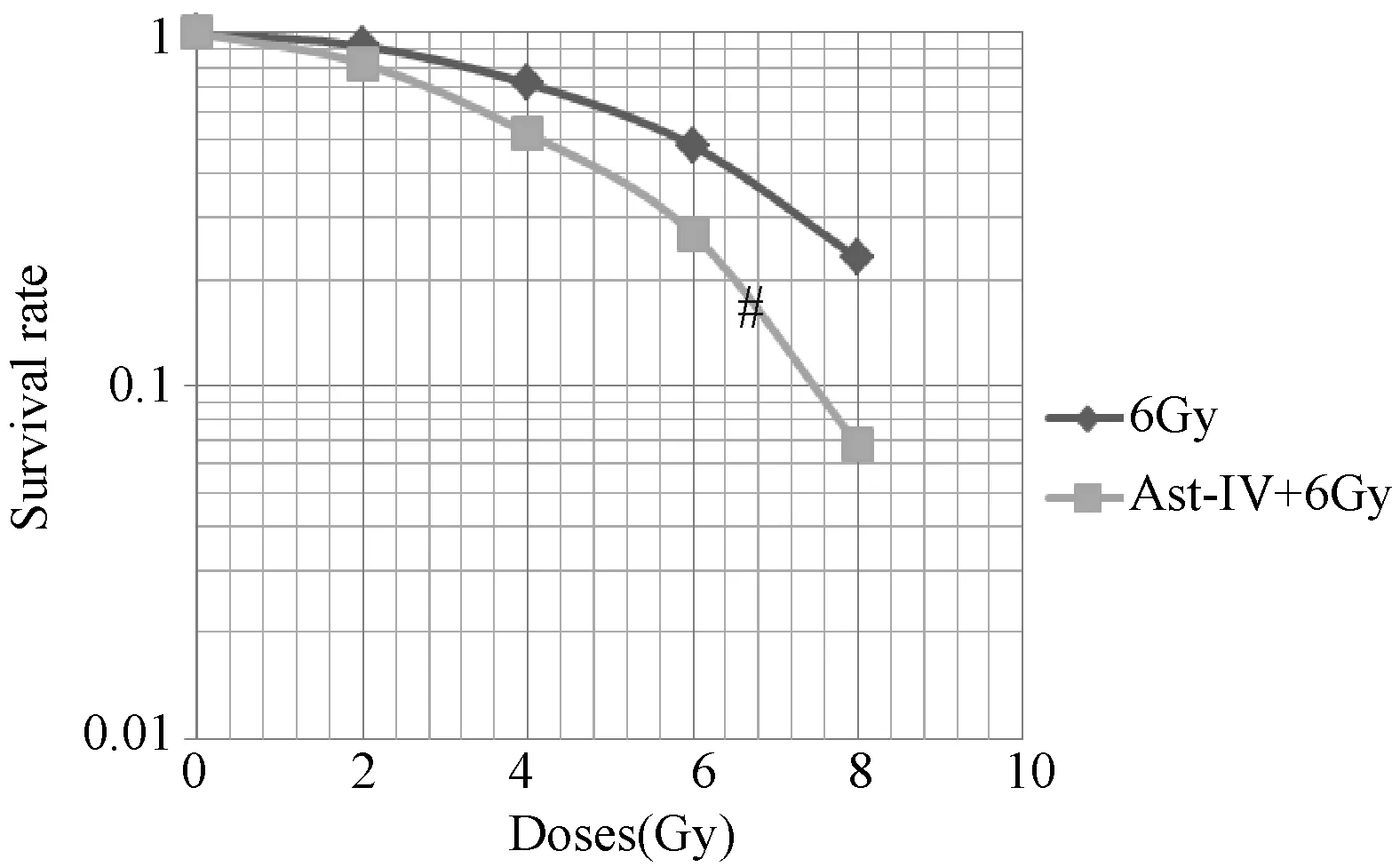
注:黄芪甲苷联合放射处理后,细胞的存活曲线左移; 与6Gy放射组比较:#P<0.05图1 黄芪甲苷对Hela细胞的放射增敏作用
2.2 黄芪甲苷对Hela细胞的放射增敏作用
表2图1显示,Hela细胞经过60 μg/mL黄芪甲苷处理后,采用不同剂量的γ射线照射,计算放射生物学参数和存活分数,拟合细胞存活曲线,在各照射剂量下,联合组的存活分数低于放射组(P<0.05);与放射组比较,联合组的整体生存曲线左移。
表2 黄芪甲苷对Hela细胞的放射生物学参数
注:与照射组的相同放射生物学参数比较:#P<0.05
2.3 黄芪甲苷对Hela细胞周期的影响
图2表3显示,与空白对照组比较,其余各组的细胞均出现不同程度的G2/M期阻滞。其中,联合组细胞的G2/M期所占比例显著低于单药组和照射组(P<0.05),提示黄芪甲苷可降低Hela细胞放射诱导的G2/M期阻滞。
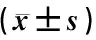
表3 黄芪甲苷对Hela细胞周期G2/M期的影响
注:与control组比较:*P<0.05;与6Gy组比较:#P<0.05
2.4 黄芪甲苷对Hela细胞凋亡的影响
图3表4显示,Tunel实验结果显示,采用60 μg/mL黄芪甲苷培养细胞24 h后,联合组细胞凋亡数显著高于空白对照组与单药组(P<0.05),其中正常对照组和溶剂对照组之间的细胞凋亡比较差异无统计学意义(P>0.05)。

图2 黄芪甲苷对Hela细胞周期G2/M期的影响
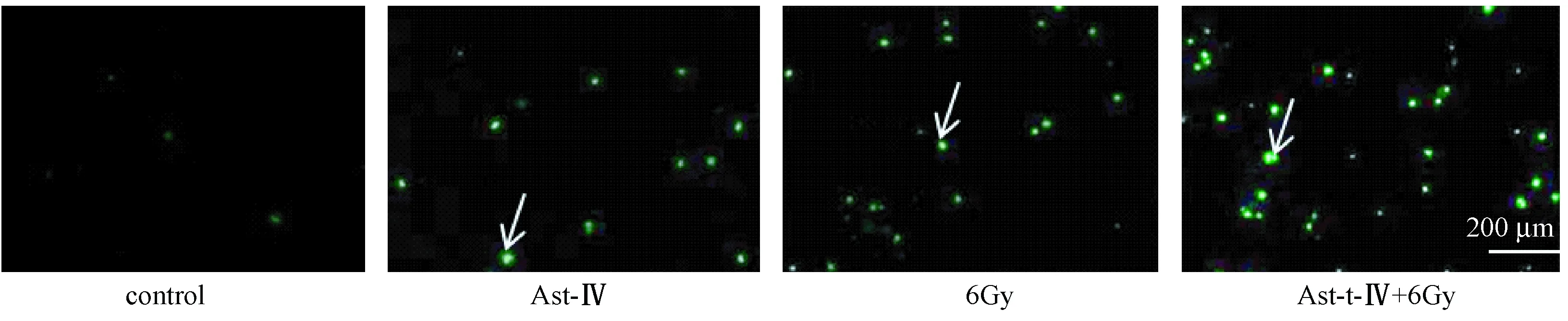
注:图中箭头指向为凋亡细胞图3 黄芪甲苷对Hela细胞凋亡的影响(×200)
2.5 黄芪甲苷对Hela细胞中p-EGFR、EGFR与p-Akt、Akt的蛋白表达影响
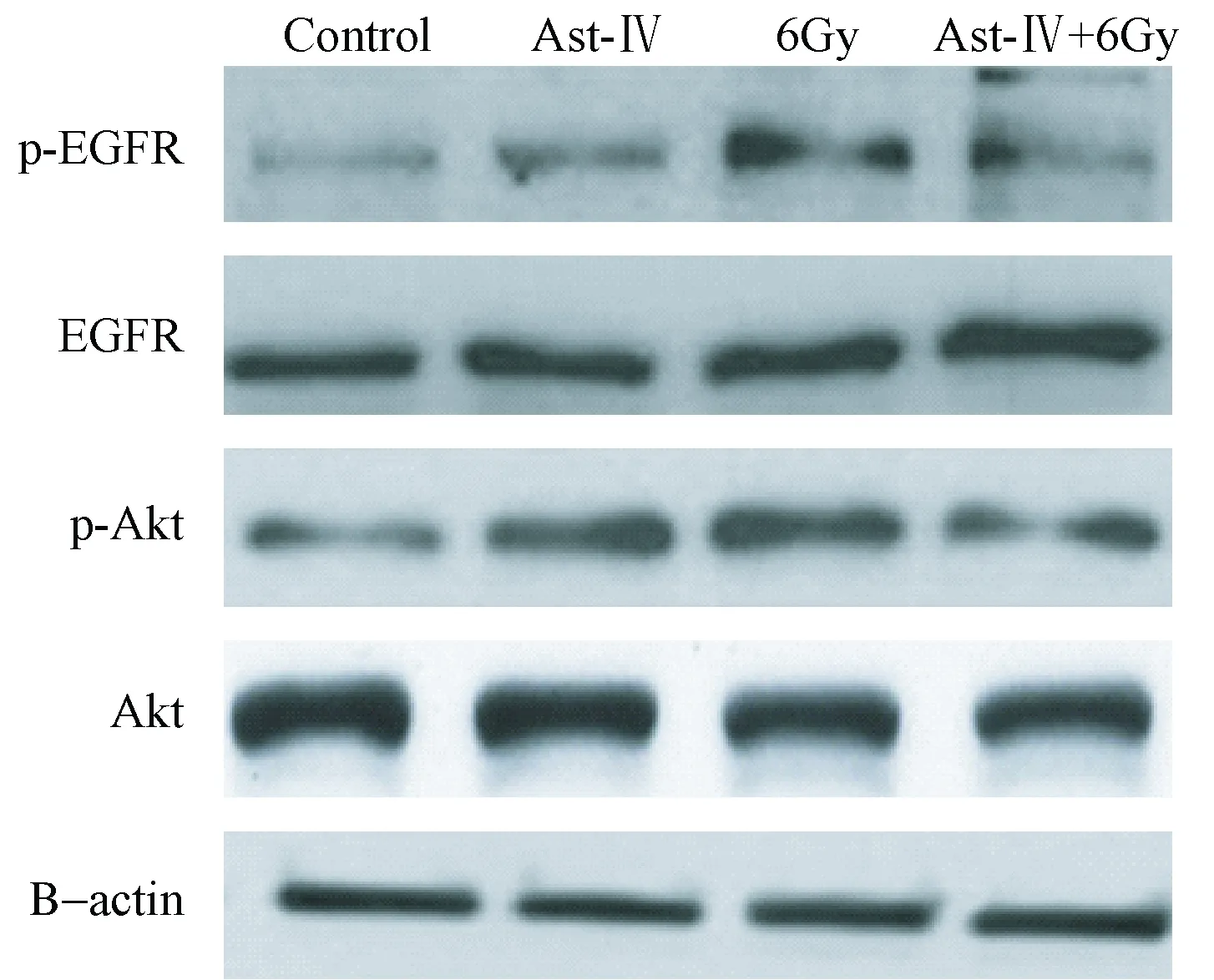
图4 黄芪甲苷对Hela细胞中p-EGFR、EGFR与 p-Akt、Akt蛋白表达的影响
图4表5显示,照射组细胞中p-EGFR与p-Akt的蛋白表达明显高于空白对照组和单药组,联合组细胞中p-EGFR与p-Akt的蛋白表达显著低于照射组和单药组(P<0.05),各组细胞中EGFR与Akt比较差异无统计学意义(P>0.05)。
表4 黄芪甲苷对Hela细胞凋亡的影响
注:与control组比较:*P<0.05
表5 黄芪甲苷对Hela细胞中p-EGFR,EGFR与p-Akt、Akt的蛋白表达影响
注:与control组比较:*P<0.05;与6Gy组比较:#P<0.05
3 讨论
从MTT试验结果可知,当黄芪甲苷浓度达到60 μg/ml后,随着药物浓度增大,Hela细胞的存活率变化不明显,因此在后续实验中黄芪甲苷的浓度采用60 μg/ml,提示黄芪甲苷具有抑制宫颈癌细胞Hela增殖的作用。通过多靶单击模型绘制放射存活曲线可见,在各个照射剂量下,Ast-IV+ 6 Gy组的存活分数均低于6 Gy组,提示黄芪甲苷具有增强X射线对Hela细胞的杀伤作用;与6 Gy组比较,Ast-IV+ 6 Gy组的曲线向左下方移,Dq值减小,肩区减少,提示黄芪甲苷对Hela细胞的亚致死性损伤修复能力减弱,D0值减少,提示黄芪甲苷具有增加Hela细胞X射线敏感性的作用。
多项实验显示,细胞的DNA损伤修复能力是细胞照射敏感性的主要决定因素之一[5],当辐射产生的高活性化学自由基造成细胞内DNA损伤,双链断裂。另一个重要决定因素是细胞周期调控,大量实验数据表明,细胞的放射敏感性随细胞周期时相的变化而改变。肿瘤细胞受到放射引起DNA损伤后,会导致细胞周期阻滞与G1/S期或G2/M期,进行DNA损伤的修复[6],未能成功修复则启动凋亡信号使细胞产生凋亡[7]。细胞周期不同阶段对放射敏感性不同,其中以G2/M期最为敏感,出现G2/M期阻滞,一方面细胞无法增殖分化,另一方面DNA损伤修复也会被阻止[8]。本实验发现,单独射线照射诱导Hela细胞的G2/M期阻滞,采用黄芪甲苷干预后显著缩短放射诱导Hela细胞的G2/M期所占比例,降低阻滞程度,从而增加Hela细胞的放射敏感性。
表皮生长因子受体(EGFR)具有酪氨酸激酶活性,是调节肿瘤细胞分化和生长的重要信号传导通路之一。当其过表达或异常活化时,与促进肿瘤形成且参与放射诱导的肿瘤细胞损伤修复机制密切相关[9]。研究表明,放射线直接或间接作用于DNA,导致DNA损伤,通过DNA双键断裂诱导细胞凋亡。EGFR活化后从胞浆进入胞核,与DNA-PK结合,催化DNA-PKcs磷酸化,直接调节放射诱导的DNA损伤修复。研究发现,X射线照射显著引起肝癌细胞中的PI3K磷酸化,进一步引起Akt发生磷酸化;黄芪甲苷联合照射处理后,Hela细胞的EGFR与Akt磷酸化水平低于单纯照射的细胞,提示黄芪甲苷能够抑制X射线照射引起的Akt磷酸化,抑制PI3K/Akt信号通路的活化,从而提高Hela细胞的放射敏感性。进一步推测黄芪甲苷可能通过抑制内源化配体与EGFR结合,进一步阻断EGFR介导的PI3K/Akt信号通路,干扰DNA损伤修复,抑制肿瘤细胞增殖,降低放射抗拒性,增加对Hela细胞放射增敏的作用。更具体的黄芪甲苷增加Hela细胞放射敏感性的深入机制,正在进行更严谨的实验中。本实验对黄芪甲苷用于宫颈癌的临床研究提供了实验基础。黄芪是常用的传统中草药,容易获得且可以广泛推广使用。
猜你喜欢
新农业(2022年21期)2022-11-18
中日友好医院学报(2022年4期)2022-10-15
皮肤性病诊疗学杂志(2021年5期)2021-11-27
山东医药(2021年3期)2021-02-24
三农资讯半月报(2021年1期)2021-01-27
看世界·学术下半月(2020年7期)2020-09-10
安徽中医药大学学报(2020年2期)2020-04-21
中国老年学杂志(2019年20期)2019-10-17
中成药(2017年9期)2017-12-19
中成药(2017年7期)2017-11-22
