
不同地点的黄秋葵部分数量性状表达差异
2019-01-09 07:09章毅颖黄志城赵洪徐丽李寿国高玲陈海荣任丽
热带作物学报 2019年12期
关键词:黄秋葵
章毅颖 黄志城 赵洪 徐丽 李寿国 高玲 陈海荣 任丽
摘 要 以16个黄秋葵品种为材料,通过种植试验探究不同生态区的环境条件差异对黄秋葵品种数量性状的影响。结果表明:叶片长度、宽度在品种及地区间数据波动较大,因此这2个性状受品种及环境的互作效应较大;叶柄长在品种间差异较小,但受环境影响较大;果实厚度和果实长度在品种内容易出现不稳定的情况,而果实长度和果实直径在品种间的平均变异系数相对较小,分别为9.68%和8.68%,因此环境作用对其影响相对较小;单荚种子数在不同生态区表现出显著差异,易受环境影响;在品种类型中,矮杆类型、长果类型、红果类型的黄秋葵相较于其他类型更易受到环境的影响。
关键词 种植地点;栽培环境;黄秋葵;数量性状
中图分类号 Q949.757.3 文献标识码 A
The Expression Difference of Some Quantitative Characteristics of Okra (Abelmoschus esculentus) in Different Regions
ZHANG Yiying1,2, HUANG Zhicheng1,2, ZHAO Hong1,2, XU Li3, LI Shouguo2, GAO Ling3, CHEN Hairong1,2, REN Li1,2*
1. Institute for Agri-food Standards and Testing Technology, Shanghai Academy of Agricultural Sciences, Shanghai 201403, China; 2. Shanghai Sub-center for Plant New Variety Tests, Ministry of Agriculture and Rural Affairs, Shanghai 201415, China; 3. Tropical Crops Genetic Resources Institute, Chinese Academy of Tropical Agricultural Sciences / Danzhou DUS Testing Station of New Variety of Plants, Ministry of Agriculture and Rural Affairs, Danzhou, Hainan 571737, China
Abstract In order to investigate the effects of environmental conditions on the quantitative traits in okra, 16 okra varieties were used as the materials in different sowing sites. The results showed that the leaf length and leaf width fluctuated greatly among varieties and sites. Therefore, the two traits were more affected by the variety and environment. The petiole length was less varied among varieties, but it was greatly affected by the environment. Fruit thickness and fruit length were prone to instability among the varieties, 9.68% and 8.68% respectively, while the average coefficient variation (CV) of fruit length and fruit diameter among varieties was less significant. The number of seeds per pod had significant differences in different sowing sites, so it was susceptible to environment. Dwarf type, long fruit type, and red fruit type of okra were more susceptible to the environment than other types.
Keywords sowing sites; cultivation environment; Abelmoschus esculentus; quantitative traits
DOI 10.3969/j.issn.1000-2561.2019.12.016
近年來,黄秋葵(Abelmoschus esculentus)作为一种特色蔬菜被大众所熟知,其热度居高不下,目前已广泛栽培于世界各地[1]。其别名有补肾菜、洋辣椒、羊角豆、咖啡黄葵等,属一年生(热带地区为多年生)日中性自花授粉植物[2-3]。有研究表明,黄秋葵具有一定的保健作用,其果中富含丰富的氨基酸[4];黄秋葵嫩果中丰富的黏性物质主要是由多聚半乳糖、半乳聚糖和阿拉伯聚糖与果胶组成[5],具有降低血糖、血脂和减少胆固醇、提高免疫力等方面的效果[6];除了果实可以食用之外,茎叶和果皮还可以作为动物饲料[7-8]。20世纪90年代,我国开始从我国台湾地区和日本进行黄秋葵的引种栽培,主要用于出口,并未形成产业化生产经营[9]。经过20多年的发展,目前我国北京、山东、海南、台湾以及东南部地区等均广泛种植[10]。
2016年5月16日,黄秋葵(咖啡黄葵)列入中华人民共和国农业植物品种保护名录(第十批)。受保护的植物可以申请品种权,目前,黄秋葵新品种权的累积申请量仅为2份。新品种权的审查必须进行植物新品种特异性(distinctness)、一致性(uniformity)和稳定性(stability)测试(简称DUS测试),DUS测试需要依据该物种测试指南中所列出的性状进行品种描述,测试性状包括质量性状(QL)、假质量性状(PQ)和数量性状(QN),其中数量性状极易受到环境条件的影响,同一品种在不同年份、不同生态区的性状表达可能会有较大的差异。黄秋葵的栽培范围较广,我国南北方均有种植,生长环境差异较大,而由于南北的种植方式、气候条件等因素的影响,黄秋葵的数量性状存在一定的差异。因此,本研究通过在上海和儋州2个地点种植黄秋葵品种,研究不同生态区环境对黄秋葵数量性状表达的影响,明确各数量性状表达的差异,从而进行稳定性评价,为在不同测试区科学地筛选更为合适的标准品种提供理论依据。
1 材料与方法
1.1 试验地点
试验材料共16个黄秋葵品种,种植在上海和儋州2个DUS测试试验区(表1)。
1.2 材料
试验材料为16份黄秋葵资源品种(表2),由中国热带农业科学院热带作物品种资源研究所提供。分别种植于农业部植物新品种测试(上海)分中心测试基地和农业部植物新品种测试(儋州)分中心测试基地。
1.3 试验设计
上海:2018年4月17日播种,营养钵育苗培大,5月3日定植,露地栽培,小区高畦单行种植,畦面带沟宽1.6 m,株距75 cm,每品种不少于60株,2次重复。
儋州:2018年9月10日育苗,育苗移栽,9月26日定植,大棚栽培,株距80 cm,行距80 cm,每小区11株,2次重复,田间管理按大田生产管理方式进行。
1.4 性状观测
按照《植物品种特异性、一致性和稳定性测试指南 黄秋葵》要求,在盛花期(50 %以上植株开花),每个品种随机选取20个植株,每个植株取1个典型的完整叶,用测绘尺对叶片长度、叶片宽度、叶柄长度共 3个数量性状进行测量。在商品果采收期,每个品种随机选取20个植株,每个植株取1个果实,用游标卡尺对果实厚度进行测量。在果实的生理成熟期(即果实开始转色),每个品种随机选取20个植株,每个植株取1个成熟的果实,用卷尺和游标卡尺对果实长度和果实宽度进行测量。在果实完熟期(即种子成熟),每个品种随机选取20个植株,每个植株取1个成熟的果实,收集果实中的成熟种子,对每个果实中的种子数量进行计数。以20个样品的均值为最终值。
1.5 数据处理
采用WPS Excel 2016软件进行数据整理,计算各数量性状的最小值、最大值、平均值、中值、标准差、变异系数和t检验分析。
2 结果与分析
2.1 不同种植地点黄秋葵品种数量性状的稳定性和显著性分析
采集2个生态区16份黄秋葵的数量性状数据并进行汇总分析。从表3可以看出,上海生态区中变异系数大小排序为:果实厚度>果实长度>叶片宽度>单荚种子数>果实直径>叶柄长度>叶片长度,其中最高为果实厚度(31.2%),最低为叶片长度(15.2%)。儋州生态区中变异系数大小排序为:单荚种子数>果实长度>叶片宽度>果实厚度>叶片长度>果实直径>叶柄长度,其中单荚种子数(23.1%)最高,最低为叶柄长度(12.3%)。可以看出,2个生态区叶片宽度、果实长度、果实厚度的变异系数均大于叶片长度、叶柄长度和果实直径。7个数量性状中叶片宽度、叶柄长度、果实直径、果实厚度和单荚种子数在2个生态区之间存在显著差异;而叶片长度和果实长度差异不显著。
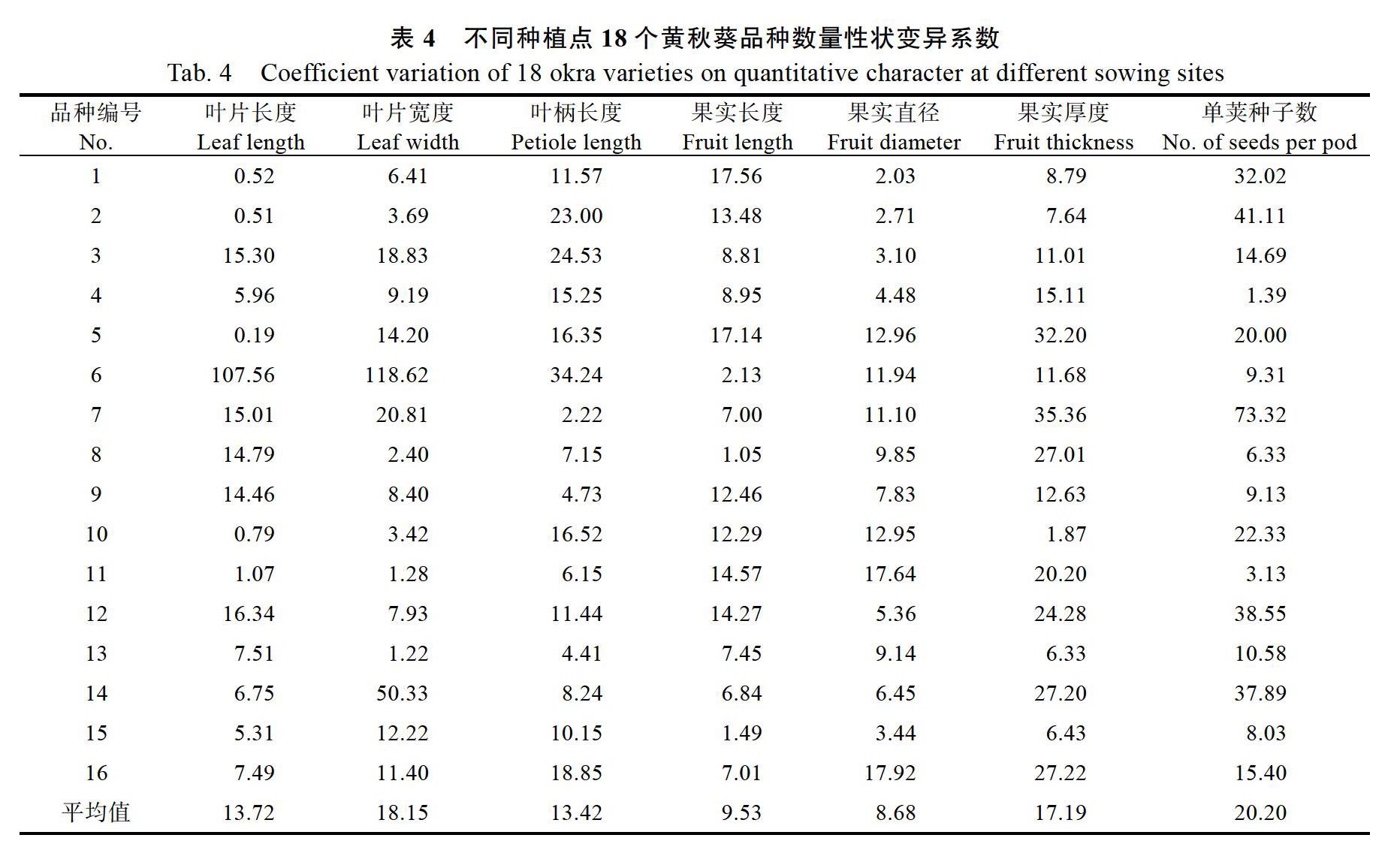
2.2 不同种植地点间黄秋葵各数量性状的变异系数
从表4可知,上海和儋州两地间黄秋葵各性状的变异系数呈现很大差异。总体看来,平均变异系数由大到小分别为单荚种子数>叶片宽度>果实厚度>叶片长度>叶柄长度>果实长度>果实直径。单荚种子数的平均变异系数最大,为20.20%,表明其受环境影响较大,但其中4号、11号品种的变异系数在0~3.13%,说明这2个品种的单
荚种子数受环境作用较小。性状叶片长度与叶片宽度的变异系数在品种间波动较大,表明这2个性状受品种及环境的互作效应较大,极为不稳定。而果实长度和果实直径这2個性状在品种间的平均变异系数相对较小,因此环境作用对其影响相对较小。而在16个品种中,6号品种在叶长、叶宽以及叶柄长度3个形状上的变异系数很大,说明该品种在叶上的性状极易受环境影响。
2.3 不同生态区各黄秋葵数量性状的变化
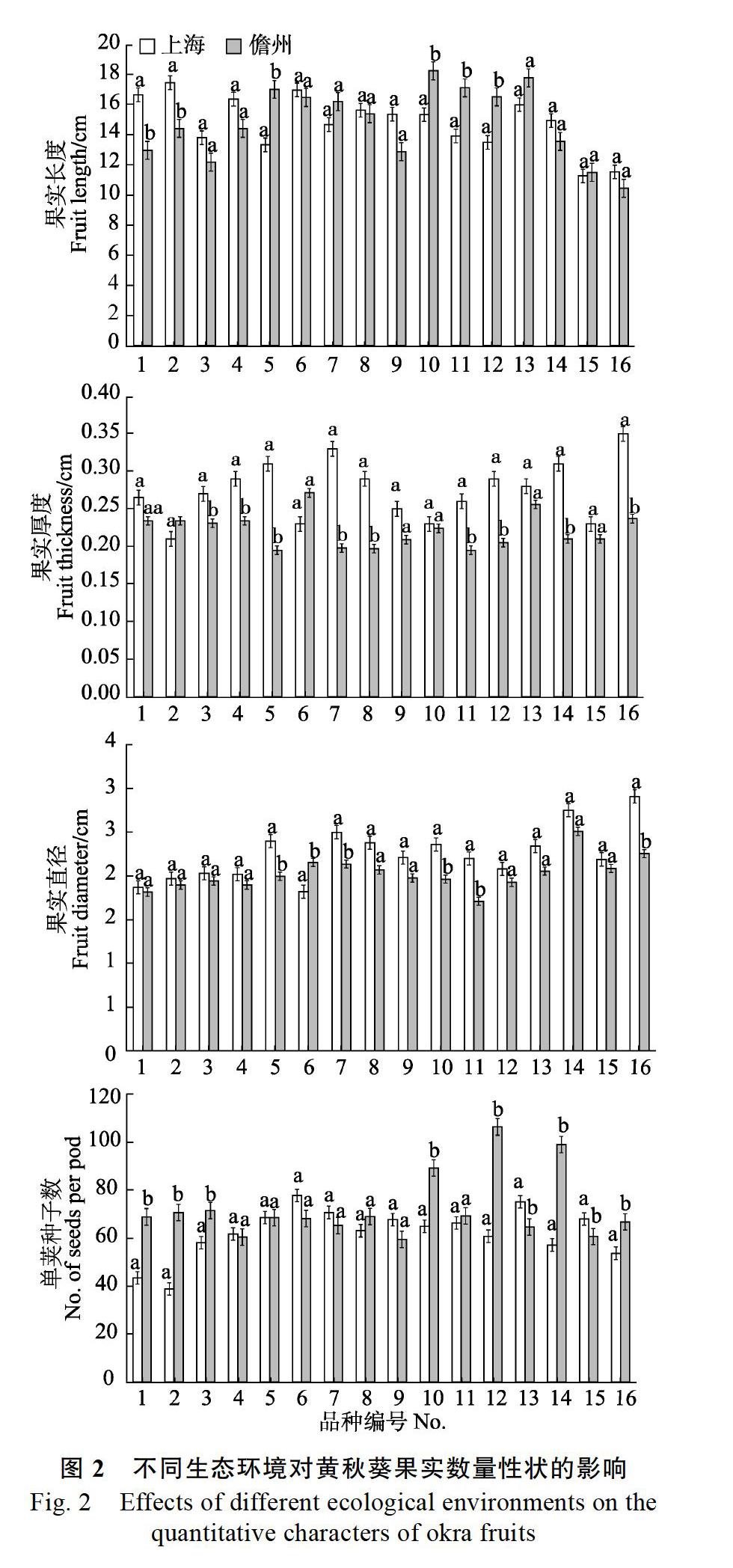
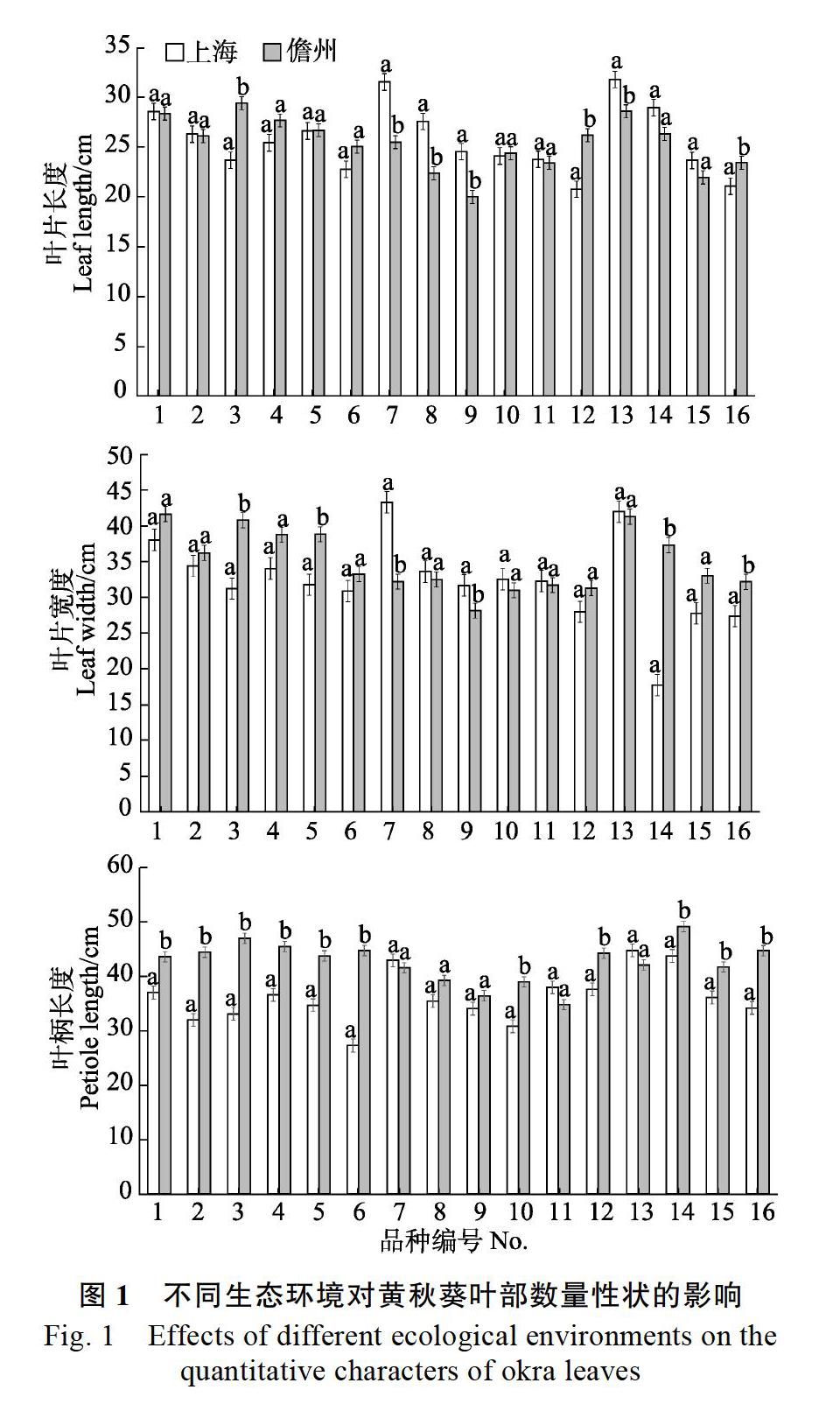
2.3.1 不同生态区对黄秋葵叶部数量性状的影响 由图1可知,16个品种中有7个品种在叶片长度上有显著差异,有6个品种在叶片宽度上有显著差异,有11个品种在叶柄长度上有显著差异,因此,叶柄长度这一性状相对来说更容易受到环境的影响。在性状叶片长度中,1号、2号、5号、10号、11号品种在两地的差异极不明显,而3号、7号、8号和12号品种在两地的差异极其显著;在性状叶片宽度中,8号和11号品种在两地差异极不明显,而3号、7号和14号品种的差异极其显著;在性状叶柄长度中,7号品种在两地差异极不明显,而2号、3号、12号、16号品种的差异极其显著。此外,3号和16号品种在叶部的3个性状上都有显著差异水平,说明这2个品种在叶部性状上受环境影响较大;而11号品种在3个性状上均无显著差异水平,说明该品种受环境影响相对较小。
2.3.2 不同生态区对黄秋葵果实数量性状的影响 从图2可以看出,16个品种中,在果实长度和果实直径2个性状上,均有6个品种有显著差异;在果实厚度和单荚种子数2个性状上均有9个品种有显著差异。在果实长度性状中,6号和8号品种在两地的差异极不明显,而1号、2号、5号和10号品种在两地的差异极显著;在果实直径性状中,2号和3号品种在两地差异极不明显,而5号、10号、11号和16号品种的差异极显著;在果实厚度性状中,15号品种在两地差异极不明显,
而在9个差异显著的品种中有8个品种为差异极显著;在单荚种子数性状中,4号和5号品种在两地差异极不明显,而1号、2号、10号、14号和16号品种的差异极显著。除此之外,5号、10号、11号、12号和16号品种在果实的3个性状上都有显著差异水平,说明该5个品种在果实性状上受环境影响较大;有6个品种在果实3个形状上无显著差异;而有9号品种在4个性状上均无显著差异水平,说明该品种受环境影响相对较小。
3 讨论
黄秋葵为日中性植株,其易受光照条件的影响,但对土壤的适应性较强。黄秋葵叶部数量性状受环境影响较大,徐丽等[11]通过露地和大棚2种栽培环境种植黄秋葵,研究表明黄秋葵叶片长度和叶片宽度受栽培环境影响较明显,这与本研
究结果一致;而在果实的数量性状上,果实长度和果实横径2个性状受环境影响程度与黄秋葵品种类型有关。李春梅等[12]在不同播期下种植黄秋葵,由于不同播期下环境温度和降雨量的不同,较低气温环境对黄秋葵株高、茎粗、叶面积、果形和品质等方面均有促进作用。本研究是通过在地理环境差别较大的2个试验地种植黄秋葵,上海的平均气温要低于儋州,而在果实直径和果实厚度这2个性状上,上海种植点的绝大部分品种均大于儋州,结合前人研究,较低气温可能对果实某些性状上有促进作用。
除了环境因素对黄秋葵各数量性状的影响外,其本身品种的特征也影响相关性状的表达。徐丽等[11]研究发现,绿秋葵的果柄长度和果柄粗度容易受到环境的影响,长果黄秋葵的果实长度与果实横径不易受环境影响。本研究中,绿果黄秋葵相较于红果更易受到环境的影响,红果的叶片宽度在2个地点间无明显差异;长果黄秋葵的叶片宽度和果实直径受环境影响不大;矮杆黄秋葵的果实直径和单荚种子数几乎不受环境影响。而同一性状在不同品种间的变异系数有很大差异,其根本原因可能与遗传纯度有关。
数量性状具有连续性、不确定性的特点,而其表达的结果会直接影响到对应性状的DUS测试结果,不同数量性状对环境反应程度不一样,对测试结果的重要程度也有差别[13]。在制定黄秋葵数量性状分级标准时,对观测时期进行相应的规定,根据不同栽培环境对数量性状分级区间进行调整[14]。因此,在选择DUS测试的标准品种时,尽量选择在数量性状上表现相对更稳定的品种。同样,在进行近似品种筛选时,对于不同生态区地点的测试结果要进行考量,在同等条件下,尽量选择同一生态区的测试结果作为参考,从而避免环境因素导致的数量性状表达结果差异。
参考文献
Chandra S, Saha R, Pal P. Arsenic uptake and accumulation
in okra (Abelmoschus esculentus) as affected by different arsenical speciation[J]. Bulletin of Environmental Contamination and Toxicology, 2016, 96(3): 395-400.
Osekita O S, Ariyo O J, Kehinde O B. Variation and inter-character association in the segregating F3 populations arising from two crosses of okra (Abelmoschus esculentus (L.) Moench)[J]. Moor Journal of Agricultural Research, 2000, 1(1): 32-36.
高 玲, 刘迪发, 徐 丽. 黄秋葵研究进展与前景[J]. 热带农业科学, 2014, 34(11): 22-29.
卢令格, 王光亚. 秋葵籽蛋白质的营养学研究[J]. 卫生研究, 1993, (4): 240-243.
王建蕊. 黄秋葵功能成分提取及應用研究[D]. 福州:福建农林大学, 2013.
刘晓霞. 黄秋葵花果胶类多糖的提取工艺及其性质的研究[D]. 杭州:浙江大学, 2014.
刘国道, 王东劲, 侯冠彧, 等. 黄秋葵茎叶粉对文昌鸡蛋黄着色的影响[J]. 中国农学通报, 2006, 22(7): 16-19.
石雪萍. 黄秋葵的研究进展及应用前景[J]. 中国野生植物资源, 2017, 36(4): 63-67.
龚 霄, 姜永超, 周 伟, 等. 黄秋葵研究进展及其应用[J]. 食品工业科技, 2018, 39(23): 329-333.
黄 吉. 黄秋葵种质资源多样性及应用研究[D]. 长沙:湖南大学, 2017.
徐 丽, 高 玲, 刘迪发, 等. 栽培环境对黄秋葵数量性状的影响[J]. 中国瓜菜, 2016, 29(11): 14-18.
李春梅, 曹 毅. 不同播期对黄秋葵生长及发育的影响[J]. 长江蔬菜, 2008(6): 31-32.
黄志城, 杨 坤, 周海涛, 等.不同生态环境对郑单958部分数量性状的影响研究[J]. 玉米科学, 2015, 23(5): 49-55.
徐 丽, 高 玲, 刘迪发, 等. 黄秋葵DUS测试主要数量性状的测量及分级[J]. 中国蔬菜, 2017, (12): 57-62.
猜你喜欢
福建农业科技(2019年3期)2019-09-10
农民致富之友(2019年19期)2019-08-02
山东农业科学(2017年2期)2017-03-15
农民致富之友(2016年22期)2017-02-07
吉林农业·下半月(2016年12期)2016-12-26
食品与健康(2016年12期)2016-12-17
家庭百事通·健康一点通(2014年11期)2015-01-19
周末·校园文学(2014年20期)2014-10-21
世界热带农业信息(2014年8期)2014-09-23
农村百事通(2014年3期)2014-07-24
