
不同植物乳杆菌发酵液抑菌活性及其主要有机酸组成比较
2019-04-02 03:42李书鸿柳陈坚任贝贝李强坤罗义勇李晓然
食品科学 2019年5期
李书鸿,柳陈坚,任贝贝,李强坤,罗义勇,杨 恩,李晓然*
(昆明理工大学生命科学与技术学院,云南 昆明 650500)
With the increasing health awareness of people, food safety has become a global trend for healthy consumption that relates between people’s physical health and national prosperity. Currently, foodborne pathogenic bacteria such as Staphylococcus aureus, pathogenic Escherichia coli, and Salmonella have a severe impact on food security. Chemical preservatives are the main means to preserve foods, but most chemical preservatives may pose a severe hazard to human health, such as deformity and induced cancer[1]. Biological preservatives are much better for human health, and can also maintain the sensory properties and nutritional components of food[2-3]. Therefore, discovering efficient, broad spectrum, and safe biological preservatives from microbial resources has become an important trend in the study of food safety.
Lactic acid bacteria (LAB) are important industrial microorganisms that are widely used in foods, medicines,and feed industries[4]. LAB can not only improve the food flavor and enhance food nutrition[5-7], but also maintain the intestinal micro ecological balance, enhance the body’s immunity[8], prevent cardiovascular disease[9], and reduce serum cholesterol[10]. Among LAB, Lactobacillus plantarum has been considered as one of the most safe and versatile species. Therefore, it has been extensively used in the food industry both as microbial starters and as probiotic microorganisms[11]. Furthermore, it has been shown that many L. plantarum strains have strong antagonistic activity against spoilage, pathogenic bacteria, and fungi[12-13]. Recent studies have shown that L. plantarum can produce different antimicrobial agents such as organic acids, hydrogen peroxide, diacetyl, and antimicrobial peptides, including bacteriocins[14-18]. Its antimicrobial effect on pathogenic and spoilage microorganisms has often been ascribed to the production of organic acids, including lactic and phenyllactic acids[19]. Nowadays, the increasing demand of consumers for natural and healthy food puts a focus on the fi eld of biological protection. The selection of microbial molecules and/or bacterial strains that are able to produce substances that can be used as preservatives has become an indispensable part of that fi eld. Therefore, L. plantarum with its good antibacterial effects on foodborne pathogenic bacteria could be a suitable candidate and can offer valuable opportunities for food preservation.
The main objective of this study was to compare the antibacterial activity of L. plantarum strains isolated from Douchi (a traditional fermented food) and to investigate the major components that account for its antimicrobial activity.This provides information for the development of new biological preservatives.
1 Materials and Methods
1.1 Strains and materials
Eighteen strains of L. plantarum were previously isolated from Douchi, a traditional fermented food in Yunnan Province, and were maintained in glycerol stocks at -80 ℃. The utilized pathogen indicator bacteria were Escherichia coli O157:H7 and Staphylococcus aureus ATCC 6538, which were obtained from our frozen stock culture collection.
1.2 Methods
1.2.1 Effects of carbon sources on antibacterial activity and pH
The 18 strains of L. plantarum, stored at -80 ℃, were grown on de Man-Rogosa-Sharpe broth (MRS broth) at 4‰ (V/V) inoculation and incubated at 30 ℃ for 24 h. The cultures were centrifuged for 10 min at 2 863 × g (4 ℃), and the pH of supernatants were measured via micro pH meter(AS ONE, Japan). The antibacterial activity of L. plantarum strains against pathogenic bacteria was analyzed via spot-on-lawn assay as described by Schillinger et al[20]. The 5 µL overnight cultures of L. plantarum strains containing 5 × 106CFU cells were spot on the center of MRS agar medium, and incubated at 30 ℃ for 24 h. Then, the plates were covered with 7.5 mL of soft Brain heart infusion agar(BHI agar, containing 0.75% agar) containing 106CFU/mL of an overnight culture of S. aureus ATCC 6538 suspension or 7.5 mL of soft Luria-Bertani agar (LB agar, containing 0.75%agar) containing 106CFU/mL of an overnight culture of E. coli O157:H7 suspension. Then, the plates were incubated at 30 ℃ for an additional 24 h. The formation of clear zones of growth inhibition around L. plantarum colonies and their diameters were recorded. The glucose in the MRS medium was replaced by the same amount of starch, sucrose,lactose, or fructose and antibacterial activity research was conducted according to the above method to explore the effects of different carbon sources on the antibacterial activity of L. plantarum.
1.2.2 Antibacterial activity of cell free supernatant and ultrafiltrate
The activated strains were inoculated in MRS broth at 108CFU/mL for expanded cultivation and were incubated at 30 ℃ for 72 h; then, centrifuged for 10 min at 2 863 × g(4 ℃), and the cell free supernatant was collected. In addition, proteinase K (50 µg/mL) was added to the appropriate amount of the supernatant and then incubated at 30 ℃ for 4 h. The treated supernatant was ultrafiltrated via millipore (molecular weight cut-off of 3 000 Da), and the ultrafiltrate was collected. Finally, the cell free supernatant and ultrafiltrate were fi ltered through a 0.22 µm fi lter.
Using the agar well diffusion method as described by Yang et al[21], E. coli O157:H7 and S. aureus ATCC 6538 were added to 20 mL of soft LB agar (containing 0.75% agar)or soft BHI agar (containing 0.75% agar), respectively, to keep the indicator bacteria concentrations at 106CFU/mL.After cooling and solidification of the culture media, they were punched with diameters of 7 mm; then, 100 µL cell free supernatant or underlayer filtrate were added to the holes of the culture media, and the same amount of MRS broth was added in the control group. The culture media were incubated at 30 ℃ for 24 h after the liquid was completely absorbed, and then, the diameters of the inhibition zones were determined to explore the antibacterial activity of cell free supernatant and underlayer fi ltrate, respectively.
1.2.3 Analysis of the major organic acids of cell free supernatant via HPLC
L. plantarum strains were inoculated in 10 mL MRS broth with the inoculation of 108CFU/mL and incubated at 30 ℃ for 72 h. The culture solution was centrifuged at 2 863 × g (4 ℃) for 10 min, and the cell free supernatant was filtered through a 0.22 µm microfiltration membrane of the water system. The concentrations of organic acids, including lactic acid, acetic acid, propionic acid, citric acid, and malic acid (LC, Sigma-Aldrich, USA), were determined via high performance liquid chromatography (HPLC) with a Atlantis T3 C18column (250 mm × 4.6 mm, 5 µm, Waters, USA) at 30 ℃ using 0.03 mol/L ammonium dihydrogen phosphatemethanol (95 : 5, V/V) as eluent, a fl ow rate of 0.8 mL/min,and a sample volume of 10 µL. The detection wave length was 210 nm.
1.2.4 Dynamic monitoring of growth curves, pH and antibacterial ratios
The three L. plantarum strains QB3-3, 5-1, and S8 were chosen for further research on the changes of growth curves, pH, and antibacterial ratios due to their different antibacterial effects with glucose as carbon source.L. plantarum strains activated with a concentration of 108CFU/mL were inoculated in MRS broth which had glucose added as carbon source and incubated at 30 ℃for 72 h. Bacterial suspension of 1 mL was extracted from samples every 3 h, respectively, and centrifuged at 2 863 × g(4 ℃) for 10 min to separate the supernatant from cells.Next, the bacteria were diluted with 0.9% physiological salt, the absorbance at 600 nm was measured via ultraviolet spectrophotometer, the colony number was calculated with Wheat’s method, and the pH of culture supernatant was determined. To determine the antibacterial ratios of L. plantarum strains, 300 µL supernatant was added to 4.5 mL BHI liquid medium containing 106CFU S. aureus ATCC 6538, and control group was 4.8 mL BHI liquid medium containing 106CFU S. aureus ATCC 6538. After culture for 6 h,the samples were diluted with 0.9% physiological salt and the absorbance was determined at 600 nm. The antibacterial ratio was calculated as follow.

Where N represented the colony number in the experimental group and N0represented the colony number in the control group at the same time.
The strains had inhibitory effects on S. aureus ATCC 6538 when antibacterial ratios were below one, while the strains had no inhibitory effects when antibacterial ratios were above one.
1.3 Statistical analysis
All experiments were performed in triplicates (n = 3)and the reported values are the average of the triplicate. Data were analyzed with Pearson’s correlation and significances of differences between data were performed via One-way ANOVA test, at a significance level of P < 0.05. All statistical analyses were performed using SPSS Statistics 22.0 software.
2 Results and Analysis
2.1 Effects of carbon sources on the antibacterial activity and pH
A spot-on-lawn assay was performed to study the effects of different carbon sources on the antibacterial activity and pH of different L. plantarum cultures (Tables 1-3). The test revealed a different range of antimicrobial activity, depending on L. plantarum strain tested, carbon source used, and pathogen considered.
All L. plantarum strains exhibited higher inhibitory effects on E. coli O157:H7 than S. aureus ATCC 6538 when glucose and lactose were used as carbon sources. The majority of L. plantarum strains exhibited higher inhibitory effects on E. coli O157:H7 than S. aureus ATCC 6538 when sucrose and fructose were used as carbon sources. However,none of the L. plantarum strains had antibacterial activity to the two pathogenic bacteria when starch was used as carbon source.
One strain had different antibacterial effects with different carbon sources, and the effects were also influenced by indicator bacteria. When E. coli O157:H7 was used as an indicator bacterium, glucose as carbon source can produce the highest inhibitory effect, and the pH of the culture supernatant was between 3.6-3.8 which had a negative correlation with bacteriostasis (r = -0.565, P < 0.05); the antibacterial effect of fructose as carbon source was lower than that of glucose; while the strains had no antibacterial activity with starch as carbon source, and the pH of the supernatant was between 6.1-6.8. When S. aureus ATCC 6538 was used as an indicator bacterium, fructose was the best fermented substrate, and the pH of the supernatant ranged between 3.7-4.5; the antibacterial effect of sucrose as carbon source was higher than that of glucose; while the strains also had no antibacterial activity with starch as carbon source.

Table 1 Effect of carbon source on antibacterial activity of L. plantarum cultures against E. coli O157:H7
There were different antibacterial activities with the same carbon sources in different strains. When glucose was used as the carbon source, the antibacterial effect of L. plantarum KY3-7 on S. aureus ATCC 6538 was twice that of YmL-4-5 and S8; however, the antibacterial effect of L. plantarum YmL-4-5 on E. coli O157:H7 was twice of S5 and S10. The overall trend of antibacterial activity to pathogenic bacteria with different carbon sources was: glucose > fructose >sucrose > lactose > starch; therefore, the subsequent experiment used glucose as the carbon source of culture.
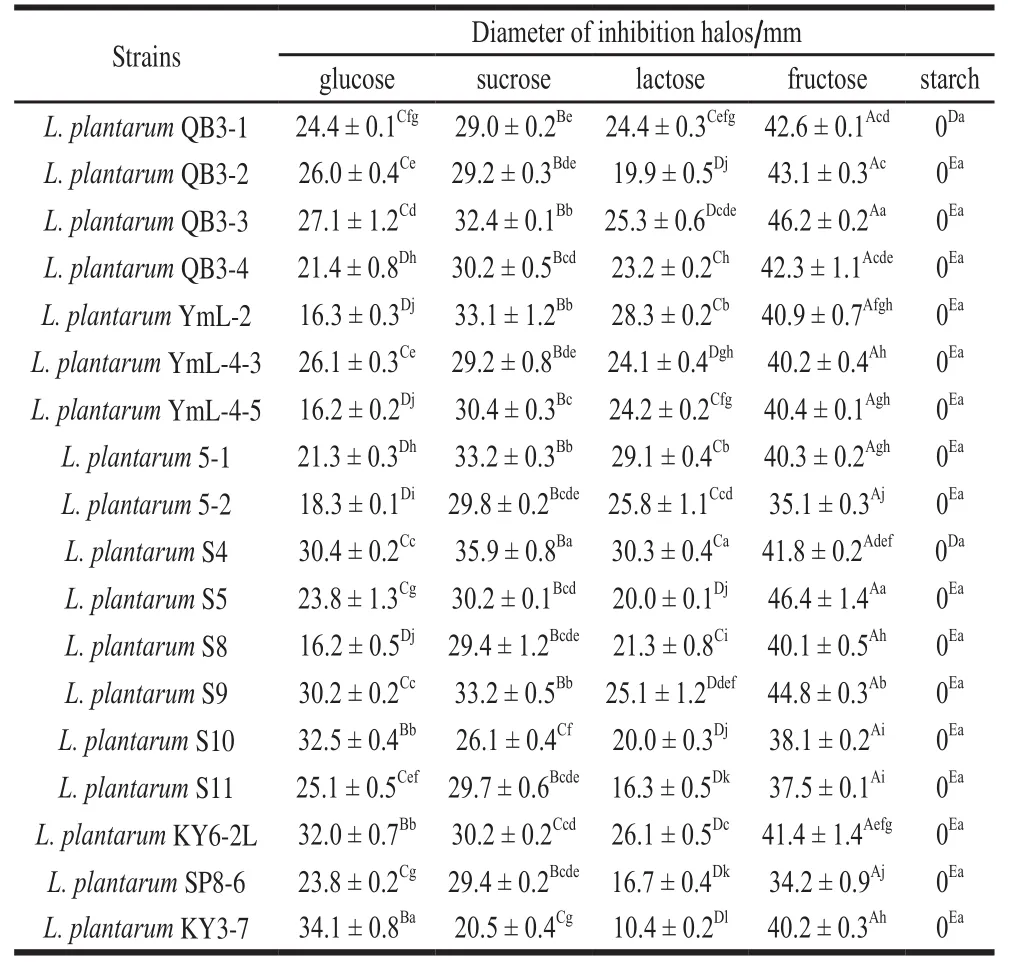
Table 2 Effect of carbon source on antibacterial activity of L. plantarum cultures against S. aureus ATCC 6538

Table 3 Effect of carbon source on pH of L. plantarum cultures
2.2 Antibacterial activity of cell free supernatant and ultrafiltrate
The antibacterial activity of cell free supernatant and ultrafiltrate of L. plantarum cultures were analyzed via the agar well diffusion method (Fig. 1). Almost all cell free supernatants and ultrafiltrates exhibited higher inhibitory effects on E. coli O157:H7 than S. aureus ATCC 6538 except for L. plantarum S4 and SP8-6. Most of the supernatants and ultrafiltrates of the same strain exhibited similar inhibitory effects on the indicator bacteria. Some of the supernatant exhibited slightly higher inhibitory effects than the ultrafiltrate, and both of them had no difference.
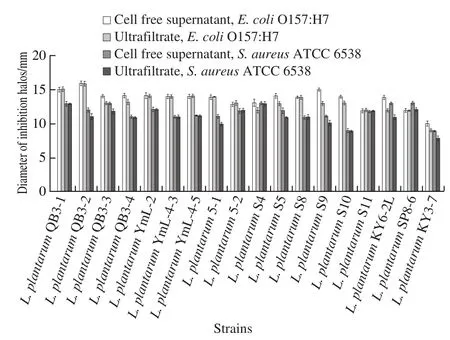
Fig. 1 Comparision of antibacterial activities of cell free supernatant and ultrafiltrate of L. plantarum
2.3 Analysis of the major organic acids in the cell free supernatant
According to the results shown in Fig. 2, the major organic acid in L. plantarum cultures was lactic acid, followed by malic acid, acetic acid, and citric acid, and the production of propionic acid was relatively low. The total acid production(4 885.4 µg/mL) was highest in the cultures of L. plantarum QB3-3, in which, lactic acid, citric acid, malic acid, and acetic acid accounted for 39.5%, 28.1%, 16.9%, and 15.5%,respectively. Lactic acid was detected in all cultures of 18 L. plantarum strains, while, propionic acid was not detected in cultures of L. plantarum QB3-1, QB3-3, YmL-4-5,and KY3-7, malic acid was not detected in the cultures of L. plantarum QB3-4 and S11, acetic acid was not detected in the cultures of L. plantarum S9, and citric acid was not detected in the cultures of L. plantarum 5-2 and S11. The correlation between organic acid production and antibacterial activity of L. plantarum cultures was analyzed, and the results showed that the contents of lactic acid and acetic acid had a positive correlation with the antibacterial activity of E. coli O157:H7 (r = 0.474 and 0.488, respectively, P < 0.05). The yield of lactic acid was between 930.9 and 3 008.7 µg/mL, and the concentration of acetic acid ranged from 0 to 898.3 µg/mL.
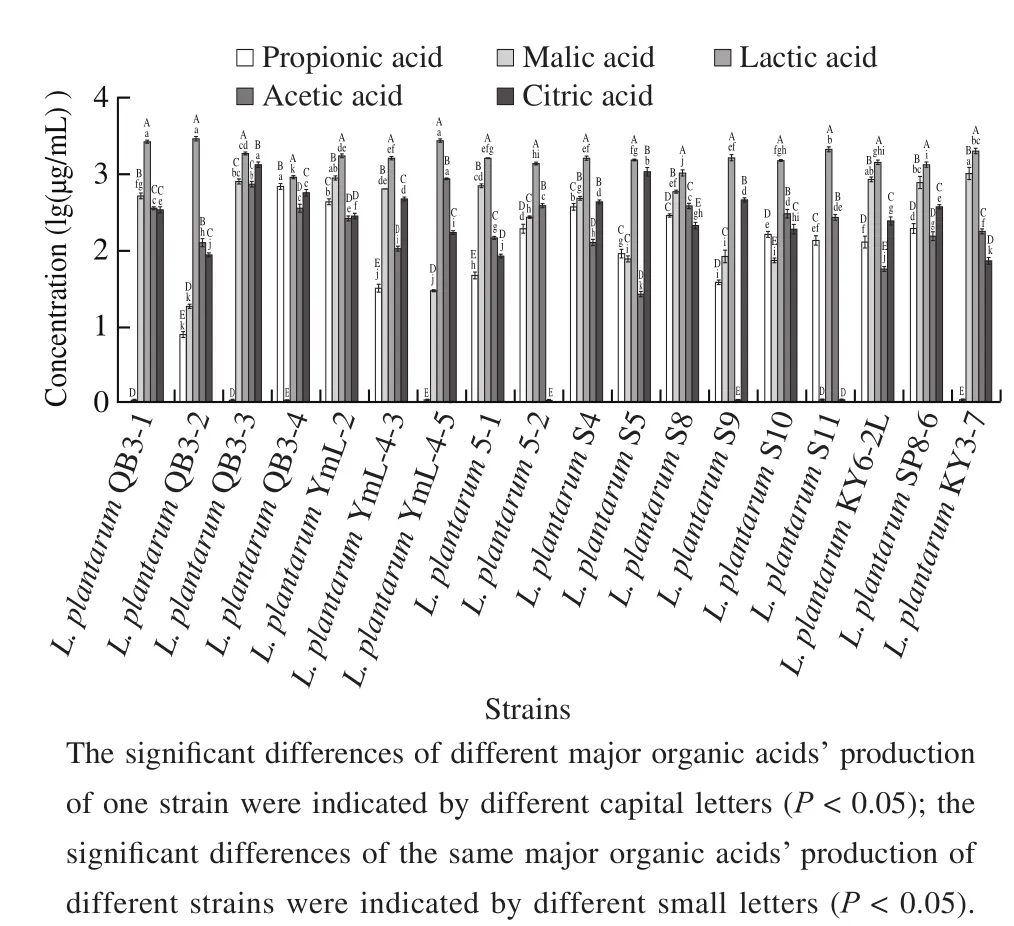
Fig. 2 Production of organic acids during L. plantarum fermentation with glucose as carbon source
2.4 Dynamic monitoring of growth curves, pH, and antibacterial ratios
L. plantarum QB3-3, 5-1 and S8 had different antibacterial effects with glucose as carbon source. The antibacterial effect of L. plantarum QB3-3 was relatively strong in 18 strains, L. plantarum 5-1’s antibacterial effect was relatively general, and L. plantarum S8’s antibacterial effect was relatively weak (Table 1 and 2). Therefore,dynamic monitoring of growth curves, pH, and antibacterial ratios was conducted (Fig. 3).
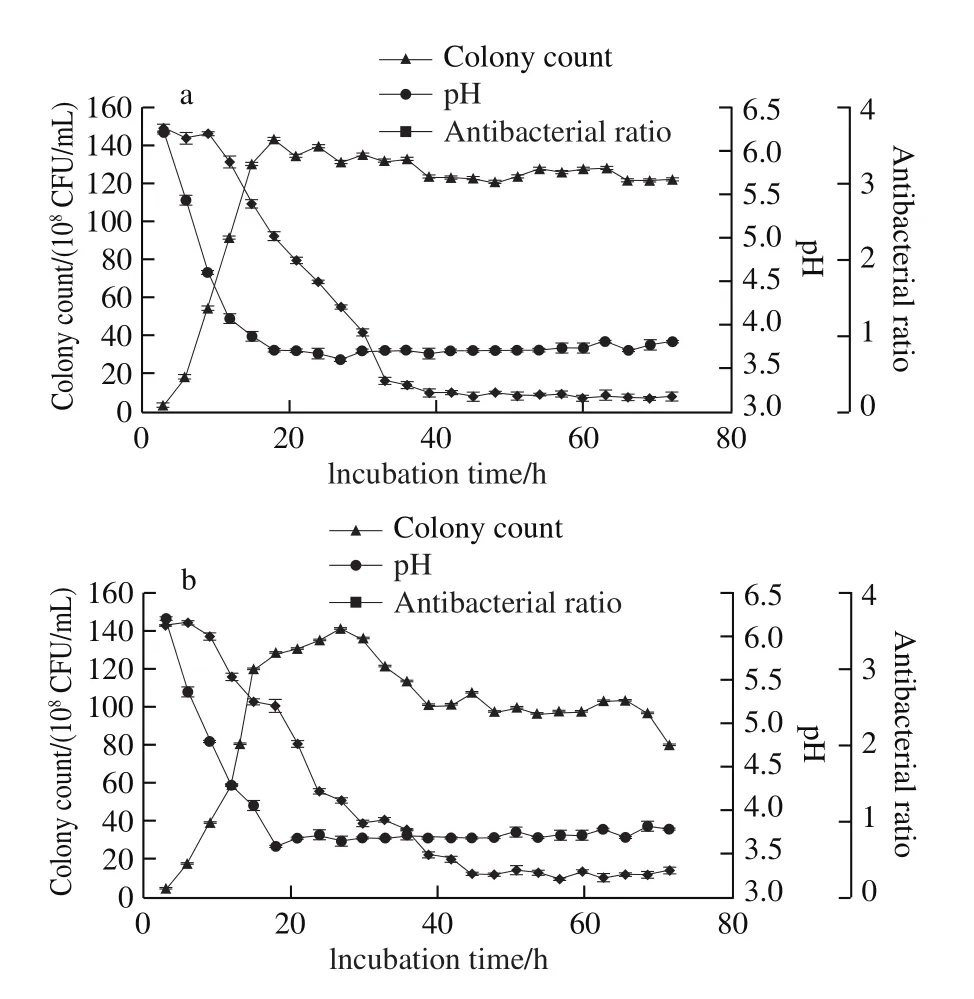
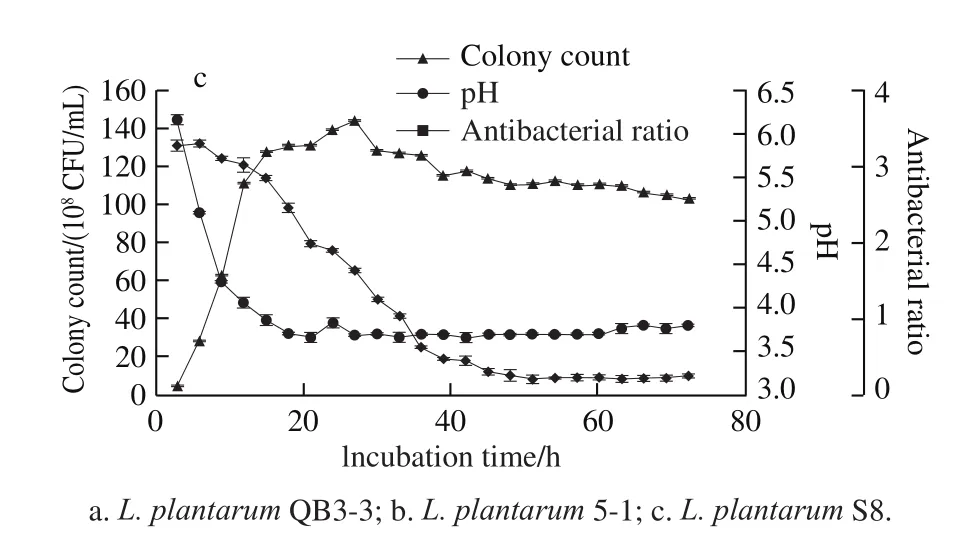
Fig. 3 Growth curves, pH, and antibacterial activity of three selected L. plantarum strains
From the results in Fig. 3a, L. plantarum QB3-3 entered the exponential phase after 3 h of incubation and entered the stationary phase after 15 h; however, the number of colonies decreased slightly after 18 h; then, the number of colonies changed little and were basically stable. The pH of the supernatant had a sharp decline after 3 h of incubation,following a minimum of 3.7 at 18 h; then, the pH remained basically unchanged and the overall trend was stable.Furthermore, the supernatant had no obvious antibacterial activity to S. aureus ATCC 6538 during the fi rst few hours,until the antibacterial ratio decreased to 0.404 at 33 h;after that, the antibacterial ratio decreased slowly, and the strongest antibacterial activity was obtained at 60 h with an antibacterial ratio of 0.174.
The results in Fig. 3b indicated that L. plantarum 5-1 entered the exponential phase after 6 h of incubation,and entered the stationary phase after 15 h; however, the number of colonies decreased slightly after 27 h, dropping to 1.01 × 1010CFU/mL at 39 h; then, changed little, basically stable. The pH of the supernatant declined sharply after 3 h of incubation, achieving a minimum of 3.6 at 18 h, and then, the overall trend of pH remained stable. In addition, there was an obvious antibacterial activity to S. aureus ATCC 6538 at 33 h,and after that, the antibacterial ratio decreased to 0.335 at 45 h,the antibacterial ratios remained relatively stable at about 0.3.
L. plantarum S8 had the same trend with L. plantarum QB3-3 and L. plantarum 5-1 (Fig. 3c). It also entered the exponential phase after 3 h of incubation, the number of colonies increased linearly until 15 h, and decreased slightly at 27 h, then remained basically stable. The pH of the supernatant declined sharply after 3 h of incubation, and remained at 3.7-3.9 after 18 h of incubation. Furthermore,the supernatant had obvious antibacterial activity to S. aureus ATCC 6538 at 36 h with an antibacterial ratio of 0.636. After 45 h, the antibacterial ratios were below 0.3, which showed good antibacterial activity.
3 Discussion
Foods produced via fermentation contain an abundant microflora[22]. The microbes that play a role in the fermentation are responsible for the release of anti-microbial factors[23]. This is the reason why most of the fermented foods have good antibacterial and antifungal properties and can keep a longer storage time. In light of this, this study focused on the antibacterial activity in cultures of L. plantarum strains isolated from Douchi, a traditional salt-fermented soybean food of Yunnan Province, and their metabolites, which have antagonistic activity.
Based on the results shown in Table 1 and 2, the best carbon source for L. plantarum on the antibacterial activity was glucose. A similar result was reported in a different experiment[24], where the authors studied the major organic acids and antimicrobial activity of Pediococcus acidilactici cultures, and they also found glucose was the best carbon source for P. acidilactici. As we all know, glucose and fructose are monosaccharides, which are easily absorbed and utilized, while, lactose and sucrose belong to disaccharides,and starch is a polysaccharide, which need to be broken down into monosaccharides to be used. The process of decomposition requires enzymes and energy; therefore,the cells will use monosaccharide as carbon sources first.However, there were different antibacterial activities with the same carbon sources in different strains, and this conclusion was in agreement with the results reported by Arena et al[14].Therefore, we hypothesized that different strains may have different glycosylated pathways, resulting in differences in the type and yield of organic acids.
Our findings demonstrated that the major antibacterial substances in the cell free supernatant were organic acids.Since the cell free supernatant was treated by proteinase K to remove the inhibitory effect of proteins, the molecular weight of lower filtrate after ultrafiltration remained below 3 000 Da, which meant that the antibacterial substances were mainly small molecule non-protein substances. Furthermore,there was no significant difference between the antibacterial activity of the cell free supernatant and the ultrafiltrate;therefore, the major antibacterial substances in the cell free supernatant were organic acids. Xiong Jun et al[25]studied the LAB strains with high antibacterial activity from fermented soybean. The results indicated that the inhibition mechanism of L. plantarum YM-4-3 was that of organic acid, and it was preliminarily conjectured that the mechanism of bacteriostatic used organic acid as leader, then bacteriocin and other chemistry substances. In general, organic acids can reduce the intracellular pH of the pathogenic bacteria. In this case,the coupling effect of electron transfer and ATP formation is relieved by organic acids as uncoupling agents, after which,the productivity of bacteria is reduced and its growth is inhibited. Moreover, on one hand, undissociated organic acids are liposoluble to different degrees, which have interacted with phospholipid molecules and lipopolysaccharide (LPS)of cytomembrane and then, the membrane stability is destroyed leading to the content loss. On the other hand,organic acids have the ability to inhibit the synthesis of biological macromolecule of pathogenic bacteria such as DNA and enzymes[26]. Compared to bacteriocins, which are mainly active against Gram-positive bacteria, organic acids exhibit a broader spectrum of antimicrobial action.Organic acids are also insensitive to proteases and have good stability. Therefore, organic acids have a great potential as biological preservatives.
The results shown in Fig. 1 demonstrated that the cell free supernatant and ultrafiltrate exhibited higher inhibitory effects on E. coli O157:H7 than S. aureus ATCC 6538. It has been reported that the antibacterial effects of organic acids produced by L. plantarum strains on E. coli was better than S. aureus, and could last longer, which may be related to the difference of cell wall between Gram-negative bacteria and Gram-positive[14]. The study of Dieuleveux et al[27]found that phenyllactic acid could act on the cytoderm of Listeria monocytogenes. Similarly, Li Qiangkun et al[28]also suggested that organic acids destroy the cell wall of E. coli and S. aureus. LPS is one of the components of the cytoderm of Gram-negative bacteria. As mentioned above,organic acids could interact with LPS and then destroyed the cell wall. Moreover, Kim et al[29]found that the combined treatments of medium-chain fatty acids and organic acids to E. coli O157:H7 showed clear membrane disintegration and/or cell death. Undissociated organic acids could penetrate the cell membrane and dissociative H+could destory the pH homeostasis in the cytoplasm leading to inhibition of the essential cell metabolism. Thereore, comepared to Gram-negative bacteria, Gram-positive bacteria may have better acid resistance. In addition to lowering the pH, there is a study reported that lactic acid also functions as a permeabilizer of the Gram-negative bacterial outer membrane and may act as a potentiator of the effects of other antimicrobial substances[30].
There was a significant difference in the content of organic acids between the 18 L. plantarum strains, and the common point was that lactic acid yielded the highest contents. This indicated that the major organic acid in L. plantarum cultures was lactic acid. The contents of lactic acid and acetic acid had a positive correlation with antibacterial activity to E. coli O157:H7. The organic acids could decrease the pH environment to inhibit pathogens.These results indicated that lactic acid and acetic acid had an important role for the inhibition, which could be used in industrial production. Poppi et al[31]determined pH values and organic acid concentrations from Lactobacillus isolates after 24 h of cultivation, and the results demonstrated that the pH values, lactic acid, and acetic acid production of L. plantarum isolate 30b were 3.83, (18.5 ± 0.3), and(4.2 ± 0.5) g/L, respectively. In the study of Schillinger et al[20],the minimum inhibitory concentration of lactic acid and acetic acid to S. aureus ATCC 25923 were 1.28 and 0.32 mg/mL, respectively. Cheng et al[32]who studied the influence of organic acids, including acetic acid,propionic acid, and lactic acid on the growth and survival of E. coli O157:H7, also reported that lactic acid had greater antimicrobial effects on pathogenic bacteria than acetic acid and propionic acid; however, the combination of the organic acids exhibited an important synergistic effect in inhibiting the pathogens, which was similar to our results.Moreover, it has been reported that short-chain organic acids had high antimicrobial activity against microorganisms due to the easy diffusion of non-dissociated forms through the cell membranes of pathogens[33]. Lactic acid exerted its antimicrobial effect by interfering with the maintenance of cytoplasmic membrane potential, thus inhibiting active transport, and hindering a variety of metabolic functions[34].
According to the study of Poppi et al[31], L. plantarum strains exhibited a higher inhibitory effect on E. coli than S. aureus due to the difference of cell walls between Gram-positive germs and Gram-negative germs. It was more difficult to inhibit S. aureus than E. coli; therefore, we chose S. aureus ATCC 6538 as indicator strain for further study. According to the results shown in Fig. 3, the supernatant had obvious antibacterial activity to S. aureus ATCC 6538 when the pH of supernatant was stable at about 3.7. However,Li Dongxia et al[35]who studied on antimicrobial activity of LAB including L. plantarum subsp. plantarum, reported that the antimicrobial substance remained active under acidic condition (pH 2.0-6.0), but became inactive under neutral and alkaline conditions (pH 7.0-9.0). The mechanism of organic acid permeation of cell membrane was pH-dependent since the undissociated forms were prevalent when the pH was below the pKa of the organic acid. According to our results, after about 45 h of incubation, the antibacterial ratios reached the minimum, and no significant changes were detected thereafter, which could be used as a reference for the fermentation time of further antibacterial substances. Xiong Jun et al[25]screened a strain of L. plantarum YM-4-3 with high efficiency from fermented soybean and found that YM-4-3 entered the exponential phase after 9 h of incubation and the stationary phase after 15 h, the pH tended to be stable at 24 h at 3.4 and had good antibacterial effects at 36-48 h,especially at 42 h. Their results were similar to the trends of the three strains we investigated.
4 Conclusions
This study showed the differences between Lactobacillus plantarum strains of the antibacterial activity to E. coli O157:H7 and S. aureus ATCC 6538. Our data indicates that the highest antimicrobial activity was observed in the case of L. plantarum YmL-4-5 when E. coli O157:H7 was the pathogenic bacteria and glucose was the carbon source. L. plantarum S5 had the strongest bacteriostasis when S. aureus ATCC 6538 was used as pathogen and fructose as carbon source. The composition of organic acids, which were the major antibacterial substances in L. plantarum cultures,were roughly identical, while there were obvious differences in content between different strains. The major organic acid was lactic acid (930.9-3 008.7 µg/mL) followed by malic acid and acetic acid. There was a positive correlation between the antibacterial activity to E. coli O157:H7 and the productions of lactic acid and acetic acid (r = 0.474 and 0.488,respectively, P < 0.05). The study provides a screening basis for the development of new biological preservatives.
猜你喜欢
中国机械工程(2022年18期)2022-10-08
当代水产(2019年11期)2019-12-23
中国机械工程(2019年19期)2019-10-28
中国机械工程(2019年11期)2019-06-13
中国机械工程(2019年4期)2019-03-06
中成药(2018年5期)2018-06-06
天然产物研究与开发(2018年4期)2018-05-07
中成药(2018年1期)2018-02-02
现代园艺(2017年19期)2018-01-19
农业环境科学学报(2017年2期)2017-03-20
