
东北边境地区啮齿动物中新埃立克体感染调查及groEL基因序列分析
2019-05-21 08:20王建伟2王秀红许志伟圃2吴益民
中国人兽共患病学报 2019年4期
王 卓,王建伟2,于 淼,冯 立,王秀红,许志伟, 田 圃2,吴益民
新埃立克体(CandidatusNeoehrlichiamikurensis,Nm)是近年来新发现的一种蜱媒病原体,隶属立克次体目无形体科新埃立克体属(CandidatusNeoehrlichia)[1-2]。Nm最早于1999年在荷兰的篦子硬蜱(Ixodesricinus)中检测到, 被认为是一种类似埃立克体(Ehrlichia-like)的病原体[3]。2004年,日本学者Kawahara等通过基于16S rRNA基因和groEL基因的系统发育分析将该病原体归为无形体科的一个新种,命名为CandidatusNeoehrlichiamikurensis[2]。近年来,欧洲多个国家和地区以及俄罗斯(远东、西伯利亚地区)、日本等地的蜱或啮齿动物中均有关于Nm发现的报道, 表明该病原体呈世界性分布[1-6]。流行病学调查研究证实我国一些省份小啮齿动物存在Nm感染, 并从蜱咬发热患者中发现Nm感染病例[7-8]。
东北地区是我国重要的虫媒传染病自然疫源地,特别是蜱媒传染病,媒介蜱和动物宿主携带病原体种类丰富,如黑龙江立克次体、查菲埃立克体、森脑病毒和伯氏疏螺旋体等[9-11]。为了解Nm自然感染状况和特点, 本研究在东北地区的密山、集安和宽甸采集啮齿动物标本,应用PCR方法进行Nm感染的病原分子生物学检测。
1 材料与方法
1.1鼠标本来源 选择黑龙江省密山县(2016年)低山丘陵耕地,吉林省集安市、辽宁省宽甸县(2012年)针阔混交林区和林缘耕地,采用夹夜法和笼夜法捕获啮齿动物。所有啮齿动物现场分类鉴定, 经消毒处理后解剖取其脾脏,置于1.5ml细胞冷冻管内,液氮保存。运回实验室后置-80 ℃保存。鼠标本采集地点见图1。
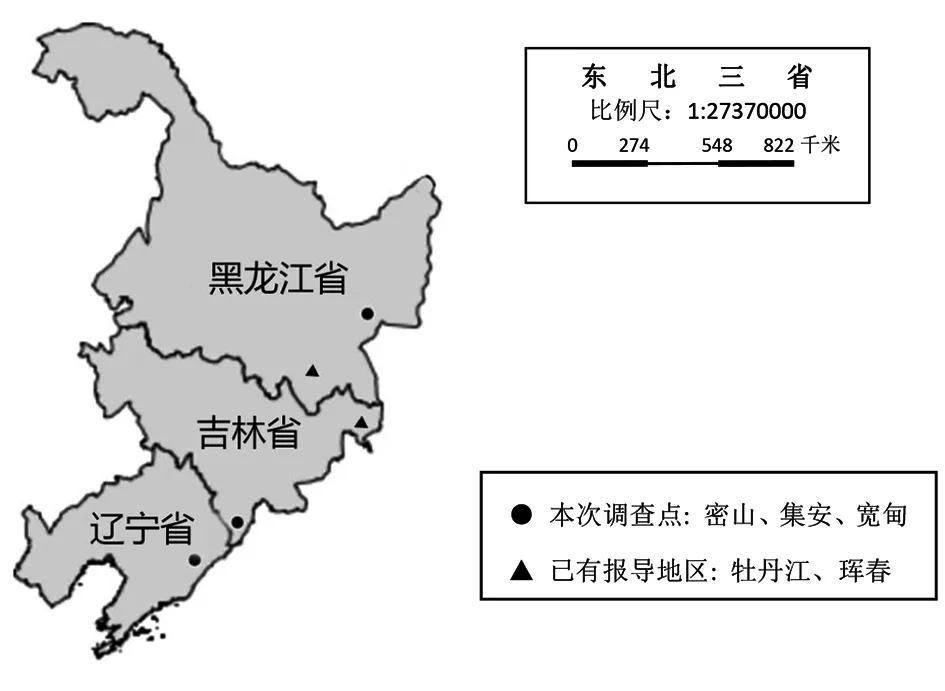
图1 本研究采样点示意图Fig.1 Geographic location of collected rodents
1.2鼠脾标本DNA提取 剪取约3 mm3脾组织,置于玻璃研磨器中充分研碎,应用DNeasy Blood &Tissue Kit(QIAGEN) 按说明书操作提取DNA,DNA提取液置-20 ℃保存备用。
1.3PCR扩增 用巢式PCR扩增NmgroEL基因(热休克蛋白基因),groEL基因引物按文献[8]设计,由上海申工生物工程公司合成。Nm的一轮PCR采用外引物Nm-out1 (5′-TGGCAAATGTAGTTGTAACAGG-3′)/Nm-out2(5′-TCTACTTCACTTGAACCGCCA-3′)。扩增条件: 94 ℃预变性5 min, 94 ℃变性30 s, 53 ℃退火30 s, 72 ℃延伸90 s, 40个循环后,72 ℃再延伸7 min。扩增产物目的片段大小为1 147 bp;二轮PCR采用内引物Nm-in1(5′-CCTATTAGTAAGCCTTATGGTAC-3′)/Nm-in2(5′-GAAGAATTACTATCTACGCTACC-3′)。二轮扩增条件除72 ℃延伸60 s, 35个循环不同, 其它与一扩相同。最终扩增产物目的片段大小为891 bp。
1.4序列测定及分子遗传进化分析 PCR 阳性产物由上海申工生物技术公司测序。应用Internet网中BLAST操作平台,将测序结果与GenBank中注册的NmgroEL基因序列进行比较,采用DNAstar及Mega5.0软件进行同源性分析和构建系统进化树。
2 结 果
2.1啮齿动物种类与分布 在东北地区3个调查点捕获啮齿动物共181只,包括6个鼠种,鼠种与分布见表1。黑线姬鼠(Apodemusagrarius)所占比例最高,占70.17%;其次为林姬鼠(Apodemuspeninsulae)、大仓鼠(Tscherskiatriton)和褐家鼠(Rattusnorvegicus), 分别占13.26%、8.29% 和7.18%。小林姬鼠(Apodemussylvaticua)和鼩鼱(Sorexaruneus)较少,分别占0.55%。
2.2啮齿动物中Nm感染情况 应用groEL基因的巢式PCR检测3地181只野鼠,5只感染Nm, 感染率为2.76%(表1)。其中, 集安(JA)和宽甸(KD)野鼠感染率分别为5.97%(4/67)和1.52%(1/66),密山野鼠未检出阳性, 3地野鼠Nm感染率差异无统计学意义(χ2=4.3124,P=0.1158)。感染鼠种主要涉及黑线姬鼠(2.36%)和林姬鼠(8.33%), 其它鼠种未检出阳性。
2.3基因序列比较 经序列比对分析,5份NmgroEL基因阳性测序标本的基因序列相似性为99.88%~100%。其中,3份黑线姬鼠标本(JA4、JA9,KD46)基因序列完全一致, 2份林姬鼠标本(JA14、JA23)基因序列基本一致, 与黑线姬鼠标本相似性为99.88%。
表1 不同地区不同鼠种的Nm感染情况
Tab.1 Prevalence ofCandidatusNeoehrlichiamikurensisin rodents from the 3 regions

啮齿动物种 类样本数(%)Nm检出阳性率(%)宽甸集安密山感染率黑线姬鼠127(70.17)1/49(2.04)2/39(5.13) 0/39(0)3/127(2.36)林姬鼠24(13.26)0/13(0)2/11(18.18)02/24 (8.33)小林姬鼠1(0.55)0/1(0)000/1(0)大仓鼠15(8.29)00/15(0)00/15 (0)褐家鼠13(7.18)0/3(0)0/1(0)0/9(0)0/13 (0)鼩鼱 1(0.55)00/1(0)00/1(0)合计181(100)1/66(1.52) 4/67(5.97)0/48(0)5/181(2.76)
2.4遗传进化分析 选择GenBank数据库中Nm注册的核苷酸序列构建进化树,并以浣熊新埃立克体(CandidatusNeoehrlichialotoris)、嗜吞噬无形体和查菲埃立克体作为参照。见图2。进化树分析显示集安和宽甸的5份鼠标本与俄罗斯远东地区全沟硬蜱株(FJ966363, FJ966359)及我国黑龙江省蜱株(JQ359077、KC106716)、黑龙江省Nm感染病人株(JQ359062)同处一个分枝,归类于ClusterⅠ。与欧洲Nm株(ClusterⅡ)、我国西南和东南地区的Nm株及日本的Nm株(Cluster Ⅲ、Cluster Ⅳ)有较大差异,说明Nm基因型分布与地理来源相关。
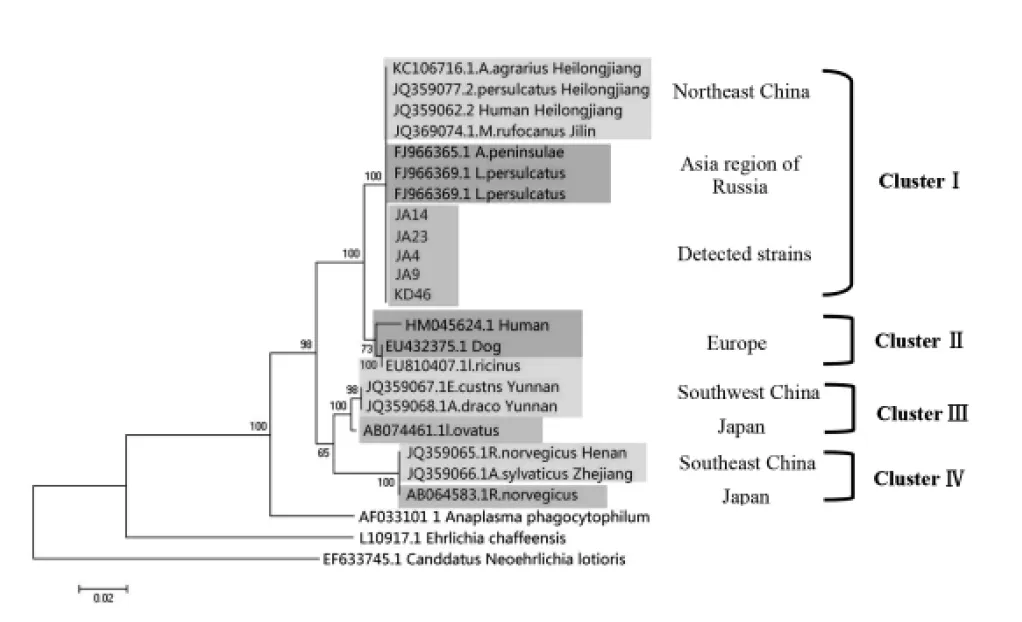
图2 基于groEL基因部分片段(827 bp)构建的系统进化树Fig.2 Phylogenetic tree ofCandidatus Neoehrlichia mikurensisinferred from 827 bp of thegroELgene sequences
3 讨 论
目前,全世界正面临着新发传染病的重大威胁,大部分属于人兽共患、媒介生物传播性疾病,而新发蜱媒病原体(如伯氏疏螺旋体、黑龙江立克次体、嗜吞噬细胞无形体等)引起的人类疾病即属于这一范畴。Nm是新发现的归属于无形体科的蜱媒病原体,巳知硬蜱属(Ixodesspp.)和啮齿动物分别为其主要的传播媒介和宿主动物[1-2,4-6]。近年来,在欧洲多个国家和地区,澳大利亚、尼日利亚及我国周边国家俄罗斯、日本、蒙古等地均有Nm相关报导[1-8]。北美地区浣熊中发现了与Nm进化关系较近的病原体,其命名为CandidatusNeoehrlichialotoris[12]。我国Nm的研究起步较晚,2003年Pan等[13]在广州褐家鼠(Rattusnorvegicus)脾脏样本中检测到的16S rRNA基因与无形体科埃立克体属(Ehrlichia)有明显区别,暂命名为Ehrlichiasp.rattus variant[13]。2012年Li等[8]应用PCR进行鼠中新埃立克体感染调查, 我国11个调查地区中有7个地区鼠标本Nm检出阳性,为我国多个地区可能存在Nm自然疫源地提供了分子流行病学证据。目前,世界范围内Nm感染病例报导较少,自2010年发现第一例感染患者以来仅有报导13例,欧洲散发病例6例[1,14],其余7例从我国黑龙江省牡丹江林区蜱咬发热病人发现[7]。
Burri等人通过动物接种实验证明啮齿动物为Nm的动物宿主[16], 在欧洲多个国家的啮齿动物中均发现Nm感染,涵盖6个鼠种, 不同地区啮齿动物Nm感染率差异较大, 如德国感染率高达52.7%, 而法国仅1.8%[1,5]。在亚洲,中国、日本、俄罗斯(西伯利亚和远东地区)等多个地区的啮齿动物中多个鼠种存在Nm感染[1,8]。本研究对我国东北边境地区三个调查点进行捕鼠采样,通过分子生物学方法调查鼠中Nm感染情况,从集安和宽甸黑线姬鼠和林姬鼠检出Nm5份, 感染率为2.76%(5/181), 说明野鼠可能是该病原体的重要储存宿主, 调查地区可能存在Nm自然疫源地。Li等报导[7-8],我国不同地区鼠类Nm感染率为4.0%(1.5%-13.8%), 黑龙江省牡丹江林区鼠类感染率为3.8%, 本研究结果(2.76%)与国内报导基本一致。以往的研究证明,集安和宽甸的不同鼠种携带多种蜱媒病原体,包括斑点热立克次体、查菲埃立克体、无形体、伯氏疏螺旋体等, 并存在复合感染[10,15], 本研究从该两地鼠类检出了Nm, 说明该地区存在多种蜱媒传染病并存的自然疫源地。
无形体科的遗传进化分析主要以groEL基因、rrs基因和gltA基因具有较好的分辨力。Kawahara等采用rrs基因、groEL基因系统发育分析将Nm归类为无形体科一个新种[2]。2012年Li等[8]依据rrs基因、groEL基因系统发育分析可将Nm分为4种基因型,我国东北地区以及俄罗斯亚洲地区的Nm基因型为Cluster I, 欧洲的Nm均为Cluster II, 我国西南和东南地区以及源于日本的Nm分别为Cluster III和Cluster IV(图2), 显示出基因型分布与地理来源密切相关[7]。本研究获得的NmgroEL基因序列与俄罗斯亚洲地区以及我国东北地区已报道的序列一致,属于ClusterⅠ型。
本次研究结果表明,调查地区啮齿动物存在Nm感染,感染率为2.76%,基因型别为ClusterⅠ型, 应重视Nm对人的潜在威胁。Nm作为一种新发的对人存在致病性的蜱媒病原体,其流行病学调查研究较少,进一步开展媒介蜱和啮齿动物Nm流行病学研究以及感染病例调查,为自然疫源地确定及该病的防治提供科学依据。
利益冲突:无
本文引用格式:王卓, 王建伟, 于淼, 等. 东北边境地区啮齿动物中新埃立克次体感染调查及groEL基因序列分析[J]. 中国人兽共患病学报, 2019,35(4): 330-333. DOI: 10.3969/j.issn.1002-2692.2019.00.032
猜你喜欢
环球时报(2022-04-07)2022-04-07
海外星云(2021年6期)2021-10-14
中国当代医药(2021年24期)2021-09-27
智慧健康(2021年5期)2021-03-30
科学(2020年3期)2020-11-26
科学(2020年3期)2020-11-26
动漫星空(兴趣百科)(2020年5期)2020-06-10
学生天地(2018年29期)2018-10-12
消费导刊(2016年7期)2017-08-12
大观(2016年6期)2016-07-05
