
聚块大小对双斑长跗萤叶甲空间分布的影响
2019-06-11 11:55毕守东张书平余燕王振兴李尚
植物保护 2019年1期
毕守东 张书平 余燕 王振兴 李尚


摘要为了明确聚块大小对茶园双斑长跗萤叶甲空间分布的影响,为确定最佳样方面积提供科学依据。用聚块样方方差分析方法和聚集强度指数法研究两种茶园两年双斑长跗萤叶甲空间分布的规律性,聚集格局的差异和种群聚集均数变化。不同大小聚块的均方差均出现1~2个峰值,聚块中有1~8个基本样方(K)时,随着K的增大,聚集格局的扩散系数C和种群聚集均数λ都不断增大,其他格局时C和λ不断变小,聚块内基本样方数K为2、4、8时与K为1时的聚集程度|w|值均小于|w|0.05。茶园双斑长跗萤叶甲存在1~2种不同尺度的聚集空间,空间分布具有规律性,个体群占据最小范围为2个基本样方,即4 m2,为最佳样方的选择提供了科学依据。
关键词双斑长跗萤叶甲;空间分布格局;聚块样方方差分析;种群聚集均数
中图分类号:Q 968.1
文献标识码:A
DOI:10.16688/j.zwbh.2018147
双斑长跗萤叶甲Monolepta hieroglyphica (Motschulsky) 国外主要分布于东亚和东南亚的俄罗斯(西伯利亚地区)、朝鲜、日本、越南、印度、新加坡、菲律宾、马来西亚、印度尼西亚等国家,在中国分布较广,为多食性害虫,主要为害玉米、棉花、向日葵、谷子、豆类和十字花科蔬菜等多种经济作物,还取食苍耳、葎草、马齿苋、苘麻、藜、龙葵等数十种杂草[1],也是茶树上的重要害虫之一,成虫、幼虫均可为害茶树,是近年南方茶区为害成灾的新害虫[2]。张聪等[35]研究了玉米田中该虫的发生特点和抽样方法,王立仁等[6]通过研究为害玉米等作物的双斑长跗萤叶甲,提出了抢种、深翻和施药相结合的防治措施。李广伟等[7]和袁海滨等[8]研究了温度对双斑长跗萤叶甲成虫寿命及生殖力的影响。土地含水量、温度等理化特性对双斑长跗萤叶甲的产卵量影响作用很大[913],聂强等[14]研究了该虫的取食选择性。李尚等[15]和余燕等[16]研究了双斑长跗萤叶甲的优势种天敌动态。植保工作中野外抽样调查是重要的内容,样本容量和样本范围大小影响总体真值估计的准确性。双斑长跗萤叶甲种群的聚块大小对其聚块均方差及分布格局的影响以及种群聚集原因和个体群聚集的最小范围均未见报道。本文利用聚块样方方差分析等研究‘乌牛早和‘白毫早茶园2015年和2016年双斑长跗萤叶甲聚块大小对聚块均方差及对分布格局的影响,并研究种群聚集原因和个体群聚集的最小范围,为双斑长跗萤叶甲抽样调查时采用的样方大小提供科学依据。
1材料与方法
1.1调查地点和时间
调查地点为安徽农业大学科技示范园茶园,调查茶树品种为‘乌牛早和‘白毫早,树龄13年。每品种茶园面积为0.2 hm2。‘乌牛早茶园和‘白毫早茶园调查时间均为2015年3月28日—11月14日,2016年3月27日—11月17日。约15 d调查一次,2015年调查17次,2016年调查16次。两种茶园周边为其他品种茶园,茶园按常规措施管理,但不施用农药。
1.2调查方法
采用平行跳跃法随机在茶园选取3行,茶树行距为1 m,每行间隔1 m取2 m长的样方。每行10个样方,共取30个样方,先目测调查,每样方随机选取10片叶,调查一些不易振落害虫及天敌种类和个体数,然后用沾有洗衣粉水液的搪瓷盘对样方中的所有枝条进行盘拍(搪瓷盘口长为40 cm,宽30 cm,洗衣粉水溶液浓度为1 000倍),调查记载害虫及其天敌物种数和个体数,对于一部分不能准确鉴定的物种样本编号保存,装毒瓶带回室内鉴定或请专家鉴定。
1.3数学分析方法
根据聚块样方方差分析的要求,参与数学分析的资料是第一个样方连续至第24个样方的调查资料。
1.3.1双斑长跗萤叶甲空间动态的聚块样方方差分析
聚块样方方差分析是在不同大小样方上的方差分析,是一种简单、有效的生态学空间格局分析方法。该法要求供试田块上的样方在空间上相互连接,随着聚块所包含的基本样方数目K从1,2,4,8…(指数级数)不断增加,聚块方差值常随之改变,通过不同大小聚块方差值的变化,了解研究对象随尺度增大的变化动态[17]。
2.1‘乌牛早茶园双斑长跗萤叶甲的聚块样方方差分析
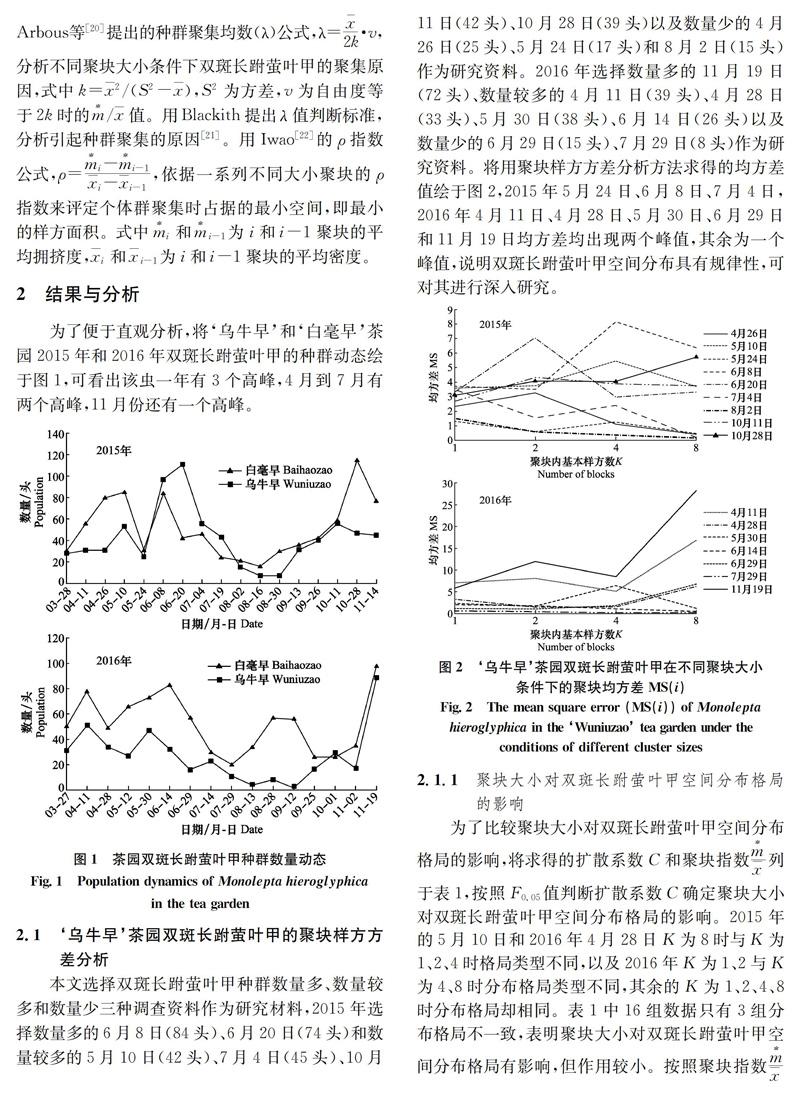
本文选择双斑长跗萤叶甲种群数量多、数量较多和数量少三种调查资料作为研究材料,2015年选择数量多的6月8日(84头)、6月20日(74头)和数量较多的5月10日(42头)、7月4日(45头)、10月11日(42头)、10月28日(39头)以及数量少的4月26日(25头)、5月24日(17头)和8月2日(15头)作为研究资料。2016年选择数量多的11月19日(72头)、数量较多的4月11日(39头)、4月28日(33头)、5月30日(38头)、6月14日(26头)以及数量少的6月29日(15头)、7月29日(8头)作为研究资料。将用聚块样方方差分析方法求得的均方差值绘于图2,2015年5月24日、6月8日、7月4日,2016年4月11日、4月28日、5月30日、6月29日和11月19日均方差均出现两个峰值,其余为一个峰值,说明双斑长跗萤叶甲空间分布具有规律性,可对其进行深入研究。
2.1.1聚块大小对双斑长跗萤叶甲空间分布格局的影响
为了比较聚块大小对双斑长跗萤叶甲空间分布格局的影响,将求得的扩散系数C和聚块指数列于表1,按照F0.05值判断扩散系数C确定聚块大小对双斑长跗萤叶甲空间分布格局的影响。2015年的5月10日和2016年4月28日K为8时与K为1、2、4时格局类型不同,以及2016年K为1、2与K为4、8时分布格局类型不同,其余的K为1、2、4、8时分布格局却相同。表1中16组数据只有3组分布格局不一致,表明聚块大小对双斑长跗萤叶甲空间分布格局有影响,但作用较小。按照聚块指数的判断标准,16组数据有2015年5月10日、5月24日、7月4日、10月28日和2016年4月11日、4月28日、6月14日、6月29日、7月29日、11月29日共10組分布格局一致,2015年6月8日、10月11日,K为1时与其他三个的格局不同,6月20日K为8时与其他三个的格局不同,其余的是K为1、2或K为1、8与其他不同。综合分析表明聚块大小对双斑长跗萤叶甲的空间分布有影响,但影响较小。
2.1.2聚块大小对双斑长跗萤叶甲种群聚集程度的影响
将聚块内基本样方数K为2、4、8时与K为1时的双斑长跗萤叶甲的聚集程度差异值|w|列于表2,可看出所有的|w|值均小于|w|0.05值,K为2、4、8时与K为1时的|w|值差异均不显著,表明聚块大小对双斑长跗萤叶甲的聚集程度影响不显著。
2.1.3聚块大小对双斑长跗萤叶甲的种群聚集均数λ值的影响
聚块大小对双斑长跗萤叶甲λ值的影响,即对双斑长跗萤叶甲空间分布聚集强度的影响,λ值也列于表2。可看出λ值为正值时随着聚块变大,λ值也随之变大,如2016年11月19日,λ值为负值时随着聚块变大,λ值随之变小,因为λ=2kv,K=2S2-,聚集分布时,K>0时,2>0,S2-必大于0,其他分布时K<0,2>0,S2-必小于0,所以出现以上情况。按照Blackith的标准,λ>2时是害虫自身原因引起的聚集,λ<2时是环境中某一因子引起的,表2中2015年5月10日、6月8日、10月11日、10月28日和2016年4月11日都是K为2、4、8时,λ都大于2,而K为1时,λ小于2,表明聚块大小对λ值有影响。
2.1.4不同聚块大小条件下双斑长跗萤叶甲的ρ指数
将‘乌牛早茶园2015年和2016年聚块内基本样方数由1至2,2至4,4至8个变化的ρ指数分别列于表3和表4,表3可看出,聚块内样方数由2至4变化时ρ的平均值为1.019 9±0.050 8,大于聚块内样方数K由1至2和由4至8变化的ρ指数的平均数,表4聚块中K由2至4变化时ρ指数平均值为1.128 7±0.114 4,也大于其他两种情况,表明双斑长跗萤叶甲个体群聚集的最小范围为2个基本样方即4 m2。
2.2‘白毫早茶园聚块大小对双斑长跗萤叶甲空间分布格局的影響
为了进一步分析聚块大小对双斑长跗萤叶甲空间分布的影响,用与‘乌牛早茶园同样的方法研究‘白毫早茶园双斑长跗萤叶甲不同聚块下的空间格局变化,2015年选择数量多的10月28日(85头)、数量较多的4月26日(63头)以及数量少的8月2日(18头);2016年选择数量多的11月19日(92头)、数量较多的5月30日(51头)以及数量少的7月29日(18头)作为研究材料。2015年3个时间点的MS(i)均有一个峰值。
2016年5月30日和11月19日均有两个峰值,7月29日有一个峰值,表明双斑长跗萤叶甲的空间分布具有规律性。按照C值判断标准,6组数据有5组的K为1、2、4、8同属一种分布格局,只有2015年10月28日K为1、2为聚集格局,K为4、8时为非聚集格局。由C值大小可看出,C值大小与样方中个体总数没有直接关系,如2015年的数量较多的4月26日,K为1、2、4、8时都是聚集格局,数量多的10月28日反而不全是聚集格局,2016年数量较多的5月30日,K为1、2、4、8时都是聚集格局,数量多的11月19日只有K为1时是聚集格局。用David等1954年提出的|w|公式分析的结果表明,不同聚块大小的双斑长跗萤叶甲的空间分布聚集程度差异不显著。‘白毫早茶园该虫的种群聚集均数λ值的变化与‘乌牛早茶园相同。
将‘白毫早茶园2015年和2016年的ρ值列于表5,可看出聚块样方数由2个变为4个时的ρ的平均值为1.005 3±0.088 4,大于其他情况,表明双斑长跗萤叶甲个体群占据的最小范围为2个基本样方,即4 m2,同‘乌牛早茶园的结果一致。
3小结与讨论
利用聚块样方方差分析法研究‘乌牛早茶园和‘白毫早茶园2015年和2016年聚块大小对双斑长跗萤叶甲空间分布的影响以及个体群占据最小范围的确定,结果是:
1)不同大小聚块的聚块均方差值均出现1~2个峰值,双斑长跗萤叶甲的空间分布是有规律性的。
2)双斑长跗萤叶甲在聚块内基本样方数为2、4、8时与为1时的|w|值差异不显著,即不同聚块大小条件下双斑长跗萤叶甲的聚集程度差异不显著。
3)在不同聚块大小条件下,聚集格局时双斑长跗萤叶甲的聚集是本身原因引起的,在λ值为正值时λ值随K的增大而增大;λ值为负值时随K的增大而变小。
4)不同聚块大小条件下双斑长跗萤叶甲在聚集格局时,随着K的增大,扩散系数C值也增大,在其他格局时随着K的增大,C值变小,聚块指数没有这种情况。
5)用不同大小聚块的ρ指数判断双斑长跗萤叶甲个体群聚集时占据的最小范围为2个基本样方,即4 m2。
采用聚块样方方差方法研究动物的空间分布格局动态是一种简便易行的方法。刘飞飞等[23]还用该法研究蜘蛛类天敌与茶尺蠖幼虫的空间关系,表明在聚块内有1~32个基本样方时随着聚块内基本样方数的增多,聚集格局的扩散系数C值不断增大,均匀格局的C值不断减少,基本样方数K为2、4、8、16和32时与K为1时之间的空间聚集程度差异均不显著;在种群聚集均数λ为正值时,随着聚块内基本样方数的增加,种群聚集均数λ不断增大;茶尺蠖个体群聚集格局的最小面积是聚块中具有1个基本样方。王建盼等[24]用聚块样方方差分析法研究柑橘粉虱Dialeurodes citri的空间格局变化,基本结论是一致的。
双斑长跗萤叶甲的卵在土壤表面0~15 cm深处越冬,翌年春夏季孵化,幼虫共3龄、主要取食寄主根系,成虫取食寄主叶片,具有假死性,能短距离飞翔,一般一次飞翔2~5 m,雌成虫在25℃时平均产卵量93.8粒[25],先把腹部末端插入土中缝隙,一般会分批产卵[26],由此可看出该虫受生物学习性影响,具有聚集生活习性,但同时善于短距离飞行,在一定空间范围内容易扩散,小范围(K=1时)的随机格局可扩散形成大范围(K=2、4、8时)的随机格局,或者小范围(K=1时)的聚集分布格局可扩散成大范围(K=2、4、8时)的聚集格局。从表1可看出,‘乌牛早茶园2015年4月26日、5月24日、6月8日、6月20日、7月4日、8月2日、10月14日和10月28日及2016年5月30日、6月4日和7月29日,聚块内基本样方数K为1时是随机分布,K为2、4、8时也是随机分布,2016年4月11日、4月28日、6月29日、11月19日聚块内基本样方数K为1时是聚集分布,K为2、4、8时仍然是聚集分布,表明聚块大小对双斑长跗萤叶甲的空间分布影响较小,该虫空间分布具有明显的规律性,这是该虫产卵特性和善飞习性等生物学特性所致。
本研究中种群为聚集格局时种群聚集均数λ随着聚块内基本样方数的增多,λ值不断增大,即双斑长跗萤叶甲的聚集强度逐渐加大,究其原因,这是由于当处于聚集分布格局时,其聚集强度指数K大于零。K>0,K=2S2-,2>0,则S2-必大于0,则S2>。λ=S2-v,将K=2S2-代入λ=2kv式中,则λ=S2-2,因此随着聚块内基本样方数增加,即聚块范围增大,种群密度增大,由于S2>,所以λ值不断增大,聚集强度增大,主要是样方数值的离散性作用结果。
参考文献
[1]王振华,王宏富,刘鑫,等.双斑长跗萤叶甲在相邻农田生态系统中种群消长规律[J].植物保护,2018,44(1):161165.
[2]张汉鹄,谭济才.中国茶树害虫及其无公害治理[M].合肥:安徽科学技术出版社,2004:244250.
[3]张聪,郭井菲,王振营,等.双斑长跗萤叶甲玉米田间成虫数量估计的抽样方法研究[J].植物保护,2013,39(1):7176.
[4]张聪,葛星,赵磊,等.双斑长跗萤叶甲越冬卵在玉米田的空间分布型[J].生态学报,2013,33(11):34523459.
[5]张聪,袁志华,王振营,等.双斑长跗萤叶甲在玉米田的种群消长规律[J].应用昆虫学报,2014,51(3):668675.
[6]王立仁,刘斌侠,付泓,等.玉米田双斑长跗萤叶甲的发生危害情况与防治对策[J].陕西农业科学,2006(2):123131.
[7]李广伟,陈秀琳,张延萍,等.温度对双斑长跗萤叶甲成虫寿命及繁殖的影响[J].昆虫知识,2010,47(2):322325.
[8]袁海滨,齐兴林,孙长东,等.温度对水稻田双斑长跗萤叶甲种群发生动态的影响[J].吉林农业大学学报,2015,37(6):654657.
[9]RUESINK W G. Egg sampling techniques [M]∥KRYSAN J L,MILLER T A,eds.Methods for the study of pest Diabrotica. New York:SpringerVerlag,1986:8399.
[10]PRUESS K P,WEEKMAN G T,SOMERHALDER B R. Western corn rootworm egg distribution and adult emergence under two corn tillage systems [J]. Journal of Economic Entomology,1968,61(5): 14241427.
[11]WEISS M J,MAYO Z B,P N J. Influence of irrigation practices on the spatial distribution of corn rootworm (Coleoptera:Chrysomelidae) eggs in the soil [J]. Environmental Entomology,1983,12(4): 12931295.
[12]FOSTER R E,RUESINK W G,LUCKMANN W H. Northern corn rootworm egg sampling [J].Journal of Economic Entomology,1979,72(5): 659663.
[13]GUSTIN R D. Effect of two moisture and population levels on oviposition of the western corn rootworm [J]. Environmental Entomology,1979,8(3): 406407.
[14]聶强,孙强.双斑长跗萤叶甲成虫的取食选择性研究[J].黑龙江八一农垦大学学报,2009,21(4):3841.
[15]李尚,王振兴,王建盼,等.白毫早和乌牛早茶园卵形短须螨和双斑长跗萤叶甲优势种天敌的差异[J].华南农业大学学报,2016,37(4):238245.
[16]余燕,王振兴,李尚,等.天敌对乌牛早茶园4种害虫空间跟随关系密切程度的年度间差异[J].生态学报,2018,38(13):117.
[17]傅伯杰,陈利顶,马克明,等.景观生态原理及应用[M].北京:科学出版社,2003:216217.
[18]邹运鼎,王弘法.农林昆虫生态学[M].合肥:安徽科学技术出版社,1989:152157.
[19]DAVID F N, MOORE P G. Notes on contagious distribution in plant population [J]. Annals of Botany,1954,18: 4753.
[20]ARBOUS A G, KERRICH J E. Accident statistics and the concept of accidentproneness [J].Biometrics,1951,7(4):340432.
[21]BLACKITH R E. Nearestneighbour distance measurements for the estimation of animal populations [J]. Ecology,1958,39: 147150.
[22]IWAO S.Application of the m*-m method to the analysis of spatial patterns by changing the quadrat size [J]. Researches on Population Ecology,1972,14: 97128.
[23]刘飞飞,柯胜兵,王建盼,等.蜘蛛类天敌与茶尺蠖幼虫空间关系的聚块样方方差分析[J].浙江大学学报(农业与生命科学版),2015,41(2):133146.
[24]王建盼,覃盛,刘飞飞,等.柑橘粉虱与主要捕食性天敌之间的空间关系研究[J].中国生态农业学报,2015,23(4):454464.
[25]李广伟.双斑长跗萤叶甲的生物学、生态学习性及综合防治的研究[D].石河子:石河子大学,2008.
[26]陈静,张建萍,张建华,等.北疆新害虫双斑长跗萤叶甲生物学特性的研究[M]∥成卓敏.植物保护与现代农业,北京:中国农业科技出版社,2007:395397.
(责任编辑:田喆)
