
江苏盐城原生滨海湿地土壤细菌群落多样性分析
2019-08-13 08:54赵婧祝遵凌
江苏农业科学 2019年2期
关键词:群落结构
赵婧 祝遵凌
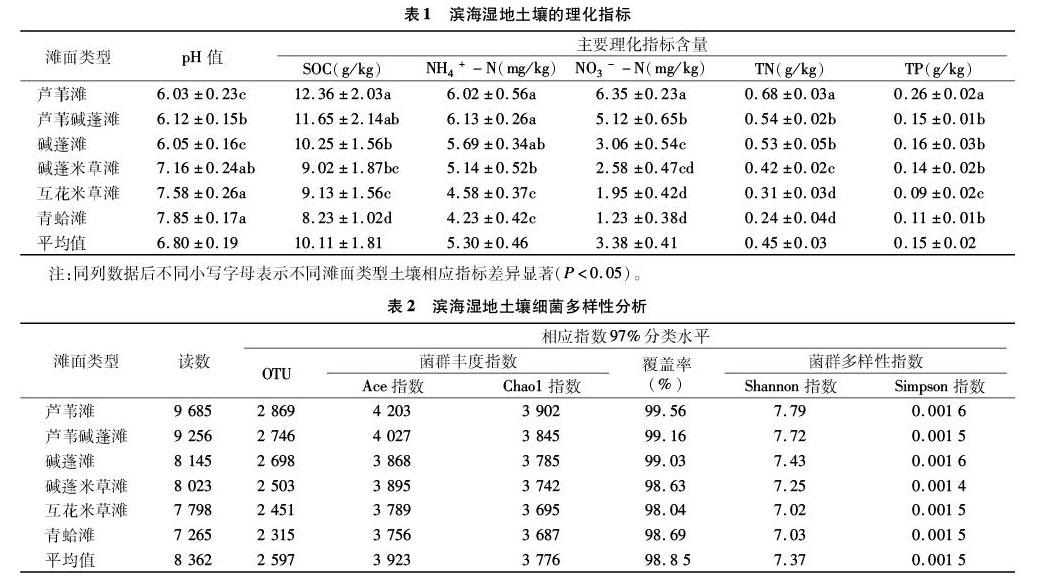
摘要:以江苏盐城原生滨海湿地芦苇滩、芦苇碱蓬滩、碱蓬滩、碱蓬米草滩、互花米草滩、青蛤滩这6种典型滩面为研究对象,采用高通量测序方法测定土壤细菌群落的功能多样性,利用冗余分析细菌群落与环境因子的关系,以探讨滨海湿地土壤细菌群落结构空间分布及环境影响因子。结果表明,滨海湿地土壤理化指标有明显差异,由芦苇滩到青蛤滩,土壤pH值呈逐渐上升趋势,土壤有机碳(SOC)、全氮(TN)、铵态氮(NH+4-N)、硝态氮(NO3--N)含量呈逐渐降低趋势,土壤全磷(TP)含量变化趋势不明显;滨海湿地30个土壤样品高通量测序共得到50 172条高质量序列,平均长度为8 362 bp,有97%相似,土壤样品文库的覆盖率范围为98.04%~99.56%;不同土壤细菌群落丰度指数与Shannon指数排序一致,为芦苇滩>芦苇碱蓬滩>碱蓬滩>碱蓬米草滩>互花米草滩>青蛤滩,丰度相对较高的细菌门为变形菌门、放线菌门、酸杆菌门,丰度值分别为27.26%~33.06%、23.47%~29.03%、15.03%~19.02%,主要的优势菌纲为α-变形菌纲、放线菌纲、酸杆菌纲;由芦苇滩到青蛤滩,土壤细菌Chao、Ace、Shannon指数、OTU数量呈逐渐降低趋势,土壤细菌覆盖率、Simpson指数没有明显规律性变化;土壤pH值、SOC含量、NH4+-N含量是滨海湿地土壤细菌群落结构和多样性的主要影响因子。
关键词:滨海湿地;土壤细菌;群落结构;江苏盐城;芦苇滩;青蛤滩
中图分类号: S154.38+1 文献标志码: A 文章编号:1002-1302(2019)02-0258-04
整个生态圈中,陆地和江河湖泊占据了大部分生态区域,而湿地作为二者的过渡地带,其独特的生态环境和生物群落分布使之备受关注,处于水、土交界的地带存在着较强的生物生产力,对碳、氮等元素的转化、循环起着重要的促进作用,成为地表元素运动的重要途径之一[1-3]。微生物作为重要的生物群落之一,是湿地物质能量交换、湿地生态的维系者,在促进湿地生态系统正常运转方面起着不可忽视的作用[4]。微生物生长及作用的发挥受环境的影响较大,从群落结构、生长状况等研究微生物和环境之间的关系,可为湿地元素的转化研究奠定基础,也有利于对湿地的管理和合理开发利用[5-6]。近年来,分子生物学发展迅速,高通量测序技术因其低成本、无需克隆、通量高等独特优势,在短时间内可深入研究微生物的群落特征,甚至稀有物种的生长发育状况[7-10]。
保持土壤肥力和活性是土壤保持生命活力的前提,而土壤微生物在肥力保持方面起着不可忽视的作用,不仅能够使腐殖质等迅速分解,转化为土壤养分来源,而且能够疏松土壤、增强土壤孔隙度等,对有机化合物的分解、碳氮循环等起着无可替代的作用,同时,能够促进能量的转化循环[11]。有研究表明,微生物作为土壤质量的重要指标之一,其多样性和活性对土壤肥力起着重要的作用[12-13];细菌作为微生物群落的重要组成部分,占微生物类群的70%,且遗传多样性较为丰富,能促进土壤有机质分解及大量营养物质释放,在碳、氮循環过程中起着重要的促进作用,参与整个物质循环和能量交换[14-16]。
我国面积最大、生物多样性明显的淤泥质滨海湿地为盐城滨海湿地。20世纪80年代,江苏人民在沿海进行大规模围垦,全省近70%的围垦集中在盐城地区。随着人为干预的增强,滨海湿地发生巨大变化,其土壤类型和物种多样性也发生巨大变化[17]。李龙山等研究发现,滨海地区植物交替明显,景观呈现典型的动态变化,土壤覆被类型也在不断变化[18]。但是,湿地微生物群落经历什么变化,这些变化与土壤之间关系怎样,如何合理利用滨海湿地资源等等,有关这些研究相对较少。本研究从盐城滨海湿地出发,探究微生物特点与土壤之间的关系,分析细菌多样性对土壤带来的影响、作用机制及其微生物群落的生长发育特点,为盐城滨海湿地土壤资源的合理开发利用提供有益参考。
1 材料与方法
1.1 研究区概况
江苏省盐城滨海湿地属典型的季风气候特征,受到大陆气候和海洋季风的影响,具有较为独特的气候特点,年均降水量近1 000 mm,最冷月份在1月,平均气温约为3 ℃,最热月集中在6—8月,平均气温达到30 ℃左右,9月至翌年2月并不寒冷。盐城滨海湿地核心区地处新洋河和斗龙河过渡区,该区域受到人为的干扰并不明显,依然保持较为完好的生态环境,呈现典型的滨海生态特点,具有差异明显的分带性,能从不同层次反映湿地的生态环境,从陆地向滨海延伸分布着芦苇滩、碱蓬滩、互花米草滩,同时还因青蛤广布而形成青蛤滩,对湿地研究具有很好的代表性。
1.2 土样采集
于2016年11月2日枯水期,采用“S”形5点取样法,分别从滨海湿地分层地带采集0~10 cm土层的土壤,一部分用于测定土壤理化性状,另一部分提取DNA用于测序。
1.3 测定内容和方法
1.3.1 土壤理化性状 分别采用氯化钾溶液提取-分光光度法(HJ 634—2012)、碱熔-钼锑抗分光光度法(HJ 632—2011)测定土壤总氮(TN)、总磷(TP)含量;采用氯化钾溶液提取-分光光度法(HJ 634—2012)测定铵态氮(NH4+-N)、硝态氮(NO3--N)含量。
1.3.2 细菌16S rRNA基因测序 采用Omega试剂盒、引物F27提取16S rDNA宏基因组,采用ABI Gene Amp 9700型PCR仪进行扩增,试验采用Trans Gen AP 221-02,20 μL反应体系为:5×fast pfu buffer 4 μL,2.5 mmol/L dNTPs 2 μL,5 μmol/L 正反向引物各0.8 μL,Fast Pfu聚合酶0.4 μL,TemplateDNA 10 ng,加H2O至20 μL。反应参数为:95 ℃预处理210 s; 55 ℃ 30 s,72 ℃ 45 s,27次循环;72 ℃延伸 10 min。采用AxyPrep DNA凝胶回收,进行Tris_HCl洗脱处理,电泳检测。采用Illumina MiseqPE 300平台测序。
1.4 生物信息处理
对原始DNA通过Mothur软件过滤处理,去除其中的嵌合体;以97%相似性为依据划分可操作分类单元;通过稀释性曲线分析获取丰度、覆盖度、多样性指数,通过主成分分析获取运算分类单元(OTU)相似性;采用贝叶斯算法,获取不同分类水平上的样品群落特点,整理OTU列表获得分类信息与丰度,在门和纲分类水平下进行冗余分析(RDA),得到细菌群落组成结构及其与环境因子的关系。
2 结果与分析
2.1 滨海湿地土壤理化指标
由表1可知,滨海湿地土壤pH值在6.03~7.85之间,平均值为6.80,由芦苇滩到青蛤滩,土壤pH值呈逐渐增加趋势;土壤有机碳(SOC)含量在8.23~12.36 g/kg之间,平均值为10.11 g/kg,由芦苇滩到青蛤滩,SOC呈逐渐降低趋势;从芦苇滩到青蛤滩,NH4+-N、NO3--N含量呈逐渐降低趋势,在芦苇滩出现最大值,分别为6.02、6.35 mg/kg,NH4+-N、NO3--N平均含量分别为5.30、3.38 mg/kg;TN含量变化趋势与SOC相一致,从芦苇滩到青蛤滩呈逐渐降低趋势,其平均值为0.45 g/kg;TP含量变化趋势不明显,总体上呈递减趋势,其平均值为0.15 g/kg。
2.2 滨海湿地土壤细菌的多样性
由表2可知,滨海湿地30个土壤样品的高通量测序共得到50 172条高质量序列,平均长度为8 362 bp,97%相似;各样品文库的覆盖率为98.04%~99.56%,说明土样中基因序列被检出的概率相对较高,测序结果能够代表湿地土壤细菌群落的真实情况。Chao1指数用于估算样品中所含OTU总数,反映菌群丰度,Chao1指数越大,说明细菌群落丰度越高;Shannon指数反映细菌群落alpha多样性指数,Shannon指数值越大,说明细菌群落多样性越高。由表2可知,不同土壤细菌群落丰度指数与Shannon指数排序一致,为芦苇滩>芦苇碱蓬滩>碱蓬滩>碱蓬米草滩>互花米草滩>青蛤滩,从芦苇滩到青蛤滩,土壤细菌Chao1、Ace、Shannon指数、OTU数量呈逐渐降低趋势,土壤细菌覆盖率、Simpson指数变化没有明显规律性变化。
2.3 滨海湿地土壤细菌门水平分类和纲水平分类
2.3.1 门分类水平 由图1可知,沿湖面至坡地,土壤的门分类细菌相对丰度存在一定的变化趋势,滨海湿地土壤相对丰度较高的细菌门为变形菌门、 放线菌门、酸杆菌门、绿弯菌门、芽单胞菌门、厚壁菌门、绿菌门、硝化螺旋菌门,相对丰度值分别为27.26%~33.06%、23.47%~29.03%、15.03%~19.02%、7.13%~13.58%、5.13%~9.87%、2.16%~5.12%、1.54%~4.23%、0.66%~4.87%;由芦苇滩到青蛤滩,变形菌门相对丰度大致呈先增大后减小趋势,酸杆菌门大致呈增大趋势,绿弯菌门大致呈先减小后增大趋势,绿菌门没有明显变化规律。
2.3.2 纲分类水平 由图2可知,滨海湿地土壤检测到的主要纲有α-變形菌纲、放线菌纲、酸杆菌纲、β-变形菌纲、嗜热油菌纲、芽单胞菌纲、杆菌纲、δ-变形菌纲,相对丰度值分别为26.06%~29.78%、18.30%~29.30%、15.02%~21.05%、8.26%~15.06%、5.32%~8.32%、3.12%~8.30%、1.45%~8.14%、1.21%~7.27%,其中,前6个纲在土壤中占主导地位,约占所有纲的80%。
2.4 滨海湿地土壤理化性质与土壤细菌群落结构的相关性
土壤可以提供细菌群落生长繁殖的微环境,而不同的植被类型可通过改变土壤的微环境间接影响土壤细菌群落结构的组成。由表3可知,变形菌门、酸杆菌门、厚壁菌门相对丰度与土壤pH值呈显著负相关(P<0.05),绿弯菌门相对丰度与土壤pH值呈极显著负相关(P<0.01);变形菌门、酸杆菌门、芽单胞菌门、厚壁菌门、硝化螺旋菌门相对丰度与SOC含量呈显著正相关,绿弯菌门相对丰度与土壤SOC含量呈极显著正相关;酸杆菌门、芽单胞菌门、厚壁菌门、硝化螺旋菌门相对丰度与NH4+-N含量呈显著正相关,变形菌门、放线菌门相对丰度与NH4+-N含量呈极显著正相关;变形菌门、酸杆菌门、绿弯菌门、绿菌门、绿菌门相对丰度与NO3--N含量呈显著正相关,芽单胞菌门相对丰度与NO3--N含量呈极显著正相关;变形菌门、绿弯菌门、厚壁菌门、绿菌门相对丰度与TN含量呈显著正相关,芽单胞菌门相对丰度与TN含量呈极显著正相关;不同细菌门相对丰度与TP含量相关性不显著(P>0.05)。
2.5 滨海湿地环境因素对土壤细菌群落结构的影响
由图3可知,对土壤理化环境因子与细菌主要菌落(门)所占比重进行中心标准化,并进行校正,提取第一、第二坐标轴信息,对方差总解释率分别为61.25%、23.58%;第一主轴上,SOC、NH4+-N含量是主要的影响因子,第二主轴上,SOC含量、pH值是主要的影响因子。因此,与滨海湿地土壤细菌群落相关性较大的土壤理化指标为pH值、SOC含量、NH4+-N 含量。
3 结论与讨论
生物的多样性对维系整个生态平衡具有举足轻重的作用,微生物群落的多样性对促进整个生态循环同样具有重要的意义,不仅有利于营养盐的循环,且在有机物降解、养分转化方面发挥着重要作用[19]。本研究发现,芦苇滩菌群的丰度、多样性相对最高,其次是芦苇碱蓬滩、碱蓬滩,而碱蓬米草滩、互花米草滩的丰度及多样性并不明显,而青蛤滩微菌群分布相对最少;土壤菌群的丰度、多样性、OTU数量等从陆地向滨海地带多呈递降趋势,这可能是距离河岸较远的地方土壤肥力相对较强,对微生物的生长发育有利。对位置相对接近的地带,其土壤菌群分布具有较明显的相似性[20]。有研究发现,微生物的生长发育及活动明显受到水文状况的制约,干湿交替对部分细菌生长具有明显的促进作用,与淹没区有显著不同[21];水位状况及其变化直接影响微生物群落的分布,且水文条件影响下的土壤沉积差异对微生物群落的影响更为显著[22-23]。本研究结论与之较为吻合。
微生物菌群中的变形菌门在黄河三角洲湿地具有较高的丰度[24]。对盐城滨海湿地而言,相对丰度较高的变形菌门α、β、δ-变形菌纲中,前2类菌纲具有较多的固氮细菌,能够明显提升土壤的固氮水平,对土壤肥力的保持具有重要作用,而δ-变形菌纲为食菌性细菌[25],能够明显促进湿地养分循环;酸杆菌门相对丰度值也较高,相对丰度值15.03%~19.02%,与张琼琼等的研究结果[26]基本一致,但其多为未培养细菌,虽然在土壤中广泛存在,但相关研究并不多;试验中菌群相对丰度位于第4位的绿弯菌门,以CO2为能量来源,在光合作用下获取能量,进而使其能够在SOC含量较低的土壤中拥有较强的生存能力和适应性。通过冗余分析发现,SOC含量、NH4+-N含量、pH值对湿地菌群分布具有明显的制约效应。
参考文献:
[1]庞丙亮,崔丽娟,马牧源,等. 若尔盖高寒湿地生态系统服务价值评价[J]. 湿地科学,2014,12(3):273-278.
[2]张 彪,史芸婷,李庆旭,等. 北京湿地生态系统重要服务功能及其价值评估[J]. 自然资源学报,2017,32(8):1311-1324.
[3]魏 强,佟连军,杨丽花,等. 三江平原湿地生态系统生物多样性保护价值[J]. 生态学报,2015,35(4):935-943.
[4]丁冬静,李 玫,廖宝文,等. 海南省滨海自然湿地生态系统服务功能价值评估[J]. 生态环境学报,2015(9):1472-1477.
[5]李 伟,崔丽娟,庞丙亮,等. 湿地生态系统服务价值评价去重复性研究的思考[J]. 生态环境学报,2014,23(10):1716-1724.
[6]张 晶,陈书明,王小国. 天鹅湖湿地微生物群落PCR-DGGE分析[J]. 基因组学与应用生物学,2016(9):2423-2428.
[7]陈 伟. 纳帕海高原湿地微生物多样性研究[D]. 昆明:昆明理工大学,2016.
[8]鲁青原. 辽河三角洲滨海湿地微生物群落组成及其环境意义[D]. 北京:中国地质大学,2016.
[9]常军军,吴苏青,梁 康,等. 复合垂直流人工湿地微生物特征对典型污水的响应差异[J]. 环境科学研究,2016,29(8):1200-1206.
[10]黄兴如,张琼琼,张瑞杰,等. 再生水补水对河流湿地香蒲根际细菌群落结构影响研究[J]. 中国环境科学,2016,36(2):569-580.
[11]王 林,李 冰,朱 健. 高通量测序技术在人工湿地微生物多样性研究中的研究进展[J]. 中国农学通报,2016,32(5):10-15.
[12]吴文卫,刘 昂,谷照虎,等. 采用PCR-DGGE技术研究处理农田退水组合人工湿地微生物群落特征[J]. 应用与环境生物学报,2016,22(6):978-985.
[13]魏佳明,崔丽娟,李 伟,等. 表流湿地细菌群落结构特征[J]. 环境科学,2016,37(11):4357-4365.
[14]李龙山,倪细炉,李昌晓,等. 生活污水对土壤及湿地植物根际细菌群落的影响[J]. 农业环境科学学报,2016,35(11):2163-2170.
[15]王 鹏,陈 波,张 华. 基于高通量测序的鄱阳湖典型湿地土壤细菌群落特征分析[J]. 生态学报,2017,37(5):1650-1658.
[16]隋 心,张荣涛,杨立宾,等. 模拟氮沉降对三江平原小叶章湿地土壤细菌多样性的影响[J]. 草业科学,2016,33(4):589-598.
[17]靳 亮,高学梅,杜建華,等. 尕海湖湿地泥炭细菌多样性分析[J]. 微生物学通报,2016,43(11):2396-2404.
[18]李龙山,倪细炉,李昌晓,等. 生活污水对土壤及湿地植物根际细菌群落的影响[J]. 农业环境科学学报,2016,35(11):2163-2170.
[19]武海鹏. 生态环境修复及江湖关系改变对洞庭湖湿地土壤微生物的影响[D]. 长沙:湖南大学,2016.
[20]徐 飞,蔡体久,杨 雪,等. 三江平原沼泽湿地垦殖及自然恢复对土壤细菌群落多样性的影响[J]. 生态学报,2016,36(22):7412-7421.
[21]靳振江,曾鸿鹄,李 强,等. 起源喀斯特溶洞湿地稻田与旱地土壤的微生物数量、生物量及土壤酶活性比较[J]. 环境科学,2016,37(1):335-341.
[22]周雪芳,王立立,刘 铃,等. 珠江河口湿地沉积物中耐盐硝化细菌的筛选及其河口硝化特性研究[J]. 生态环境学报,2016,25(12):1991-1998.
[23]黄 媛,褚文珂,谢蔚鹏,等. 杭州西溪湿地沉积物中不同培养基分离的细菌种群多样性[J]. 杭州师范大学学报(自然科学版),2016,15(6):600-605.
[24]曾 静. 锡林河湿地厌氧氨氧化菌群多样性、丰度及空间特征研究[D]. 呼和浩特:内蒙古大学,2016.
[25]张瑞杰,张琼琼,黄兴如,等. 再生水湿地香蒲根内生细菌群落多样性及其水质特征分析[J]. 中国环境科学,2016,36(3):875-886.
[26]张琼琼,黄兴如,郭逍宇. 基于T-RFLP技术的不同水位梯度植物根际细菌群落多样性特征分析[J]. 生态学报,2016,36(14):4518-4530.
猜你喜欢
热带农业科学(2016年11期)2017-01-21
热带农业科学(2016年11期)2017-01-21
河北渔业(2015年12期)2015-12-21
河北渔业(2015年11期)2015-11-18
河北渔业(2015年11期)2015-11-18
河北渔业(2015年10期)2015-10-15
河北渔业(2015年7期)2015-07-21
河北渔业(2015年6期)2015-06-24
河北渔业(2015年1期)2015-01-19
