
土壤铝离子对香蕉苗期生长及根系活性的影响
2019-08-26 01:24陈俊铭张宇陈小丽李进
南方农业·下旬 2019年4期
关键词:香蕉
陈俊铭 张宇 陈小丽 李进

摘 要 为研究铝离子对香蕉生长及根系活性的影响,在6个香蕉盆栽中分别加入不同量的AlCl3·6H20进行试验,并将试验结果进行对比分析。结果表明,土壤中铝离子浓度为0~4 mg·kg-1时,对香蕉生物量累积、根系活力有一定促进作用;当铝离子浓度为4~8 mg·kg-1时,土壤中的交换性铝含量显著上升(31~37 cmol·kg-1),土壤pH则显著下降,香蕉受铝害影响,其生物量累积和根系活性均被土壤中的铝离子抑制。
关键词 香蕉;根系活性;交换性铝
中图分类号:S156 文献标志码:B DOI:10.19415/j.cnki.1673-890x.2019.12.071
目前,由于汽车尾气排放、酸雨、长期不合理施用化肥等导致的土壤酸化加剧以及土壤酸化引起的植物铝毒害问题严重制约着现代农业生产的可持续发展,已成为全球性的农业环境保护问题。据统计,酸性土壤占据了全球耕地总面积的40%,且受害面积还在逐步增加,土壤酸化问题已经到了必须解决的时候[1]。同样,分布于我国南方区域15个省份的酸性土壤总面积约有203万平方千米,占全国总耕地面积的21%左右[2]。随着我国集约化农业的快速发展,长期不合理施用化肥导致我国南方耕地土壤酸化及铝毒危害加剧,这已经成为我国南方地区农业生产的主要限制因素。
在自然条件下,铝在土壤中多以难溶性的硅酸盐或者氧化铝的形式存在,如铝土矿、长石、高岭石、云母等,但随着土壤pH的降低,土壤中的铝则以化学形态存在。尤其当pH<5时,土壤中的铝主要以Al3+为主,其微克级离子浓度会对很多作物产生毒害作用,故此铝胁迫是限制作物生长发育的主要障碍因子[3]。
湛江市位于粤西地区,其代表土壤为酸性较强的砖红壤,是铝毒灾害高发地区。同时,湛江又是广东省主要的香蕉主产区,香蕉生产受土壤酸度及铝毒害影响严重。基于此,以湛江市典型砖红壤和威廉姆8号香蕉为供试材料,研究了不同铝离子浓度处理对香蕉生长、根系性状以及对土壤中交换性铝含量和pH的影响,以期为南方酸性耕地的铝毒害防控提供理论依据。
1 材料与方法
1.1 供试作物
供试作物为威廉姆8号香蕉,在真叶期取三片长势一致的香蕉幼苗,将其移栽于塑料盆中,每盆种植1株幼苗。
1.2 供试土壤
供试土壤为广东湛江广东海洋大学后山香蕉地中的砖红壤,此香蕉地种植香蕉3年以上,土壤pH为5.3,经测定土壤的基础理化性质如表1所示。采集土壤后,挑出土壤中的石块、枯枝落叶等,磨碎,过2 mm的筛,密封保存备用。
1.3 试验设计
2017年4月27日在广东海洋大学生物技术研究所温室大棚进行盆栽试验,按每千克土中加入铝的量设置处理,分别为0 mg·kg-1、0.5 mg·kg-1、1.0 mg·kg-1、2.0 mg·kg-1、4.0 mg·kg-1、8.0 mg·kg-1共6个铝浓度水平,依次记为A0、A1、A2、A3、A4、A5,每个处理有4个重复,共计24盆香蕉。
试验每盆装有3.0 kg的供试土壤,且每盆分别添加1.02 g过氯化钾、1.27 g过磷酸钙、1.26 g尿素作为基肥,充分混匀。2017年4月27日移栽供试香蕉到盆栽中,待香蕉生长2个月后,开始按处理设置加入铝离子。即分别将14.4 mg、26.8 mg、53.7 mg、107.4 mg、214.8 mg的AlCl3·6H20(鋁含量为11.18%)加入到100 mL容量瓶中,定容成100 mL的铝溶液,并分别缓慢均匀的加入到A1、A2、A3、A4、A5处理中,对照处理则加入同样体积的蒸馏水。培养香蕉8周后收获香蕉,采集土壤样品,并测定植株生物量、根系性状以及土壤的交换性铝含量和pH等相关指标。
1.4 样品分析
参考《土壤农化分析》(鲁如坤,1999),采用pH计电位法测定土壤pH(按2.5∶1的水土体积质量比),采用氯化钾淋洗-碱滴定法测定交换性Al3+,采用重铬酸钾容量法测定土壤有机质含量,采用碱解扩散法测定土壤中碱解氮的含量,用Olsen法测定土壤中速效磷的含量,采用乙酸铵浸提-火焰光度计法测定土壤中的速效钾含量[4]。
试验结束时采集植株根部和地上部,根部鲜样用于测定根系活性和丙二醛,剩余样品用烘箱在105 ℃下杀青,最后在75 ℃下烘干至恒重,分别称量地上部和根部干重,并计算根冠比;采用氯化三苯基四氮唑(TTC)法测定香蕉的根系活力[5];采用硫代巴比妥酸(TBA)法测定根系的丙二醛(MDA)含量[6]。
1.5 数据统计分析方法:
利用SPSS 18.0、Microsoft Excel 2010进行数据分析统计。
2 结果与分析
2.1 不同浓度铝对香蕉植株的生物量和根冠比的影响
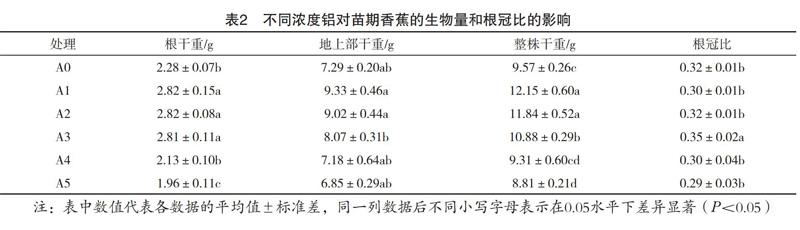
生物量是描述植物生长状况的最直接指标。不同浓度铝对香蕉植株的生物量和根冠比的影响见表2,随着铝浓度的上升,植株的地上部、根重、整株重、根冠比均呈先升高后降低趋势。生物量最高的处理为A1,其根干重和整株干重分别比对照处理A0增加了23.7%和27.0%;而A5处理的根系干重及整株干重均显著下降,相较对照A0分别下降了14.0%、7.9%;但A4处理的生物量与对照A0的差异不显著,说明加入的铝溶液浓度小于A4处理时,铝离子对香蕉的生长无明显的影响,甚至有一定的促进作用。而当加入的铝溶液浓度大于A4处理时,过高浓度的铝离子会对香蕉的根系造成毒害,抑制香蕉根系和整株增重。
根冠比是指植物根部与地上部的干重比值。在植物苗期,较高的根冠比能够给植物提供良好的营养生长条件,会增加根系营养物质的吸收面积和能力[7],因此,植物根冠比的增加是对植物有利的。如表2所示,香蕉的根冠比随铝浓度水平的增加呈先增大后减小的趋势,A3处理的根冠比显著大于其他处理。
2.2 不同浓度铝对香蕉根系活力及丙二醛含量的影响
在逆境条件下,植物的根系会受到损伤,致使根系活力下降,从而导致根系对水分和无机盐的吸收能力下降,最终使整个植株的生长发育受到抑制。如表3所示,香蕉的根系活力变化随土壤中铝浓度的增加呈先上升后又下降趋势。处理A2和A3香蕉的根系活力比对照香蕉的根系活力分别增长了21.0%、20.2%,而处理A5香蕉的根系活力比对照香蕉的根系活力降低了25.5%。
植物在逆境胁迫或衰老过程中、在细胞内活性氧代谢的平衡被破坏的情况下,对植株活性氧的积累十分有利。活性氧大量累积,膜脂的过氧化作用就会有所加强,进而会损坏细胞膜系统,最终导致植物无法生长而死亡。丙二醛(MDA)是膜脂过氧化的最终产物,其产生之后会加剧对细胞膜的损害,使细胞膜降低甚至失去选择透过性,进而引起细胞内的电解质外渗,导致细胞内的稳态环境遭到破坏[8],因此,丙二醛含量经常被用于植物衰老生理和抗性生理的研究中。如表3所示,随着铝含量的增加,香蕉根系中的MDA含量呈降低趋势,从A0到A4均有小幅度下降,但处理间差异不显著;当铝的加入量为A5时,香蕉根系的MDA含量较对照A0上升了104.0%,且A5处理香蕉根系中的MDA含量显著大于其他处理。
2.3 不同浓度铝对土壤中交换性铝含量和pH的影响
从表4可以看出,随着铝浓度的增加,土壤中的交换性铝含量也显著增加,相对于对照组土壤中的交换性铝,A5处理的交换性铝含量增加了134.9%。相反,土壤pH随着铝浓度的增加有所下降,高铝浓度A5和A6处理的土壤pH下降明显,显著小于A0、A1、A2、A3处理。
3 结论与讨论
3.1 讨论
3.1.1 不同浓度铝对香蕉植株生物量的影响
低浓度(0~4 mg·kg-1)的铝处理对于香蕉幼苗的根系生长具有一定程度的促进作用(表2),在此浓度内,可能造成香蕉根系的质膜透性增加但不会致其完全失去生物活性,从而使得各种物质进出细胞的能力增加,使根系对水分和各种营养物质吸收的能力增强[9];但也有可能是由于低浓度的铝促进了根系细胞的分裂分化[10],从而促进了根系生物量的增加。
3.1.2 不同浓度铝对香蕉根系活力及丙二醛含量的影响
研究结果(表3)显示,在A2~A4可能存在1个铝浓度临界值,小于此临界值,香蕉的根系活力会随着铝浓度的上升而上升;而当超过这个临界值时,香蕉根系的抗逆性到达极限,铝胁迫开始对香蕉造成伤害,进而导致了根系活力的下降。刘鹏等[11]在相关研究中也发现大豆、木豆等植物存在类似的铝浓度与根系活力的关系。
试验表明(表3),在低浓度铝条件下(0~4 mg·kg-1),MDA的积累量相对较低,不会对香蕉根系的质膜有太大影响,此时香蕉根系的细胞膜能够维持正常的选择透过性,使膜脂过氧化作用处于较低水平。但随着铝浓度的增大,香蕉根系产生了大量的MDA,对根系质膜损伤加大,使膜脂过氧化作用增强,导致质膜透性增大,使香蕉的各项生长指标(如生物量、根系活力等)受到严重抑制。纪雨薇[12]的相关研究也表明,马尾松在较高铝浓度时,其根系MDA含量达到最高值,此时马尾松的根系结构会遭到严重破坏,引起根系活力下降。
3.1.3 不同浓度铝对土壤中交换性铝含量和pH的影响
土壤中的交换性铝含量与所加入铝溶液的浓度呈正相关性,当铝的加入量达到A4水平,即土壤交换性铝含量为31 cmol·kg-1时,香蕉的生物量和根系活力均开始显著降低。因此,建议通过测定土壤中的交换性铝含量来确定某地区的铝毒危害的临界浓度,当铝离子含量达到临界水平时,才会对作物生长产生铝毒危害,这时才是改良土壤酸度,减轻作物铝毒危害的最佳时机,避免了大量或不合理施用石灰类土壤调理剂所引起的土壤板结、复酸化、重金属污染等一系列问题。
3.1.4 土壤中交换性铝含量和pH与香蕉生物量的关系
由表2和表4可以看出,在较低pH和较高交换性铝含量的情况下,香蕉植株的生物量显著降低。梁文君等[13]也发现在土壤pH低于5.5时,玉米的生物量明显降低。因此,在酸性土壤中,植株生长会受制于土壤pH、交换性铝等致酸离子[14]
3.2 结论与讨论
铝离子的加入量在0~4 mg·kg-1时,即土壤交换性铝含量为15~31 cmol·kg-1时,其对香蕉苗的生物量、根系活力无明显影响,或有一定促进作用。铝离子的加入量在4~8 mg·kg-1时,其土壤中的交换性铝含量显著上升(31~37 cmol·kg-1),土壤pH則显著下降,且香蕉苗受铝胁迫,生物量和根系活性均显著降低。说明当铝离子的加入量超过4 mg·kg-1时,即土壤交换性铝含量大于31 cmol·kg-1时,香蕉苗生长开始受到铝离子的抑制,建议施用适量的石灰类酸性土壤调理剂来减少土壤铝毒危害。
参考文献:
[1] Kochian L V. Cellular Mechanisms of Aluminum Toxicity and Resistance in Plants[J]. Annual Review of Plant Physiology & Plant Molecular Biology, 1995,46(1):237-260.
[2] 周蓉.花生耐铝性及遗传改良研究进展[J].花生学报,2003,32(Z1):144-148.
[3] 周蓉,廖伯寿,陈小媚,等.铝胁迫对花生根系的影响[J].花生学报,1998(4):1-4.
[4] 鲁如坤.土壤农业化学分析方法[M].北京:中国农业科技出版社,1999.
[5] Clemensson-Lindell A. Triphenyltetrazolium chloride as an indicator of fine-root vitality and environmental stress in coniferous forest stands: Applications and limitations[J]. Plant & Soil,1994,159(2):297-300.
[6] 王学奎.植物生理生化实验原理和技术[M].北京:高等教育出版社,2006.
[7] 马祥庆,梁霞.植物高效利用磷机制的研究进展[J].应用生态学报,2004(4):712-716.
[8] 季杨,张新全,彭燕,等.干旱胁迫对鸭茅根、叶保护酶活性、渗透物质含量及膜质过氧化作用的影响[J].草业学报,2014,23(3):144-151.
[9] 王水良,王平,王趁义.铝胁迫对马尾松幼苗根系形态及活力的影响[J].生态学杂志,2010,29(11):2097-2101.
[10] 阮建云,王国庆,石元值,等.茶园土壤铝动态及茶树铝吸收特性[J].茶叶科学,2003(1):16-20.
[11] 刘鹏,YangY.S.,徐根娣,等.铝胁迫对大豆幼苗根系形态和生理特性的影响[J].中国油料作物学报,2004(4):51-56.
[12] 纪雨薇.马尾松铝胁迫生理响应机制[D].重庆:西南大学,2016.
[13] 梁文君,蔡泽江,宋芳芳,等.不同母质发育红壤上玉米生长与土壤pH、交换性铝、交换性钙的关系[J].农业环境科学学报,2017,36(8):1544-1550.
[14] Hoyt P B, Nyborg M. Toxic Metals in Acid Soil: I. Estimation of Plant-Available Aluminum1[J]. Soil Science Society of America Journal, 1971,35(2):114-118.
(责任编辑:赵中正)
猜你喜欢
小天使·一年级语数英综合(2021年4期)2021-05-08
小天使·一年级语数英综合(2019年10期)2019-11-10
家教世界·V家长(2019年3期)2019-06-11
小学阅读指南·低年级版(2018年5期)2018-11-02
小学阅读指南·低年级版(2017年12期)2017-12-26
小天使·三年级语数英综合(2017年11期)2017-12-05
阅读与作文(小学高年级版)(2017年10期)2017-10-11
作文大王·低年级(2016年7期)2016-08-09
幼儿教育·父母孩子版(2016年4期)2016-07-25
