
2,4-D处理对大百合种子休眠解除及内源激素变化的影响
2019-08-30 07:07李凤荣李叶芳马诗雨关文灵
西南林业大学学报 2019年5期
李凤荣 李叶芳 马诗雨 关文灵
( 云南农业大学园林园艺学院,云南 昆明 650201)
大百合(Cardiocrinum giganteum)是百合科(Liliaceae)大百合属(Cardiocrinum)多年生鳞茎类草本植物[1],该属植物因植株巨大并具有网状脉的叶片而显著区别于百合属(Lilium)植物[2]。大百合植株粗壮挺拔,叶片硕大,花朵洁白芳香,具有极高的观赏价值,其鳞茎富含多种营养物质,可供食用[3],也可药用[4];果实可入药,作为马兜铃(Aristolochia debilis)的代用品[5],是优良的野生植物资源。
国内关于大百合的研究主要集中在其引种栽培[6]、组织培养[7]、细胞水平[8]以及杂交[9]等方面。而国外鲜有对大百合属的研究,主要在其种子生物学[10]、生活史[11]、花粉发育[12]等方面。关文灵等[13]的研究表明大百合种子存在形态和生理后熟现象,其休眠类型属于复杂的形态生理休眠。蔡薇等[14]研究变温层积过程中加赤霉素(GA3)、6-苄氨基腺嘌呤(6-BA)、硝普钠(SNP)处理对大百合种胚后熟的影响。2,4-二氯苯氧乙酸(2,4-D),是一种人工合成的植物生长素,具有促进细胞生长与分化,加速根、芽伸长,促进果实形成与种子萌发等作用[15]。目前,鲜见2,4-D浸种处理对大百合种子休眠解除的研究,也未见内源激素在大百合种子变温层积过程中变化的研究。本研究采用2,4-D浸种处理结合变温层积的方法,探究对大百合种子休眠破除的影响,筛选出适宜的浓度,在最佳处理浓度的基础上,研究大百合种子在不同时期内源激素的动态变化,为揭示内源激素在大百合种子休眠解除过程中的作用提供参考,为种子育苗生产实践提供科学依据。
1 材料与方法
1.1 试验材料
试验所用大百合种子,于2015年11月采自云南省哀牢山国家级自然保护区(东经100°44′~101°30′,北纬 23°36′~24°56′),在 4 ℃ 低温环境下储存备用。在变温层积结合激素处理的不同时期,对大百合种子取样,取样时间为层积后 0、30、60、90、120、130 d,在各时间点定期取样,样品经液氮冷冻后,置于-80 ℃超低温冰箱保存,用于测定种胚发育过程中植物内源激素。
1.2 试验方法
1.2.1 预处理方法
以蒸馏水为对照(CK),将大百合种子置于质量浓度分别为1、5、10 mg/L的2,4-D溶液中浸泡48 h;每个处理种子150粒,设置3个重复。将上述浸种的种子装入尼龙网袋中,再用潮湿的苔藓覆盖网袋,最后放入自封袋中,并在自封袋上用牙签戳 3~4个小孔以保持透气性。在恒温箱中进行模拟夏季—秋季—冬季的高温到低温的变温 25 ℃/15 ℃( 60 d)、15 ℃/5 ℃( 60 d)、4 ℃(50 d)层积处理,周期性光照(光照12 h,黑暗12 h)。
1.2.2 指标测定方法
1)胚长、胚生长率和发芽率的测定 在变温层积90、105、120、135 d时,每个处理中随机抽取10粒种子,在体视显微镜(徕卡公司,德国)下沿着与胚平行的方向,测量胚的长度。在变温层积150 d时,随机选取100粒,按公式(1)计算胚生长率,共重复3次。当胚根突出珠孔1 mm时,种子视为已萌发,观察并记录种子发芽起始时间。每隔5 d,统计100粒种子中的发芽率,并按公式(2)计算。

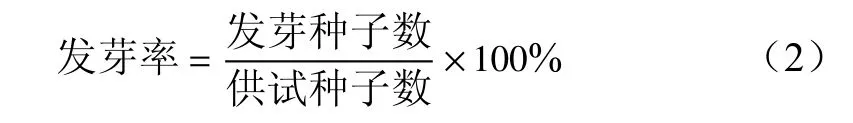
2)内源激素含量的测定 在变温层积期间,定期对处理组(5 mg/L 2,4-D)和CK浸泡48 h的种子分别取样,经液氮冷冻后,保存于-80 ℃超低温冰箱中;用液-质联用色谱仪(安捷伦,美国)测定GA3、生长素(IAA)及脱落酸(ABA)的内源激素含量,具体方法参照参考文献[16]。
1.3 数据分析
采用SPSS 19.0对种胚长度、胚生长率进行单因素ANOVA的Duncan多重比较方法进行显著性差异分析,对内源激素ABA、IAA和GA3含量进行单样本T检验分析,使用Excel 2013进行数据统计和图表制作。
2 结果与分析
2.1 不同浓度2,4-D浸种对大百合种胚长度的影响
由图1可知,随着层积时间的增加,不同处理大百合种胚的长度均增长,但在层积90 d时,不同处理间胚长差异不显著;层积105 d时,CK与5 mg/L 2,4-D处理差异显著(P<0.05),与10 mg/L 2,4-D处理差异极显著(P<0.01),CK的胚长分别高出5 mg/L 2,4-D和10 mg/L 2,4-D的胚长8.38%、14.74%。层积120 d时,CK与其他处理差异显著(P<0.05),与5 mg/L和10 mg/L 2,4-D显著极差异(P<0.01),CK的胚长分别高出5 mg/L 2,4-D和10 mg/L 2,4-D的胚长15.28%、15.88%。层积135 d时,CK与5 mg/L 2,4-D处理显著差异(P<0.05),5 mg/L 2,4-D处理的胚长高出CK的胚长9.03%。

图 1 不同浓度2,4-D处理下大百合种胚的长度变化Fig. 1 Length changes of C. giganteum embryos under different concentrations of 2,4-D
2.2 不同浓度2,4-D对大百合种子发芽的影响
由表1可知,CK的胚生长率与5 mg/L 2,4-D和 10 mg/L 2,4-D差异极显著(P<0.01);5 mg/L 2,4-D处理的胚生长率最高,达到82%,比CK高出10%;10 mg/L 2,4-D处理的胚生长率最低,为52.33%,比CK低19.67%。5 mg/L 2,4-D的发芽起始时间最早,为4 ℃下14 d,比CK早11 d。

表 1 大百合种子胚生长率和发芽时间Table 1 Seed embryo growth rate and germination time of C. giganteum
由图2可知,1 mg/L 2,4-D处理下,种子发芽率高于CK 3%,5 mg/L 2,4-D的发芽率最高,达75.33%,高于CK 15.33%,可促进大百合种子的萌发;10 mg/L 2,4-D的发芽率最低,为36.33%,明显低于CK,低了30%。结合来看,5 mg/L 2,4-D处理的胚生长百分比最高,发芽时间最早,发芽率最高,利于大百合种胚后熟,可以缩短发芽时间,提高种子萌发。
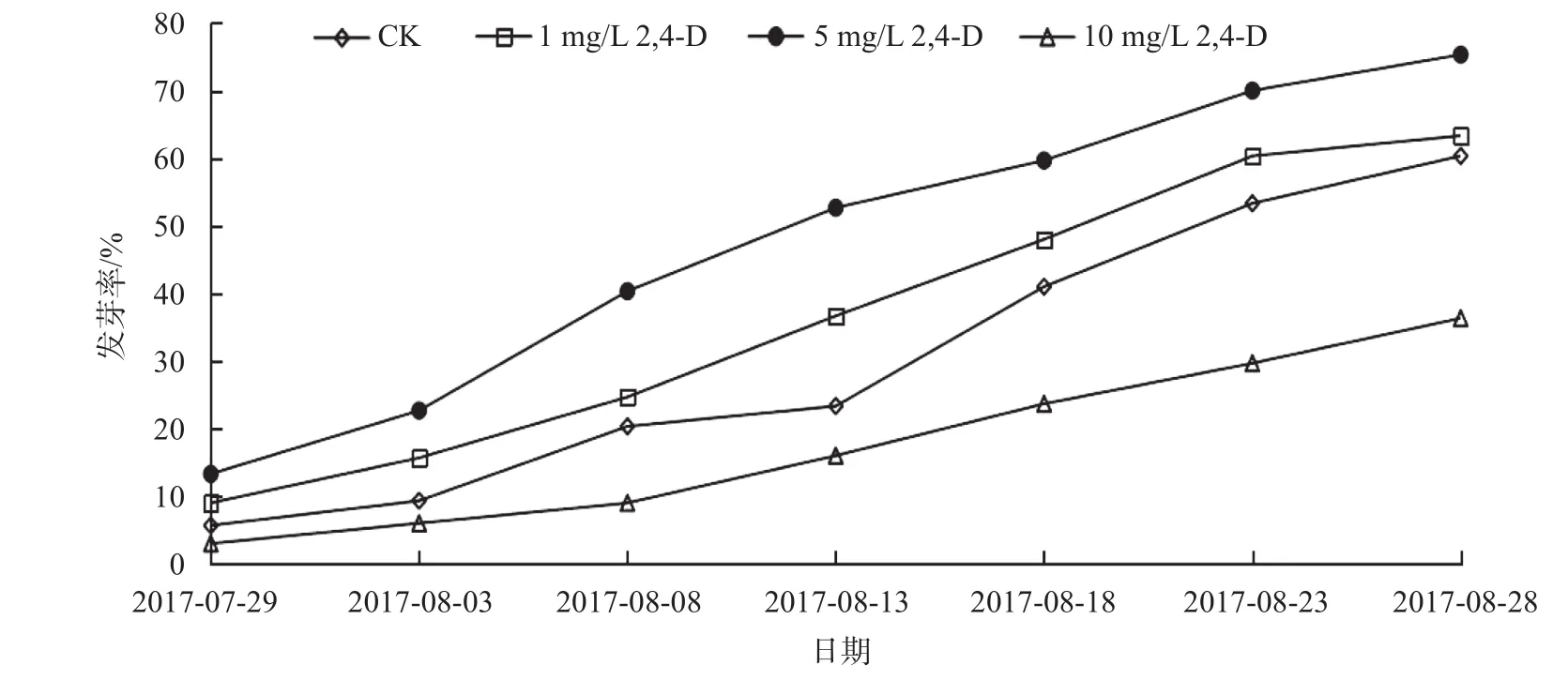
图 2 不同浓度2,4-D处理下大百合种子的发芽率Fig. 2 Germination rate of C. giganteum seeds treated with 2,4-D with different concentrations
2.3 大百合种子层积过程中内源激素的动态变化
随着层积时间的增加,在层积过程中,大百合种子ABA、IAA和GA3含量的变化表现出不同的规律,见图3。

图 3 大百合种子层积过程中内源激素含量的变化Fig. 3 Variation of endogenous hormones content in the process of C.giganteum seeds stratification
由图3a可知,在大百合种子层积过程中,CK和2,4-D处理在ABA含量的变化总体呈下降趋势。在未变温层积0 d时差异极显著(P<0.01),2,4-D处理比 CK高了 14.53 ng/g;在层积30 d时差异极显著(P<0.01),2,4-D处理的比CK低了9.55 ng/g;在层积120 d时差异极显著(P<0.01),CK比2,4-D低了5.59 ng/g;在层积60、90、130 d时,处理差异不显著。CK中ABA含量在层积60 d时,下降速度较快,层积30 d相比,降了14.1 ng/g,在层积120 d时达到最低水平9.82 ng/g;而2,4-D处理的ABA含量在层积30 d时,急速下降,降了31.69 ng/g,在层积60 d时达到最低水平14.14 ng/g。表明2,4-D 5 mg/L处理的ABA含量下降速度的时间比CK早了30 d,最低水平时间比CK早60 d。
由图3b可知,大百合种子在层积过程中,CK和2,4-D处理在IAA含量上呈先上升后下降的趋势,不同的层积时间显著极差异(P<0.01)。变温层积0 d时,IAA含量最低,2,4-D处理比CK高出3.98 ng/g;在层积60 d时,IAA含量达到峰值,外源2,4-D抑制内源IAA,CK高达1 954.52 ng/g,比2,4-D处理高出516.66 ng/g;层积120 d时,2,4-D处理的IAA含量上升较快,比 CK高出 173.82 ng/g。IAA含量在层积60 d时,上升速度较快,与30 d相比,CK增长了 1 877.49 ng/g,2,4-D处理增长了 947.2 ng/g,层积90 d时,IAA含量急速下降,与60 d相比,CK降低了1 925.39 ng/g,2,4-D处理下降了1 421.02 ng/g。
由图3c可知,大百合种子在层积过程中,CK和2,4-D处理在GA3含量的水平变化大体上呈先上升后下降的趋势。除层积130 d时差异显著(P<0.05),其他的层积时间显著极差异(P<0.01)。在变温层积0 d时,2,4-D 5 mg/L处理的GA3含量比CK高出9.86 ng/g;GA3含量在层积120 d时达到峰值,2,4-D处理比CK高出52.17 ng/g,外源2,4-D促进内源GA3;层积120 d,GA3含量上升速度较快,与90 d相比,CK增长了38.4 ng/g,2,4-D处理增长了99.72 ng/g;层积130 d时,GA3含量急速下降,与120 d相比,CK降低了40.01 ng/g,2,4-D处理下降了96.34 ng/g。
2.4 大百合种子后熟过程中各激素含量之间的比值变化
由图4可知,大百合种子后熟阶段IAA/ABA、GA3/ABA之间的比值变化也存在差异,在层积过程中,IAA/ABA、GA3/ABA之间的比值整体上呈先上升后下降的趋势。
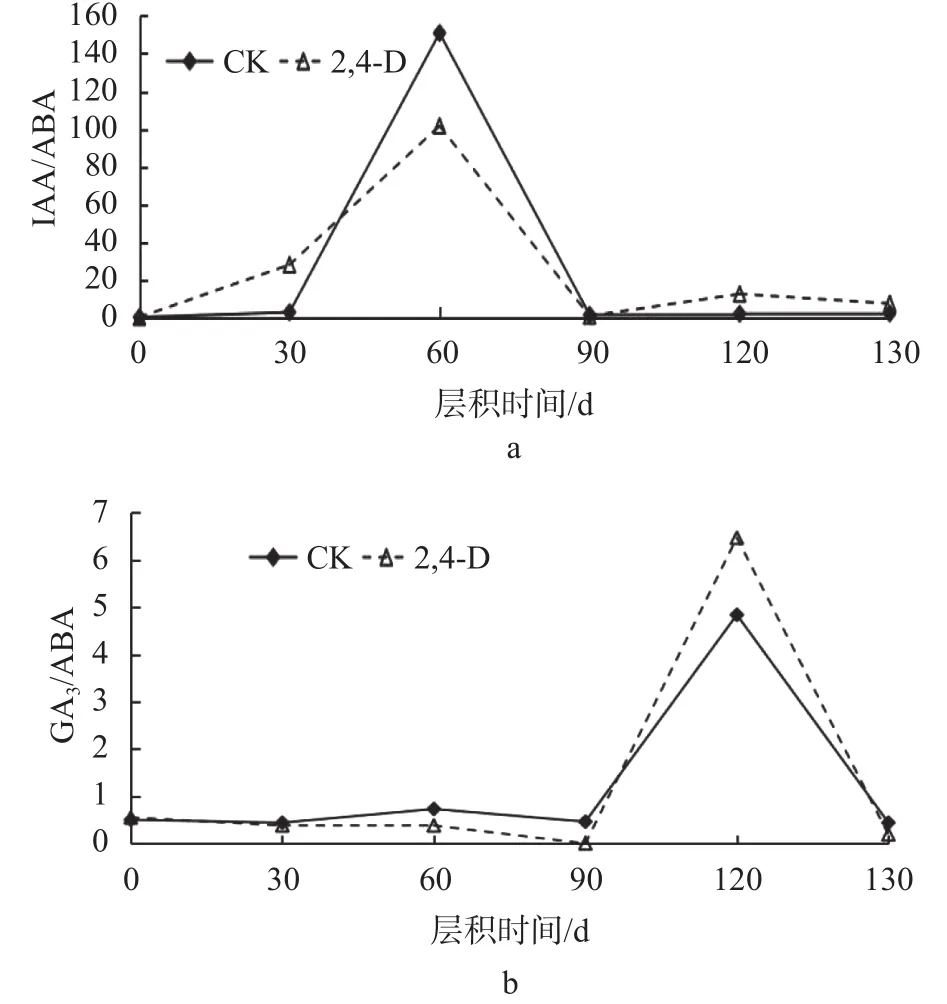
图 4 大百合种子层积过程中各激素含量之间的比值变化Fig. 4 Changes in the ratio of hormones during stratification of C. giganteum seeds
从图4a中可以看出,在层积60 d时,IAA/ABA达到峰值,CK比2,4-D处理高了49.24;IAA/ABA变化较大,在层积60 d时,上升速度较快,与30 d相比,CK的IAA/ABA增长了148.1,2,4-D处理增长了73.66;层积90 d时,IAA/ABA急速下降,与60 d相比,CK降低了149.47,2,4-D处理下降了100.84;层积120 d时,2,4-D处理的IAA/ABA比CK高出10.53。从图4b中可以看出,在层积120 d时,GA3/ABA达到峰值,2,4-D处理比CK高了1.63;GA3/ABA变化较小,在层积120 d时,上升速度较快,与90 d相比,CK的GA3/ABA增长了4.38,2,4-D处理增长了6.47;层积130 d时,GA3/ABA急速下降,与120 d相比,CK降低了4.4,2,4-D处理下降了6.28。IAA/ABA、GA3/ABA上升说明发芽促进物质占相对优势,种子向萌发的方向发展。
3 结论与讨论
种子休眠是植物经过长期的演化而获得的一种对环境条件及季节性变化的生物学适应性[17]。近年来,国际上对形态生理休眠(MPD)种子的研究已取得了一定的进展,但仍被认为是种子生态学领域中了解最少的休眠类型之一。国内对MPD的研究也才刚刚起步,已有的研究仅限于对部分植物的种子形态和萌发处理,研究还不深入、缺乏系统性。姚林君等[18]系统总结了MPD种子的形态结构、休眠打破的环境需求、MPD的起源与进化以及MPD的植物类群。形态生理休眠特性在木兰科植物中非常常见,如西康玉兰(Magnolia wilsonii)[19]。
冯小芹等[20]研究了98%浓硫酸、高温处理、机械处理等6种不同种子处理方法对漆树(Toxicdendro vernicifluum)幼苗生长的影响,本实验采用变温层积结合激素处理对大百合种子休眠解除的影响。张家玲等[21]用2,4-D对滇重楼(Paris polyphylla)种子萌发进行研究,结果表明90 d时,CK的萌发率为40.33%,0.4 mg/L 2,4-D浸种处理48 h滇重楼种子萌发率最高达53.33%,0.2~0.4 mg/L的 2,4-D可促进滇重楼种子的萌发,而高于0.4 mg/L的2,4-D对种子的萌发具有抑制作用。大百合种子的休眠类型属于MPD,本研究采用不同浓度2,4-D处理大百合种子,大百合种子的胚不完全伸长,种子也会萌发,在层积135 d时,5 mg/L 2,4-D处理的胚长比CK高出9.03%,胚的长度不能衡量种子休眠解除的程度。5 mg/L 2,4-D处理的胚生长百分比最高,达到82%,发芽时间最早,为4 ℃下14 d;发芽率最高,达75.33%,5 mg/L 2,4-D处理可以缩短发芽时间,提高种子萌发,利于大百合种胚后熟。1 mg/L 2,4-D处理促进作用不明显,10 mg/L 2,4-D处理起到明显的抑制作用。
刘序等[22]采用GC-MS方法分析鉴定了香榧(Torreya grandis)种子的外种皮、内种皮和胚乳萃取液中内源物种类及其相对含量,本研究采用LC-MS方法测定了大百合种子层积过程中3种内源激素。Khan[23]提出种子的休眠和萌发由3个因子调节,即萌发促进物质GA3、细胞分裂素和ABA之间的相互作用决定种子休眠与萌发。ABA和GA3是调控种子休眠与萌发的一对拮抗内源激素,IAA对萌发没有明显的影响,但间接影响种胚的发育和其他激素水平[24]。大百合种子层积过程中,ABA、IAA和GA3的含量发生不同规律的变化。5 mg/L 2,4-D处理的ABA含量在层积30 d时,急速下降,下降速度的时间比CK早了30 d,在层积60 d时达到最低水平,比CK早60 d。5 mg/L 2,4-D处理的IAA含量和GA3含量在层积的0 d、120 d时,均比CK高。内源激素的相对水平对种子休眠具有重要的调控作用,IAA/ABA在层积60 d达到峰值,CK比5 mg/L 2,4-D高了49.24,GA3/ABA的比值在层积120 d达到峰值,5 mg/L 2,4-D处理比CK高了1.63。
大百合种子的休眠并不是单一因素引起的,而是各种激素共同作用的结果。综合来看,ABA含量的变化与大百合种胚的发育呈负相关,IAA和GA3可能是促进大百合种子休眠解除的主要调节物质,5 mg/L 2,4-D处理通过抑制ABA而促进种胚的萌发。不同外源GA3处理天女木兰(Magnolia sieboldii)[25]胚乳中ABA含量随层积时间大体上呈持续下降趋势,GA3含量均在种子休眠解除期(120 d)含量达到最大,IAA含量呈先升高后下降的趋势,并均在层积40 d时达到峰值,随后逐渐下降,本研究中与天女木兰胚乳中内源激素含量的变化相似,表明适宜浓度的外源GA3和2,4-D浸种处理有利于种子休眠解除和萌发。
[ 参 考 文 献 ]
[1]中国科学院昆明植物研究所. 云南植物志: 第7卷 [M]. 北京: 科学出版社, 1997.
[2]冯国楣. 中国珍稀野生花卉[M]. 北京: 中国林业出版社, 1995.
[3]关文灵, 李世峰, 李叶芳, 等. 大百合鳞茎营养成分分析与评价 [J]. 西部林业科学, 2011, 40(1): 8-11.
[4]万珠珠, 龙春林, 程治英, 等. 重要野生花卉大百合属植物研究进展 [J]. 云南农业大学学报, 2007, 22(1):30-34.
[5]裴盛基, 龙春林. 应用民族植物学[M]. 昆明: 云南民族出版社, 2004.
[6]张金政, 龙雅宜, 孙国峰. 大百合的生物多样性及其引种观察 [J]. 园艺学报, 2002, 29(5): 462-466.
[7]李守丽, 石雷, 张金政, 等. 大百合子房的离体培养 [J]. 园艺学报, 2007, 34(1): 197-200.
[8]关文灵, 李世峰, 雷丽萍, 等. 大百合小孢子发生和雄配子体发育研究 [J]. 亚热带植物科学, 2011, 40(4):12-15.
[9]林凤琼, 朱昌叁, 黎素平, 等. 大百合与百合属优良品种杂交试验 [J]. 南方农业学报, 2012, 43(11): 1733-1736.
[10]Phartyal S S, Kondo T, Baskin C C, et al. Seed dormancy and germination in the giant Himalayan lily(Cardiocrinum giganteum var. giganteum): an assessment of its potential for naturalization in northern Japan [J]. Ecological Research, 2012, 27(4): 677-690.
[11]Cao G X, Worley A C. Life history trade-offs and evidence for hierarchical resource allocation in two monocarpic perennials [J]. Plant Biology, 2013, 15(1):158-165.
[12]Chen H N, Zhao C H, Liu X R, et al. Pollen development of Cardiocrinum giganteum (Wall.) Makina in China [J]. Plant Systematics and Evolution, 2012,298(8): 1557-1565.
[13]关文灵, 李世峰, 陈贤, 等. 大百合种子休眠特性及休眠破除 [J]. 西北植物学报, 2010, 30(12): 2479-2483.
[14]蔡薇, 李叶芳, 刘春雪, 等. 变温层积下激素对大百合种胚后熟的影响 [J]. 中国农学通报, 2017, 33(34):103-110.
[15]刘彩红, 李成云, 朴光一, 等. 2,4-二氯苯氧乙酸对不同羊草种子发芽特性的影响 [J]. 东北农业大学学报,2011, 42(12): 125-129.
[16]Wu J, Hettenhausen C, Meldau S, et al. Herbivory rapidly activates MAPK signaling in attacked and unattacked leaf regions but not between leaves of nicotiana attenuata [J]. The Plant Cell Online, 2007, 19(3):1096-1122.
[17]胡晋. 种子生物学[M]. 北京: 高等教育出版社, 2006.
[18]姚林君, 张克亮, 熊作明, 等. 种子形态生理休眠研究进展 [J]. 生态学杂志, 2019, 38(1): 247-255.
[19]韩春艳, 龙春林. 濒危植物西康玉兰种子休眠、萌发及贮藏特性 [J]. 云南植物研究, 2010, 32(1): 47-52.
[20]冯小芹, 胡勐鸿, 吕寻. 种子不同处理方法对漆树幼苗生长的影响 [J]. 西南林业大学学报(自然科学),2018, 38(6): 48-57.
[21]张家玲, 马英姿, 胡文俐. 不同处理对滇重楼种子萌发的影响 [J]. 经济林研究, 2018, 36(2): 100-106.
[22]刘序, 陈黎, 高捍东. 香榧种子抑制其发芽和生长的内源物筛选研究 [J]. 西南林业大学学报(自然科学),2018, 38(1): 28-33.
[23]Khan A A. Primary, preventive and permissive roles of hormones in plant systems [J]. The Botanical Review,1975, 41(4): 391-420.
[24]赵文琦, 季兰. 种子休眠与解除机理的研究进展 [J].山西农业科学, 2017, 45(3): 477-481.
[25]陆秀君, 梅梅, 刘月洋, 等. GA3和变温层积对天女木兰种子萌发及内源激素的影响 [J]. 西北植物学报,2014, 34(9): 1828-1835.
猜你喜欢
安徽农业科学(2022年18期)2022-10-13
黑龙江科学(2022年6期)2022-04-03
花卉(2020年24期)2021-01-04
防护林科技(2020年11期)2020-12-30
潍坊学院学报(2020年6期)2020-11-22
中国食品学报(2019年3期)2019-01-13
天津农业科学(2018年3期)2018-03-24
江苏农业科学(2017年12期)2017-09-16
浙江农业科学(2016年11期)2016-05-04
安徽理工大学学报·社会科学版(2014年6期)2015-02-06
