
痛记忆模型大鼠ACC脑区蛋白质组学研究及电针干预
2019-09-10 07:22徐立雷孙晶李菲何俏颖何晓芬沈醉刘伯宇方剑乔邵晓梅
世界中医药 2019年6期
徐立雷 孙晶 李菲 何俏颖 何晓芬 沈醉 刘伯宇 方剑乔 邵晓梅
摘要 目的:采用蛋白质组学方法筛选痛记忆模型大鼠ACC脑区差异蛋白,并进一步筛选与电针干预痛记忆可能相关的差异蛋白,为电针干预痛记忆的深入研究提供理论支持。方法:将18只健康雄性SD大鼠随机分为空白组(生理盐水对照组)、模型组和电针组,每组6只。模型组及电针组大鼠通过二次角叉菜胶足跖注射建立痛记忆模型。对照组注射同等剂量的生理盐水。电针组于首次注射后5 h、1~5 d进行电针治疗。模型组与空白组仅作与电针组相同的束缚处理。分别检测3组大鼠造模前,首次注射后4 h、5 d和二次注射后4 h、72 h的机械痛阈。二次注射后第3天,大鼠处死后取ACC脑组织。以双向凝胶电泳分离,双向电泳后经不同波长光激发扫描得到不同样品的蛋白质组图谱,经Image Master2D 6.0软件进行分析后,筛选相差在1.5倍以上的蛋白作为差异表达的蛋白质,并进行质谱鉴定。结果:1)与空白组比较,模型组二次未注射角叉菜胶足跖痛阈显著下降(P<0.05);与模型组比较,电针组二次未注射角叉菜胶足跖痛阈显著上升(P<0.05)。2)经蛋白质组学分析共筛选出18个差异蛋白质。其中模型组有明显差异者有11个,4个表达呈现下调,分别为微管蛋白α-1A链、DAB2相互作用蛋白、NADH脱氢酶的辅酶黄素蛋白2、转凝蛋白-3;7个表达呈现上调,分别为微管蛋白β-3链、肌动蛋白1、磷酸甘油酸激酶1、类固醇激素合成急性蛋白、肌动蛋白2、细胞色素c氧化酶6A1亚基、泛素40S核糖体蛋白S27a。与空白组比较,电针组有明显差异者有15个,3个表达呈现下调,分别为微管蛋白α-1C链、Rho GDP解离抑制因子、NADH脱氢酶的辅酶黄素蛋白2;12个表达呈现上调,分别为微管蛋白β-2A链、微管蛋白β-3链、肌动蛋白1、乌头酸水合酶、丙酮酸激酶同工酶、异柠檬酸脱氢酶、磷酸甘油酸激酶1、Ras相关Rab-19蛋白、类固醇激素合成急性蛋白、肌动蛋白2、细胞色素c氧化酶6A1亚基、泛素40S核糖體蛋白S27a。电针组与模型组比较后呈现明显差异的有8个差异蛋白,2个呈现下调,分别是Rho GDP解离抑制因子、肌动蛋白2。6个呈现上调,分别是微管蛋白α-1A链、微管蛋白β-2A链、DAB2相互作用蛋白、乌头酸水合酶、异柠檬酸脱氢酶、Ras相关Rab-19蛋白。结论:经蛋白质组学分析发现,有11个差异蛋白参与痛记忆的唤醒过程;有8个差异蛋白参与电针干预痛记忆的过程。提示电针对痛记忆的干预可能与稳定神经细胞骨架蛋白,进而抑制神经突触可塑性改变有关。
关键词 电针;痛记忆;前扣带皮层;蛋白质组学;差异蛋白;角叉菜胶;全景式;大鼠
Abstract Objective:To screen the differential protein of ACC brain region in pain memory model rats by proteomics method,to further screen the differential proteins related to electroacupuncture intervention pain memory,so as to provide theoretical support for the further study of electroacupuncture intervention pain memory.Methods:A total of 18 healthy male Sprague Dawley rats were divided into three groups,with 6 rats in each group,including a control group,a model group and an EA group.The pain memory model was made by twice cross-injection carrageenan in EA and model groups.The control group was given the same dose of normal saline in the same time point as the model and EA group.EA was applied to bilateral Zusanli(ST36)and given at 5 h,1~5 days after the first injection for 30 min per time.The model group was taken the same measures except for EA.The pain withdrawal thresholds were detected before modeling and at 4 h,120 h after the first injection and before second injection and at 4 h,72 h after the second injection.Rats were killed at 3rd day after the second injection and taking the ACC of rats for preparation.Samples were separated by 2-D gel electrophoresis(2-DE).Then we got different samples of proteomic map by different excitation wavelength light scan.The gels were respectively image analyzed by Image Master 2D 6.0,to screen the protein changed in abundance more than 1.5 folder up or down compared with which in constitution of the control group as differential proteomics,and then had the mass spectrum identification.Results:1)Compared with the blank group,the pain threshold of carrageenan ankle was significantly decreased in the model group(P<0.05).Compared with the model group,the ankle pain threshold of electroacupuncture group without twice cross-injection carrageenan was rised significantly(P<0.05).)2)After proteomic analysis,18 differential proteins were screened out.Among them,there were 11 with significant difference in the model group,4 of them had down expression,including Tubulin alpha-1C chain,DAB2 interacting protein,NADH dehydrogenase [ubiquinone] flavoprotein 2,Transglutinin-3; and 7 of them had up expression,including Tubulin beta-3,Phosphoglycerate kinase 1,Steroidogenic acute regulatory protein,Actin cytoplasmic 2,Cytochrome c oxidase subunit 6A1,Ubiquitin-40S ribosomal protein S27a.Compared with the blank group,EA group had 15 proteins with significant differences,3 of them had down expression,including Tubulin alpha-1C chain,Rho GDP-dissociation inhibitor 1,NADH dehydrogenase [ubiquinone] flavoprotein 2,and 12 of them had up expression,including Tubulin beta-2A chain,Tubulin beta-3A chain,Actin 1,aconitate hydratase,pyruvate kinase isoenzyme,isocitrate dehydrogenase,phosphoglycerate kinase 1,Ras-related Rab-19 protein,steroid hormone synthesis of acute protein,actin 2,cells Pigment c oxidase 6A1 subunit,ubiquitin 40S ribosomal protein S27a.There were 8 differentially expressed proteins in the EA group compared with the model group,and 2 were down-regulated,which were Rho GDP dissociation inhibitor and actin 2.6 were up-regulated,respectively,tubulin α-1A Chain,tubulin β-2A chain,DAB2 interacting protein,aconitate hydratase,isocitrate dehydrogenase,Ras-related Rab-19 protein.Conclusion:There were 11 differential proteomics involved in pain memory of awakening process through proteomics analysis.And 8 differential proteomics were involved in the intervention of EA on pain memory.The intervention effect of electroacupuncture(EA)pretreatment on pain memory may be related to inhibition of synaptic plasticity.And the inhibition may be related to reinforce nerve cytoskeleton structure.
Key Words Electroacupuncture; Pain memory; Anterior cingulate cortex; Proteomics; Differential proteins; Carrageenan; Panoramic; Rat
中图分类号:R245文献标识码:Adoi:10.3969/j.issn.1673-7202.2019.06.001
痛记忆目前被认为是慢性痛形成的主要原因之一,主要表现为炎性反应、创伤等引起疼痛的初始病因消除后,伤害性感受仍然存在的现象[1-3]。对这类疼痛的治疗目前临床仍以抗炎镇痛药物为主,但临床效果不明显,并且长期恐惧、回避行为将导致患者的生活能力下降、肢体废用、肌力下降等[4],对于疼痛患者的生命质量产生重大影响。因此对痛记忆的干预已逐渐成为疼痛治疗的新热点。
研究发现前扣带皮层(Anterior Cingulate Cortex,ACC)是人类疼痛研究中激活频率最高的脑区[5-7],我们以往研究也已发现电针可有效缓解痛记忆,并且其作用与抑制ACC脑区活性有关[8]。尽管既往研究初步说明电针是干预痛记忆的有效方法,但是目前对于针灸治疗痛记忆的认识极其有限,有下列问题急需解决:1)缺乏蛋白质组学层面对编码痛记忆的关键脑区(ACC)可能涉及的调控网络和细胞通路的证据。尽管有研究报道了与痛记忆变化密切相关的通路或因子包括cAMP/PKA/CERB通路、ERK磷酸化、NMDA受体等[9-11],以及与痛感觉密切相关MAPK信号通路、P2X嘌呤受体、TRPV等[12-14]。但是尚未开展就ACC脑区全景式的蛋白质组学差异蛋白比较研究。2)缺乏蛋白质组学层面针灸抗痛记忆机制认识。我们的前期工作以痛记忆模型研究对象,有力说明了电针对痛记忆早期的干预优势,遗憾的是我们仅关注了p-CREB在痛记忆唤醒过程及电针干预中的作用,没有采用更加合适的蛋白质组学技术全景式揭示电针对痛记忆的干预机制。针灸作为镇痛公认有效的作用多靶点传统疗法,对于揭示针灸镇痛原理的研究多数仍然集中在针灸对特定通路或蛋白的研究,极大限制了本领域的发展。因此,目前的研究成果不能完全阐释针灸对痛记忆干预的原理。本研究基于比较蛋白质组学技术,系统分析痛记忆唤醒阶段ACC脑区差异蛋白表达,并进一步观察电针对差异蛋白表达的影响,这对初步构建针灸干预痛记忆调制网络尤为必要,将为深入了解痛记忆唤醒机制及针灸干预奠定夯实的基础。
1 材料与方法
1.1 材料
1.1.1 动物 选用购自中国科学院上海实验动物中心的18只健康雄性SD大鼠,由浙江中医药大学实验动物中心饲养(实验动物质量合格证号为SYXK(浙)2008-0115),体质量(180±20)g。
1.1.2 药物 角叉菜胶、0.9%生理盐水。
1.1.3 试剂和仪器 Von Frey纤维丝测痛仪(美国stoelting公司),韩式HANS-200A穴位神经刺激仪(联创科技(集团)南京济生医疗科技有限公司),EttanTM DALTSix垂直板电泳仪(GE公司),Typhoon 9400多功能扫描成像系统(GE公司),DeCyder Image QuantTM V6.5凝膠图像析软件(GE公司),基质辅助激光解吸飞行时间串联质谱仪(美国应用生物系统公司),MultiTemp Ⅲ温控循环水浴系统(美国BioSciences公司)。
1.2 方法
1.2.1 分组与模型制备 选用清洁级健康雄性SD大鼠18只,体质量(180±20)g,适应性喂养1周后,将大鼠随机分分为空白组(生理盐水对照组)、模型组、电针组3组,每组6只。
本研究通过2次角叉菜胶注射诱导炎性痛,建立疼痛记忆模型。模型组与电针组大鼠首次造模于左后足跖皮下注射2%角叉菜胶0.1 mL,首次造模后14 d,以同样的方法在2组大鼠右后足跖皮下注射2%角叉菜胶0.1 mL。空白组在相应时间点注射等剂量的生理盐水。
1.2.2 干预方法 参照《实验动物穴位图谱》进行穴位定位,将0.3 mm×13 mm毫针针刺电针组大鼠双侧“后三里”穴,将参考电极置于“后三里”下1 cm处,使用韩氏穴位暨神经刺激仪连接针灸针,电针仪的刺激参数设定如下:波形为疏密波,频率2/100 Hz;波宽0.2 ms;电流强度以动物局部皮肤轻微颤动为度(1 mA~2 mA),起始强度为1 mA,每10 min增加0.5 mA,直至2 mA,共计30 min。电针时间为首次造模后5 h及第1~5天。空白组与模型组大鼠仅作与电针组相同的束缚处理。
1.2.3 指标检测与方法 1)机械缩足阈检测:采用von frey检测大鼠双侧后足缩足阈(Paw Withdrawal Thresholds,PWTs)作为机械痛阈。检测时间段为上午9点至12点,保持室内温度23~25 ℃,湿度45%~55%,噪声40 dB以下。痛阈检测时间点为首次造模前、首次造模后4 h、120 h,2次造模前、2次造模后4 h、72 h。每次测痛连续检测5次,每次间隔5 min,取后面4次缩足阈的平均值作为1个时间点痛阈值。各组大鼠于基础痛阈测量前,进行两天适应性痛阈的检测,以减少环境对实验大鼠痛阈的影响。2)提取总蛋白及电泳:3组大鼠在第2次角叉菜胶注射后的第4天,麻醉状态下经心灌注预冷生理盐水后,取大鼠左侧前扣带皮层20 mg ACC组织,加入200 μL裂解液,匀浆后静置于冰盒30 min。再置于高速低温离心机中设定11 000 r/min,4 ℃离心10 min,结束后提取上清液。并采用BCA法检测蛋白浓度,再进行双向电泳和银染法蛋白质点的检测。3)凝胶银染后图像扫描与软件分析:凝胶电泳之后使用MilliQ纯水冲洗数次,运用Typhoon9400扫描仪进行扫描成像。所获得的蛋白质组图谱采用DeCyderImage QuantTM V6.5图像分析软件进行点识别、点匹配、背景消除以及差异蛋白质分析。确定差异蛋白质后,使用250 μL枪头剪去其前端部分,将感兴趣的蛋白质点对准后,轻压切下,置于离心管中备用。4)胶内酶解:将进行银染后切下的蛋白质点置于离心管中,再用ddH2O将蛋白质凝胶反复冲洗后,依次进行洗胶脱色、分解和剩余物中半胱氨酸烷基化、酶解、萃取、纯化和结合。5)质谱分析(MALDI-TOF-TOF-MS检测):用4800串联飞行时间质谱仪(4800 Plus MALDI TOF/TOFTM Analyzer)进行质谱分析,设定激光强度为3 880,采集数据方式采用反射模式和自动获取数据的模式,每张图谱共采集1 000点,肽段扫描范围为900~4 000 Da,选择信噪比大于5 000的母离子进行二级质谱(MS/MS)分析,每个样品点上选择2个母离子,二级MS/MS激光激发碰撞能量2 KV。每个样品采集3个~5个谱图。质谱数据用Data Explorer(4.0)分析软件去除同位素峰后,用Mascot检索在NCBInr引擎中进行检索。最后,结合2-D图谱上初步的蛋白质的分子量、等电点信息鉴定蛋白质。
2 结果
2.1 电针对痛记忆模型大鼠机械痛阈的影响
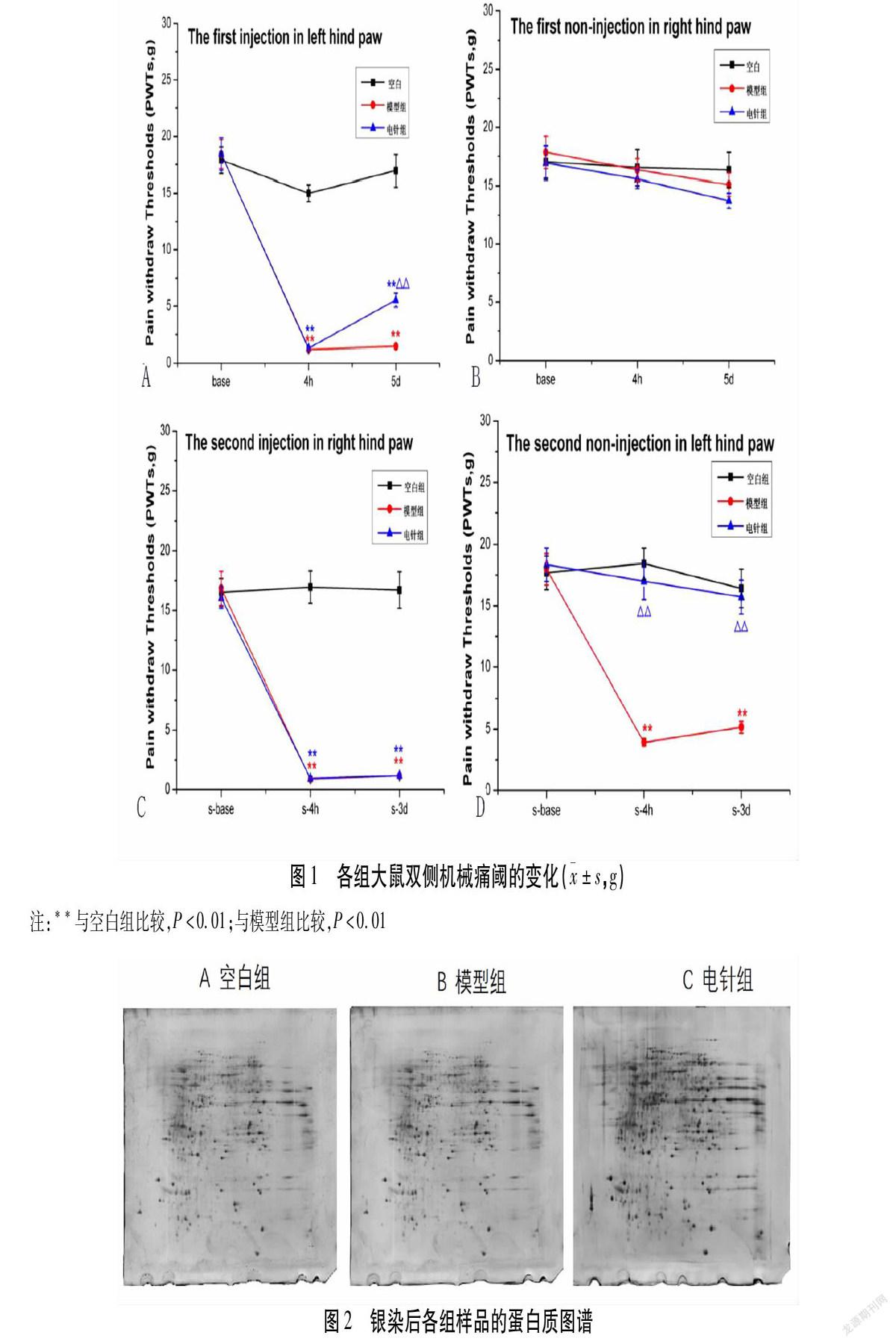
首次造模前,空白组、模型组和电针组大鼠双侧足跖基础痛阈组间比较,差异无统计学意义(P>0.05)。与空白组比较,模型组大鼠左侧足跖首次造模后4 h痛阈显著下降(P<0.01)并延续至造模后120 h(P<0.01)。电针组痛阈在4 h均显著低于空白组(P<0.01)。与模型组比较,电针组造模后4 h痛阈无统计学意义(P>0.05),但在造模后120 h痛阈显著上升(P<0.01)(见图1A)。未造模侧足跖痛阈在各时间点均无统计学意义(P>0.05)(见图1B)。
14 d之后,当大鼠右后足跖痛阈恢复到基础痛阈,进行右后足跖2次角叉菜胶造模(首次未注射侧足跖),2次造模后右后足跖痛阈变化类似于首次造模侧足跖,与空白组比较,模型组与电针组大鼠痛阈在2次造模后4 h至72 h均有显著下降(P<0.01)。模型组与电针组比较,差异无统计学意义(P>0.05)(见图1C)。在大鼠2次未造模侧足跖即首次造模侧足跖,与空白组比较,模型组痛阈在2次造模后4 h、72 h显著下降(P<0.01)(见图1D);电针组痛阈在2次造模后4 h、72 h痛阈无统计学意义(P>0.05)。与模型组比较,电针组2次造模后4 h及72 h痛阈均有显著升高(P<0.01,P<0.01)。
与此同时,空白组各时间点机械痛阈均无统计学意义(P>0.05)。
2.2 电泳图谱分析
图2显示了经过差异蛋白质组分析后的银染蛋白质图谱。可以看出各蛋白质在A空白组、B模型组、C电针组表达量的变化(余蛋白质图谱见附图)。
2.3 质谱分析
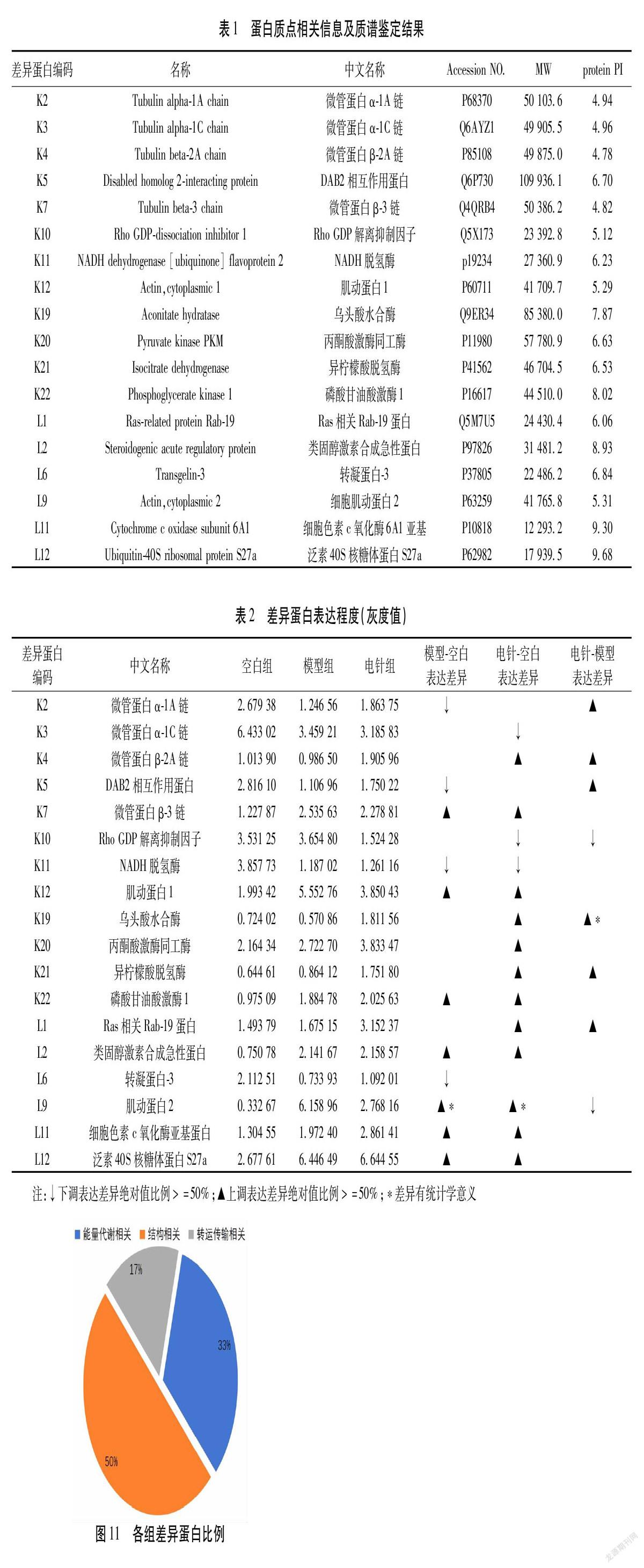
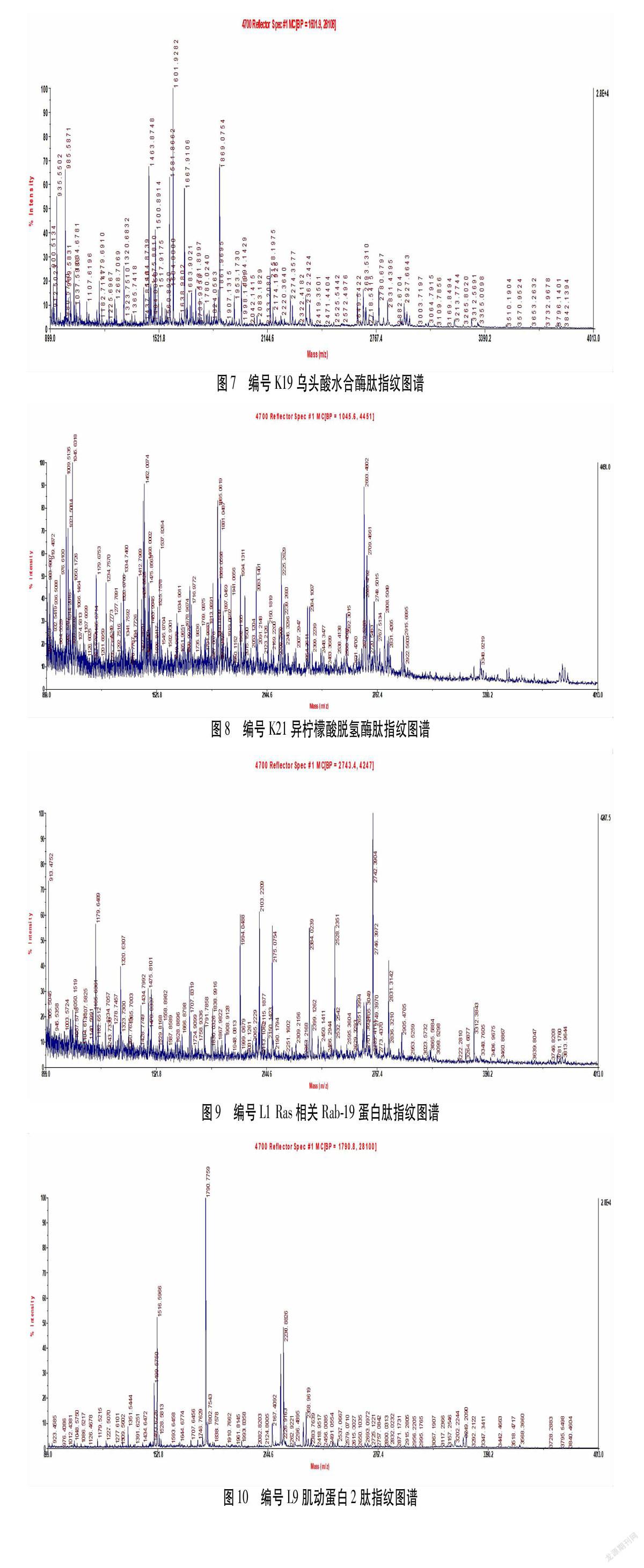
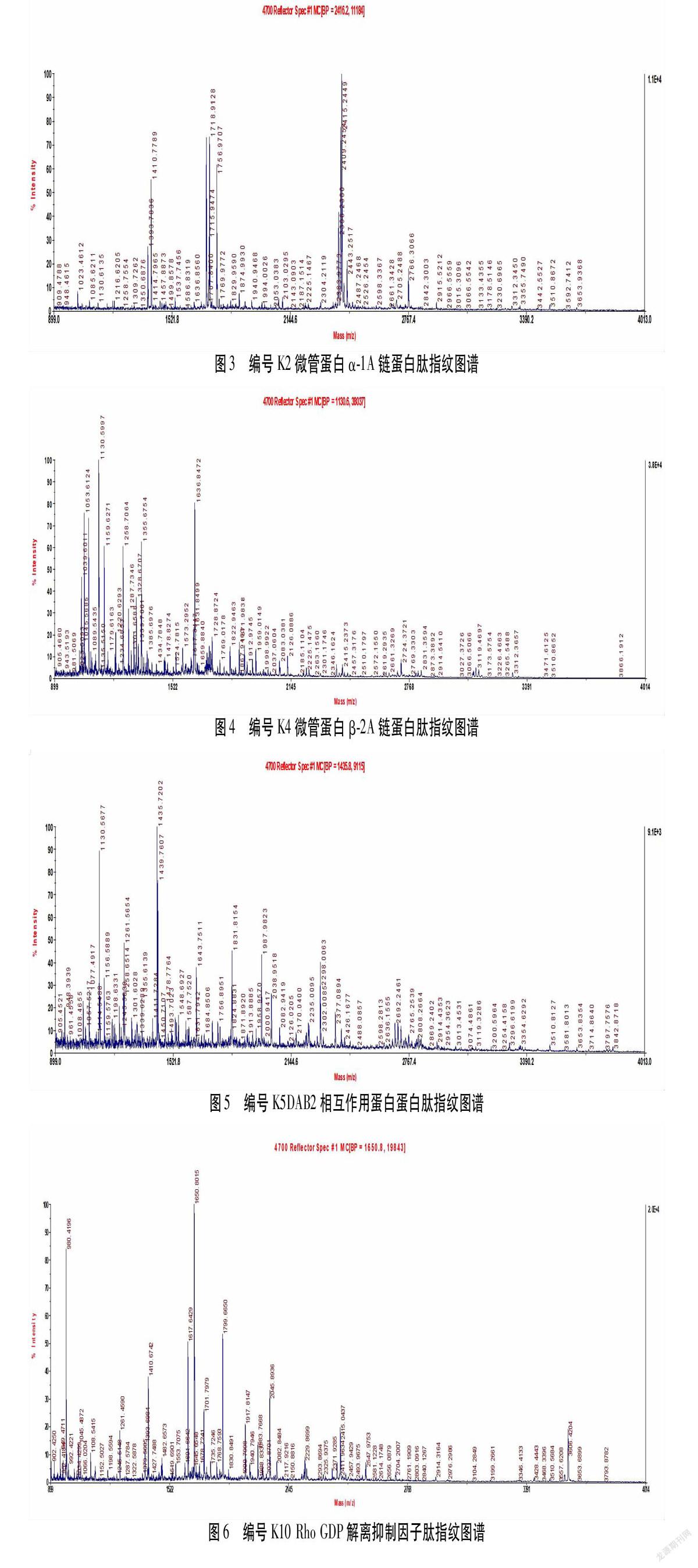
对上述3块胶蛋白质组图谱中相匹配的点进行统计学分析,选择模型组、2/100 Hz电针组与空白组相较表达差值绝对值异超过50%的蛋白质,再将这些蛋白点经胶内酶解后用基质辅助激光电离质谱进行检测,与蛋白质文库进行比对分析后,将蛋白质CI值<60者剔除后,即为本实验所获得差异蛋白质。结果共筛选出18个差异蛋白质,这18种差异蛋白分别是微管蛋白α-1A链(Tubulin alpha-1A chain,Tuba1a)、微管蛋白α-1C链(Tubulin alpha-1C chain,Tuba1c)、微管蛋白β-2A链(Tubulin beta-2A chain,Tubb2a)、DAB2相互作用蛋白(Disabled homolog 2-interacting protein,Dab2ip)、微管蛋白β-3链(Tubulin beta-3 chain,Tubb3)、Rho GDP解离抑制因子(Rho GDP-dissociation inhibitor 1;Arhgdia,rho GDI)、NADH脱氢酶的辅酶黄素蛋白2,(NADH dehydrogenase [ubiquinone] flavoprotein 2,NDUFV2)、肌动蛋白1(Actin,cytoplasmic1,Actb1)、乌头酸水合酶(Aconitate hydratase)、丙酮酸激酶同工酶(Pyruvate kinase,PKM)、异柠檬酸脱氢酶(Isocitrate dehydrogenase)、磷酸甘油酸激酶1(Phosphoglycerate kinase 1,Pgk1)、Ras相关Rab-19蛋白(Ras-related protein Rab-19,Rab19)、类固醇激素合成急性蛋白(Steroidogenic acute regulatory protein,Star)、转凝蛋白-3(Transgelin-3)、肌动蛋白2(Actin,cytoplasmic2,Actb2)、细胞色素c氧化酶6A1亚基(Cytochromec oxidase subunit 6A1,Cox6a1)、泛素40S核糖体蛋白S27 a(Ubiquitin-40S ribosomal protein S27a,Rps27a)等18个差异蛋白。其相关性质见表1。图3~图10为K2、K4、K5、K10、K19、K21、L1、L9,等8个差异蛋白的肽指纹图谱。
经质谱鉴定后各蛋白质在3组间表达程度(灰度值)及差异如表2(差异绝对值比例>50%为有统计学意义),表述如下:微管蛋白α-1A链,在空白组表达值为2.679 38,模型组1.246 56,电针组1.863 75;与空白组比较,模型组明显下调,电针组无统计学意义,电针组与模型组比较明显上调;微管蛋白α-1C,在空白组表达值为6.433 03,模型组3.459 21,电针组3.185 83;与空白组比较,模型组无统计学意义,电针组明显下调,电针组与模型组比较,差异无统计学意义;链微管蛋白β-2A链,在空白组表达值为1.013 90,模型组0.986 50,电针组1.905 96;与空白组比较,模型组无统计学意义,电针组明显上调,模型组与电针组比较明显上调;DAB2相互作用蛋白,在空白组表达值为2.816 10,模型组1.106 96,電针组1.750 22;与空白组比较,模型组明显下调,电针组无统计学意义,电针组与模型组比较明显上调;微管蛋白β-3链,在空白组表达值为1.227 87,模型组2.535 63,电针组2.278 81;与空白组比较,模型组明显上调,电针组明显上调,电针组与模型组比较,差异无统计学意义;Rho GDP解离抑制因子,在空白组表达值为3.531 25,模型组3.654 80,电针组1.524 28;与空白组比较,模型组无统计学意义,电针组明显下调,电针组与模型组比较明显下调;NADH脱氢酶,在空白组表达值为3.857 73,模型组1.187 02,电针组1.261 16;与空白组比较,模型组明显下调,电针组明显下调,电针组与模型组比较,差异无统计学意义;肌动蛋白1,在空白组表达值为1.993 42,模型组5.552 76,电针组3.850 43;与空白组比较,模型组明显上调,电针组明显上调,电针组与模型组比较,差异无统计学意义;乌头酸水合酶,在空白组表达值为0.724 02,模型组0.570 86,电针组1.811 56;与空白组比较,模型组无统计学意义,电针组明显上调,电针组与模型组比较明显上调;丙酮酸激酶同工酶,在空白组表达值为2.164 34,模型组2.722 70,电针组3.833 47;与空白组比较,模型组无统计学意义,电针组明显上调,电针组与模型组比较,差异无统计学意义;异柠檬酸脱氢酶,在空白组表达值为0.644 61,模型组0.864 12,电针组1.751 80;与空白组比较,模型组无统计学意义,电针组明显上调,电针组与模型组比较明显上调;磷酸甘油酸激酶1,在空白组表达值为0.975 09,模型组1.884 78,电针组2.025 63;与空白组比较,模型组明显上调,电针组明显上调,电针组与模型组比较,差异无统计学意义;Ras相关Rab-19蛋白,在空白组表达值为1.493 79,模型组1.675 15,电针组3.152 37;与空白组比较,模型组无统计学意义,电针组明显上调,电针组与模型组比较明显上调;类固醇激素合成急性蛋白,在空白组表达值为0.750 78,模型组2.141 67,电针组2.158 57;与空白组比较,模型组明显上调,电针组明显上调,电针组与模型组比较,差异无统计学意义;转凝蛋白-3,在空白组表达值为2.112 51,模型组0.733 93,电针组1.092 01;与空白组比较,模型组明显下调,电针组无统计学意义,电针组与模型组比较,差异无统计学意义;肌动蛋白2,在空白组表达值为0.332 67,模型组6.158 96,电针组2.768 16;与空白组比较,模型组明显上调,电针组明显上调,电针组与模型组比较明显下调;细胞色素c氧化酶6A1亚基,在空白组表达值为1.304 55,模型组1.972 40,电针组2.861 41;与空白组比较,模型组明显上调,电针组明显上调,电针组与模型组比较,差异无统计学意义;泛素40S核糖体蛋白S27a,在空白组表达值为2.677 61,模型组6.446 49,电针组6.644 55;与空白组比较,模型组明显上调,电针组明显上调,电针组与模型组比较,差异无统计学意义。
3 讨论
疼痛是一种跟机体组织受到伤害或者跟潜在性伤害有关的不愉快的主观感觉和情绪体验(IASP 1994)。疼痛的定义中包含了疼痛感觉和疼痛情绪等两方面。Apkarian等[15]认为记忆是人们不由自主的行为,疼痛的发生会与学习和记忆相关联。随着疼痛的发生,记忆就可以被唤醒而恢复。本次实验我们所使用的痛记忆模型是参照Igor的方法[16]建立的,在前人的研究结果中已经确认角叉菜胶2次交叉注射大鼠足跖方法,将其作为疼痛记忆的评价模型[17-21]。前期研究工作中,本研究小组发现2/100频率早期电针对痛记忆有较好的干预作用。
本研究经质谱仪共筛选出18个差异蛋白依其功能大致可以分为3类,即结构相关蛋白、传导转运相关蛋白、能量代谢相关蛋白。依其数量结构相关蛋白占总数的50%,传导转运相关蛋白占总数的17%、能量代谢相关蛋白占总数的33%。各组差异蛋白所占比例如图11所示。
将空白组、模型组与电针组比较,表达差异值绝对值超过50%,并符合质谱鉴定后蛋白质CI值>60者,为最终被确定的差异蛋白质,共获得18种差异蛋白,分别是微管蛋白α-1A链、微管蛋白α-1C链、微管蛋白β-2A链、DAB2相互作用蛋白、微管蛋白β-3链、Rho GDP解离抑制因子、NADH脱氢酶2,、肌动蛋白1、乌头酸水合酶、丙酮酸激酶同工酶、异柠檬酸脱氢酶、磷酸甘油酸激酶1、Ras相关Rab-19蛋白、类固醇激素合成急性蛋白、转凝蛋白-3、肌动蛋白2、细胞色素c氧化酶6A1亚基、泛素40S核糖体蛋白S27a。
痛记忆产生后(模型组)有11個差异蛋白表达比较有统计学意义。分别是结构相关:转凝蛋白-3、微管蛋白α-1A链、DAB2相互作用蛋白、微管蛋白β-3链、肌动蛋白1、肌动蛋白2;转运传输相关:类固醇激素合成急性蛋白、泛素40S核糖体蛋白S27a;能量代谢相关:NADH脱氢酶、磷酸甘油酸激酶1、细胞色素c氧化酶6A1亚基蛋白等。此11个差异表达蛋白质的组合性变化与痛记忆唤醒有关。
电针干预后有15个差异蛋白表达比较有统计学意义。分别为微管蛋白α-1C链、Rho GDP解离抑制因子、NADH脱氢酶2、微管蛋白β-2A链、微管蛋白β-3链、肌动蛋白1、乌头酸水合酶、丙酮酸激酶同工酶、异柠檬酸脱氢酶、磷酸甘油酸激酶1、Ras相关Rab-19蛋白、类固醇激素合成急性蛋白、肌动蛋白2、细胞色素c氧化酶6A1亚基、泛素40S核糖体蛋白S27a。此15个差异表达蛋白质的组合性变化与电针干预有关。
电针组与模型组比较后有8个差异蛋白表达比较有统计学意义。微管蛋白α-1A链、DAB2相互作用蛋白、Rho GDP解离抑制因子、微管蛋白β-2A链、肌动蛋白2、Ras相关Rab-19蛋白、乌头酸水合酶、异柠檬酸脱氢酶等。此8个差异表达蛋白质的组合性变化与电针干预痛记忆有关。
模型组的11个差异蛋白,与电针组的15个差异蛋白中,有8个差异蛋白相同,而此8个差异蛋白经由模型组与电针组比较后,只有肌动蛋白2一个蛋白质比较有统计学意义。且差异表达趋势与动物实验结果相同;提示肌动蛋白2在痛记忆唤醒过程及及电针干预痛记忆中扮演主要角色。
电针组与模型组比较后,电针组中有9个差异蛋白表达无统计学意义。分别是微管蛋白α-1C链、NADH脱氢酶2、微管蛋白β-3链、肌动蛋白1、丙酮酸激酶同工酶、磷酸甘油酸激酶1、类固醇激素合成急性蛋白、细胞色素c氧化酶6A1亚基、泛素40S核糖体蛋白S27a。此9个差异表达蛋白质的组合性变化其表达为电针干预后反应,与电针干预痛记忆无关。
综上所述,本研究发现11个差异蛋白参与痛记忆的唤醒过程。分别是结构相关:转凝蛋白-3、微管蛋白α-1A链、DAB2相互作用蛋白、微管蛋白β-3链、肌动蛋白1、肌动蛋白2;转运传输相关:类固醇激素合成急性蛋白、泛素40S核糖体蛋白S27a;能量代谢相关:NADH脱氢酶、磷酸甘油酸激酶1、细胞色素c氧化酶6A1亚基蛋白等;本研究发现8个差异蛋白参与电针干预痛记忆的过程。分别是结构相关:微管蛋白α-1A链、DAB2相互作用蛋白、Rho GDP解离抑制因子、微管蛋白β-2A链、肌动蛋白2;转运传输相关:Ras相关Rab-19蛋白;能量代谢相关:乌头酸水合酶、异柠檬酸脱氢酶等;电针对痛记忆的干预可能与稳定神经细胞骨架蛋白,进而抑制神经突触可塑性改变有关,但其作用的具体机制还需要进一步研究。
参考文献
[1]Choi D S,Choi D Y,Whittington R A,et al.Sudden amnesia resulting in pain relief:the relationship between memory and pain[J].Pain,2007,132(1):206-210.
[2]Terry R,Brodie E E,Niven C A.Exploring the Phenomenology of Memory for Pain:Is Previously Experienced Acute Pain Consciously Remembered or Simply Known?[J].Journal of Pain,2007,8(6):467-475.
[3]韩济生,疼痛学[M].北京:北京大学医学出版社,2012.
[4]Hylands-White N,Duarte R V,Raphael J H.An overview of treatment approaches for chronic pain management[J].Rheumatology International,2016,37(1):1-14.
[5]Chen T,Taniguchi W,Chen Q Y,et al.Top-down descending facilitation of spinal sensory excitatory transmission from the anterior cingulate cortex[J].Nature Communications,2018,5:235.
[6]Bliss T V P,Collingridge G L,Kaang B K,et al.Synaptic plasticity in the anterior cingulate cortex in acute and chronic pain[J].Nature Reviews Neuroscience,2016,10:350.
[7]Barthas F,Sellmeijer J,Hugel S,et al.The Anterior Cingulate Cortex Is a Critical Hub for Pain-Induced Depression[J].Biological Psychiatry,2014,77(3):786.
[8]Xiao-Mei S,Jing S,Yong-Liang J,et al.Inhibition of the cAMP/PKA/CREB Pathway Contributes to the Analgesic Effects of Electroacupuncture in the Anterior Cingulate Cortex in a Rat Pain Memory Model[J].Neural Plasticity,2016,2016:1-16.
[9]Cao H,Ren W H,Zhu M Y,et al.Activation of glycine site and GluN2B subunit of NMDA receptors is necessary for ERK/CREB signaling cascade in rostral anterior cingulate cortex in rats:Implications for affective pain[J].Neuroscience Bulletin,2012,28(1):77-87.
[10]Zheng W.Activation of mu opioid receptor inhibits the excitatory glutamatergic transmission in the anterior cingulate cortex of the rats with peripheral inflammation[J].European Journal of Pharmacology,2010,628(1-3):91-95.
[11]Cao H,Gao Y J,Ren W H,et al.Activation of Extracellular Signal-Regulated Kinase in the Anterior Cingulate Cortex Contributes to the Induction and Expression of Affective Pain[J].Journal of Neuroscience,2009,29(10):3307-3321.
[12]Pankratov Y,Lalo U,Krishtal O,et al.P2X receptor-mediated excitatory synaptic currents in somatosensory cortex[J].Molecular and Cellular Neuroscience,2003,24(3):0-849.
[13]Fang J Q,Fang J F,Yi Liang,et al.Electroacupuncture mediates extracellular signal-regulated kinase 1/2 pathways in the spinal cord of rats with inflammatory pain[J].BMC Complementary and Alternative Medicine,2014,14(1):541.
[14]Malek N,Pajak A,Kolosowska N,et al.The importance of TRPV1-sensitisation factors for the development of neuropathic pain[J].Molecular and Cellular Neuroscience,2015,65:1-10.
[15]Apkarian A V.Pain perception in relation to emotional learning[J].Current Opinion in Neurobiology,2008,18(4):464-468.
[16]Kissin I,Freitas C F,Jr B E.Memory of pain:the effect of perineural resiniferatoxin[J].Anesthesia & Analgesia,2006,103(3):721-728.
[17]Coderre T J,Melzack R.Increased pain sensitivity following heat injury involves a central mechanism[J].Behavioural Brain Research,1985,15(3):259-262.
[18]Levine J.Reflex neurogenic inflammation.I.Contribution of the peripheral nervous system to spatially remote inflammatory responses that follow injury[J].Neuroscience,1985,5:234.
[19]Kayser V,Guilbaud G.Local and remote modifications of nociceptive sensitivity during carrageenin-induced inflammation in the rat[J].Pain,1987,28(1):99-107.
[20]Rivat C,Laulin J P,Corcuff J B,et al.Fentanyl Enhancement of Carrageenan-induced Long-lasting Hyperalgesia in Rats[J].Anesthesiology,2002,96(2):381-391.
[21]Fletcher D,Kayser V,Guilbaud G.The influence of the timing of bupivacaine infiltration on the time course of inflammation induced by two carrageenin injections seven days apart[J].Pain,1997,69(3):303-309.
(2019-06-04收稿 責任编辑:徐颖)
猜你喜欢
中国药学药品知识仓库(2022年10期)2022-05-29
中国典型病例大全(2021年13期)2021-12-08
广东教育·职教版(2021年3期)2021-04-20
爱你·健康读本(2020年4期)2020-04-30
中国中医药信息杂志(2019年6期)2019-06-30
分析化学(2019年3期)2019-03-30
学校教育研究(2018年27期)2018-05-14
食品安全导刊(2014年3期)2014-12-02
中国针灸(2000年5期)2000-06-13
