
食源性单增李斯特菌毒力岛基因检测与致病性
2019-09-23 06:10李红欢陈朔康立超
江苏农业科学 2019年14期
李红欢 陈朔 康立超
摘要:为了解食源性单增李斯特菌分离株的毒力岛1(LIPI-1)和毒力岛2(LIPI-2)的基因及致病性。利用PCR方法对8株分离株LIPI-1和LIPI-2进行基因检测,小鼠腹腔接种分离株菌悬液5×107 CFU/只,进行致病性的初步试验。结果显示,LIPI-1的6个毒力基因除actA基因检出率为50.0%外,hly、prfA、plcA、plcB、mpl的检出率为100.0%;LIPI-2中inlA、inlB、inlC基因检出率为100.0%,inlD和inlE基因检出率分别为87.5%和75.0%,inlF和inlG毒力因子基因的检出率较低,分别为50.0%和37.5%。小鼠致病性试验显示,8株分离株均能致死小鼠,致死率在50.0%~100.0%。其中,LM5567和LM5570在5 d内全部死亡,致死率为100.0%,表明这2株菌对小鼠有强致病力。说明LIPI-1基因检出率高于LIPI-2的基因检出率;菌株的致病性与毒力岛基因的携带率无明显相关,但分离株对小鼠均有致病性,提示食品安全不容忽视。本研究为食源性单增李斯特菌毒力岛基因携带率与致病力的关系提供铺垫。
关键词:食源性单增李斯特菌;毒力岛1(LIPI-1);毒力岛2(LIPI-2);致病性;基因检出率
中图分类号: S852.61 文献标志码: A 文章编号:1002-1302(2019)14-0193-04
李斯特菌属包括单增李斯特菌(Listeria monocytogenes,LM或L. monocytogenes)、绵羊李斯特菌(L. ivanuii)、英诺克李斯特菌(L. ivanovii)、罗氏李斯特菌(L. rocourtiae)、默氏李斯特菌(L. marthii)、西尔李斯特菌(L. seeligeri)、格氏李斯特菌(L. grayi)和威尔斯李斯特菌(L. welshimeri)8种菌[1],其中,LM是食源性李斯特菌病的条件性致病菌,与人的疾病密切相关,可穿越肠道屏障、血脑屏障和胎盘屏障[2],在临床上引起脑炎、脑膜炎、流产和死胎等,同时该菌可引起家禽坏死性肝炎和心肌炎的病理变化[3]。对于人类,特别是新生儿、孕妇、老年人及免疫功能缺陷者易引起疾病,致死率达 20%~30%[4-5],在新生幼儿和免疫力低下的人群中更高达70%[6]。许多国家已将此菌列为食品微生物安全检测项目,并对进口食品提出检验该菌的严格要求[7]。
LM是革兰氏阳性、无芽胞兼性厌氧胞内寄生菌,广泛存在于自然界中,有较强的环境适应性,可在低温、高盐、酸碱等不利的环境条件下生长繁殖[8],通过多种途径进入食品及食品加工环境。LM是侵袭性胞内菌,主要通过污染的食物经口进入到宿主的胃肠道内,在细菌的黏附侵袭等相关毒力因子的作用下进入到宿主细胞内,通过血液和淋巴循环系统到达各组织器官,最终引起全身感染[9]。其感染过程包括内化、逃离吞噬泡、宿主细胞内的极向运动和细胞内的传播,整个感染过程都有相应的毒力因子参与,这些毒力因子与LM致病性有关,参与其主要的致病过程,编码这些毒力因子的L毒力基因常聚类于毒力岛。LM主要有毒力岛1(LIPI-1)和毒力岛2(LIPI-2)2 个毒力岛,毒力岛1(LIPI-1)与单增李斯特菌的胞内感染相关[10],主要由prfA、plcA、hly、mpl、actA和plcB 6个毒力基因组成,毒力岛2(LIPI-2)与单增李斯特菌的黏附、侵袭有关,由inlA、inlB、inlC等多个内化素组成。
LM的毒力因子與其致病性有密切联系,研究表明,当LM缺少某些重要的毒力因子时,LM的毒力将大大降低[11]。因此,研究食源性LM毒力因子携带率与致病力的关系对于人类李斯特菌病的监控、暴发流行的监测以及追踪污染源有重大意义。
1 材料与方法
1.1 菌株与试验动物
本试验所用食品源单增李斯特菌,由新疆农垦科学院食品检测中心分离鉴定,石河子大学动物科技学院预防兽医学实验室保存(表1)。48只6~8周昆明小鼠购自石河子大学动物实验中心。
1.2 培养基和主要试剂
脑心浸液培养基(BHI),购自青岛高科技园海博生物技术有限公司;2×PCR Mix、超纯水,均购于北京东盛生物公司;DNA Marker(2000),购于北京东盛生物公司;琼脂糖,购于Biowest公司;琼脂,购于Biotopped公司。
1.3 引物设计与合成
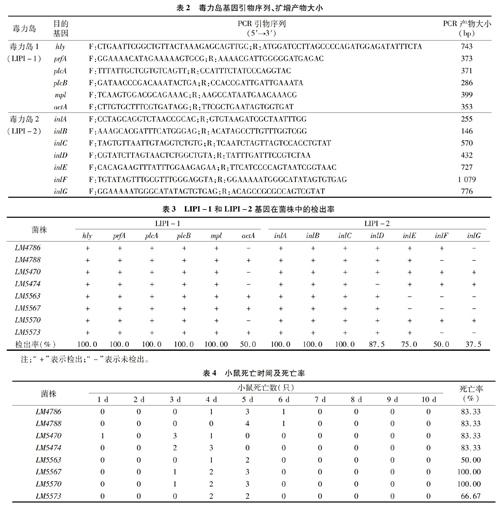
参考GenBank中登录的LMF2365基因全长序列(AE017262),用Primer 5.0软件设计毒力因子基因引物(表2),毒力岛1部分基因引物设计参考文献[12],引物由华大基因公司合成。
1.4 菌株培养
将-80 ℃保存的8株菌种取出,于BHI固体培养基上划线,并于37 ℃培养20 h,挑取单个菌落接种于BHI液体培养基中,置于37 ℃、180 r/min的恒温摇床中培养16 ~18 h,置于4 ℃备用。
1.5 分离株毒力岛基因PCR扩增及检测
PCR扩增体系:10 μL 2×PCR mix,6 μL超纯水,2 μL菌液,上游引物(25 mmol/L),下游引物(25 mmol/L)各1 μL,总体积20 μL。
PCR扩增条件:(1)hly:95 ℃ 5 min;94 ℃ 40 s,60 ℃ 30 s,72 ℃ 1 min,35个循环;72 ℃ 10 min。(2)prfA、plcA、plcB、actA、mpl:94 ℃ 5 min;94 ℃ 50 s,55 ℃ 50 s,72 ℃ 50 s,30个循环;72 ℃ 10 min。(3)inlA、inlB、inlC、inlD、inlE、inlF、inlG:95 ℃ 5 min;94 ℃ 60 s,55 ℃ 60 s,72 ℃ 60 s,35个循环;72 ℃ 10 min。
取20 μL扩增产物于1.5%琼脂糖凝胶电泳检测,与标准DS2000 DNA Marker比较。
1.6 分离株对小鼠的致病性试验
将8株LM接种于BHI中,于37 ℃培养24 h,12 000 r/min离心2 min,弃上清液,用无菌PBS洗涤2次,制备成浓度为108 CFU/mL的菌悬液,取此菌悬液经小鼠腹腔注射 0.5 mL/只,每组6只,连续10 d观察小鼠死亡情况。
2 结果与分析
2.1 分离株毒力岛基因PCR扩增结果
用LIPI-1相应的引物分别对hly、prfA、plcA、plcB、actA、mpl基因进行PCR扩增,PCR产物经过1.5%琼脂糖凝胶电泳,与DS2000相比,片段长度分别约为743、373、371、286、286、399 bp,均符合预期(图略),表明分离株中含有相应的毒力岛基因。
用LIPI-2相应的引物分别对inlA、inlB、inlC、inlD、inlE、inlF、inlG基因进行PCR扩增,PCR产物经过1.5%琼脂糖凝胶电泳,与DS2000相比,片段长度分别大约为255、146、570、432、727、1079、776 bp,均符合预期(图略),表明分离株中含有相应的毒力岛基因。
2.2 分离株毒力岛基因的检测结果
采用PCR方法检测分离株的毒力岛基因,LIPI-1和LIPI-2基因在菌株中的检出率(表3)显示,不同毒力基因的检出率不同。LIPI-1有5个毒力基因的检出率为 100.0%,分别为hly、prfA、plcA、plcB和mpl;actA基因的检出率为50.0%。LIPI-2有3个毒力基因的检出率为100%,分别为inlA、inlB和inlC;inlD基因的检出率为87.5%;inlE基因的检出率为75.0%;inlF和inlG 2个毒力基因检出率相对较低,仅为50.0%和37.5%。由此可见,8株分离株中LIPI-1毒力岛基因检出率高于LIPI-2。
8株食源性LM分离株中,LM5570和LM5470仅存在actA毒力基因的缺失,毒力岛基因的携带率最高,为92.31%(12/13),LM5563和LM5567缺失inlE、inlF、inlG基因,毒力岛基因的携带率最低,为76.92%(10/13),其余LM分离株均缺失2个毒力基因。
2.3 不同分离株对小鼠致病性试验
采用对小鼠腹腔注射细菌悬液5×107 CFU/只,感染小鼠12 h均表现精神沉郁、行动缓慢、食欲低下,接种的小鼠死亡时间集中在3~5 d,6 d后不再有小鼠死亡。LM5567、LM5570菌株接种小鼠在5 d内全部死亡,死亡率为100.00%;LM4786、LM4788、LM5470和LM5474菌株接种小鼠死亡率为83.33%,其中LM5470和LM5474菌株接种小鼠,4 d内死亡5只小鼠,LM4786和LM4788菌株接种小鼠,6 d内死亡5只小鼠。LM5573菌株接种小鼠,5 d内死亡4只,死亡率为 66.67%。LM5563菌株接种小鼠,5 d内死亡3只小鼠,死亡率为50.00%。LM5570和LM5470毒力岛基因的携带率最高(12/13),死亡率为100.00%和83.33%;LM5567和LM5563毒力岛基因的携带率最低(10/13),死亡率为100.00%和 50.00%;LM5474、LM4786、LM4788和LM5573基因携带率为84.62%(11/13),死亡率分别为83.33%、83.33%、83.33%和66.67%(表4)。表明8株分离株均能致死小鼠,但不同分离株致死率不同,且与基因携带率无明显相关。
2.4 各菌株毒力岛基因与小鼠致病力的关系
含有相同毒力基因的LM菌株所表现的致病力并不相同,LM5470和LM5570所含毒力因子相同,均缺少actA毒力因子,但小鼠死亡時间和死亡率并不同。感染LM5470的小鼠,1 d后开始死亡,4 d内死亡5只,死亡率为 83.33%;感染LM5570的小鼠,3 d后开始死亡,5 d内6只实验小鼠全部死亡,死亡率为100%。结果表明,LM5570致病力较强于LM5470菌株。
LM4788和LM5573所含毒力因子相同,均缺少inlF和inlG毒力因子,小鼠死亡时间和死亡率并不同;感染LM4788的小鼠,5 d后开始死亡,6 d内死亡5只,死亡率为83.33%;感染LM5573的小鼠在4 d后开始死亡,5 d内死亡4只,死亡率为66.67%。结果表明,LM5567致病力较LM4788强。
LM5563和LM5567所含毒力因子相同,均缺少inlE、inlF和inlG毒力因子,但小鼠死亡时间和死亡率并不同。感染M5563的小鼠,5 d内死亡3只小鼠,死亡率为50.00%;感染LM5567的6只小鼠在5 d内全部死亡,死亡率为100.00%。结果表明,LM5567的致病力较LM5563强。
上述结果表明,含有相同的毒力岛基因的分离株对小鼠的致死率并不相同,菌株基因的携带率与其致病性无明显相关,表明对LIPI-1和LIPI-2的基因检测不足反映其与致病力的关系。
3 讨论
LM一般经胃肠道感染,侵入肠上皮细胞后被单核巨噬细胞吞噬,并随着扩散到局部淋巴结,最后到达内脏器官,引起全身性感染[12]。李斯特菌整个感染过程中的每一步均有特定的毒力因子调控。LM的致病性与其毒力基因密切相关,缺失毒力基因将导致其致病性的消失或者下降。通过检测毒力基因,可以掌握这些毒力基因在LM中的分布,从而为评估LM分离株的致病性强弱提供理论基础。
本研究通过PCR技术检测了8株不同来源食源性LM分离株LIPI-1和LIPI-2的13个毒力基因,有研究表明 LIPI-1 毒力岛与LM的致病性密切相关,是细菌在细胞内生存必不可少的,具有高度的保守性[13]。本试验中LIPI-1基因除actA之外,其余基因均未缺失,检出率为100.0%,具有较高的稳定性,LIPI-1基因的检出率高于伊娜娜等的报道[14-15],说明本区域的食源性LM分离株LIPI-1的基因的检出率较高。LIPI-2与LM的黏附、侵袭有关,inlA、inlB和inlC的检出率达100.0%,inlD、inlE、inlF和inlG的检出率分别是87.5%、75.0%、50.0%和37.5%,LIPI-1基因检出率高于LIPI-2基因,本试验结果与刘二龙等的报道[16-18]一致。
本研究中将不同分离株对小鼠进行致病性试验表明,8株LM分离株均能致死小鼠,但不同分离株致死率不同,其致死率与所检测毒力岛基因携带率之间无明显相关,提示目前所检测的菌株数量或毒力岛基因不足以反映菌株的致病力,仍需进一步研究,但食品污染LM的安全不容忽视。
参考文献:
[1]Hain T,Chatterjee S S,Ghaia R,et al. Pathogenomics of Listeria spp.[J]. International Journal of Medical Microbiology,2007,297(7/8):541-557.
[2]Swaminathan B,Gerner-Smidt P. The epidemiology of human listeriosis[J]. Microbes and Infection,2007,9(10):1236-1243.
[3]Ramaswamy V,Cresence V J,Lekshmi M,et al. Listeria—Review of epidemiology and pathogenesis[J]. Journal of Microbiology,Immunology,and Infection,2007,40(1):4-13.
[4]Fonnesbech V B,Huss H H,Ojeniyi B,et al. Elucidation of Listeria monocytogenes contamination routes in cold-smoked salmon processing plants detected by DNA-based typing methods[J]. Applied & Environmental Microbiology,2001,67(6):2586-2595.
[5]Vlaemynck G,Lafarge V,Scotter S. Improvement of the detection of Listeria monocytogenes by the application of ALOA,adiagnostic,chromogenic isolation medium[J]. Journal of Applied Microbiology,2000,88(3):430-441.
[6]Klinger G,Beyene J,Shah P,et al. Do hyperoxaemia and hypocapnia add to the risk of brain injury after intrapartum asphyxia?[J]. Archives of Disease in Childhood-Fetal and Neonatal Edition,2005,90(1):49-52.
[7]张淑红,吴清平,张菊梅. 显色培养基在单核细胞增生李斯特菌快速检测中的应用研究[J]. 中国卫生检验杂志,2007,17(1):43-45.
[8]陈云飞. 单增李斯特菌ΔInlAB、ΔInlABC突变株的构建及部分生物学特性研究[D]. 石河子:石河子大学,2015.
[9]Dussurget O,Pizarrocerda J,Cossart P. Molecular determinants of Listeria monocytogenes virulence[J]. Annual Review of Microbiology,2004,58(1):587-610.
[10]Glaser P,Frangeul L,Buchrieser C,et al. Comparative genomics of Listeria species[J]. Science,2001,294(5543):849-852.
[11]Freitag N E,Port G C,Miner M D. Listeria monocytogenes — From saprophyte to intracellular pathogen[J]. Nature Reviews Microbiology,2009,7(9):623-628.
[12]李秀娟,赵 冬,潘 琢,等. 29种毒力基因在91株食源性单核细胞增生李斯特氏菌中的分布[J]. 中国人兽共患病学报,2017,33(11):972-978.
[13]Vázquezboland J A,Kuhn M,Berche P,et al. Listeria pathogenesis and molecular virulence determinants[J]. Clinical Microbiology Reviews,2001,14(3):584-640.
[14]伊娜娜. 不同毒力单增李斯特菌LIPI-1全序列分析及致病性差异研究[D]. 哈尔滨:东北农业大学,2012.
[15]乌日娜,郭邦成,陈福生. 宁夏食源性单增李斯特菌毒力基因的分布研究[J]. 宁夏大学学报(自然科学版),2016,37(2):205-210.
[16]刘二龙,袁慕云,吕英姿,等. 单增李斯特菌三重实时荧光PCR检测的建立及其毒力基因在分离菌株中分布[J]. 中国人兽共患病学报,2016,32(5):451-456.
[17]Wiedmann M,Bruce J L,Keating C,et al. Ribotypes and virulence gene polymorphisms suggest three distinct Listeria monocytogenes lineages with differences in pathogenic potential[J]. Infection&Immunity,1997,65(7):2707-2716.
[18]亢春雨. 食源性單核增生性李斯特氏菌的分布、遗传多态性及其毒理机制研究[D]. 保定:河北农业大学,2015.
猜你喜欢
中老年保健(2022年1期)2022-08-17
云南化工(2021年6期)2021-12-21
中老年保健(2021年6期)2021-08-24
农药科学与管理(2019年6期)2019-11-23
现代食品(2016年24期)2016-04-28
华南农业大学学报(2015年5期)2015-12-04
现代检验医学杂志(2014年5期)2014-02-02
小小说月刊(2013年6期)2013-05-14
