
弱光淹涝后强光对‘极高’葡萄叶片PSI和PSII的影响
2019-11-21 05:32徐玉涵傅彩琦卢蕾翟衡杜远鹏
中外葡萄与葡萄酒 2019年6期
徐玉涵,傅彩琦,卢蕾,翟衡,杜远鹏*
(山东农业大学园艺科学与工程学院,山东泰安 271018)
我国属于大陆季风性气候,雨热同季,因此淹涝是我国夏季主要的自然灾害。阴雨涝害后经常突然放晴又造成强光胁迫。研究表明,淹涝主要影响植物的PSII[1-2],进而加速叶片衰老并减少生物量的积累[3]。强光会造成植物叶片光系统功能的抑制[4]。孙永江等[5]的研究表明,植物在遭受逆境后产生光抑制,此时叠加强光交互作用后会产生附加效应,加剧对植物光系统的伤害。适应了弱光环境的叶片在突然遇到强光环境后光合受到显著抑制甚至死亡[6-8]。Eiia等[9]认为,植株由淹涝-弱光环境到强光环境时,叶片的光合机构在强光下产生较高水平的活性氧,使植物受到较大的氧化伤害,导致光合机构的光抑制。
田间调研发现,淹涝1周内葡萄便出现淹涝症状。但很多研究者通过模拟淹涝胁迫发现,植物出现淹涝症状的时间远远超过田间。李艳等[10]对葡萄进行淹水胁迫发现,在胁迫达到12 d时葡萄叶片的叶绿素含量、PSII活性等指标才会出现显著性变化,在淹水前期葡萄叶片的表观症状并没有发生显著性变化。探究原因发现,田间淹涝一般伴随雨后连阴天突然放晴的情况,发生淹涝症状的时间多在放晴后。而试验模拟淹涝一般都在正常光照条件下,由此说明强光能够加剧阴天淹涝发生程度,为探讨葡萄淹涝后强光对两个光系统的影响。本试验以葡萄栽培品种‘极高’为试材,前期弱光下淹涝模拟连续阴雨天发生,后期淹涝叠加强光胁迫模拟雨后的强光,研究淹涝后强光对葡萄叶片光合特性响应,以期为葡萄淹涝恢复提供理论基础。
1 材料与方法
1.1 材料与处理
试验于2018年8月在山东农业大学园艺实验站葡萄园基地进行,选取葡萄品种‘极高’为试验材料,种植于直径23.5 cm、高14 cm的塑料花盆中,栽培基质为沙:土=1:2。待生长到7~8片叶时,选取长势一致的植株16盆,放置于长50 cm、宽40 cm、高50 cm的白箱子中,进行淹水至塑料盆上沿,对照8株正常管理。淹涝时模拟连续阴雨天气,放置于室外遮阳网下(采用光强仪测定遮阳网下一天中光照强度为300~400 μmol/(m2·s)),处理第14天开始将淹水处理植株分为等量两部分,其中8株连同白箱子撤除遮阳网,于第14、15、16天进行连续强光胁迫淹水处理(自然光照,采用光强仪测定遮阳网下一天中光照强度,达到强光胁迫1400~1600 μmol/(m2·s)的时间为每天12时至16时,总共6 h),其余8株继续弱光条件下(遮阴网下)淹水处理。于开始淹涝后的0、3 d、6 d、9 d、14 d、15 d、16 d测定植株中部节位长势一致的叶片的叶绿素荧光参数。
1.2 叶绿素荧光参数的测定
参考邢浩等[11]方法,用连续激发式荧光仪(Handy PEA,Hansatech,英国)测定OJIP曲线,包括初始荧光(Fo)、在300 µs时的荧光(Fk)、在2 ms时的荧光(Fj)、在30 ms时的荧光(Fi)、最大荧光(Fm)、PSII最大光化学量子产量(Fv/Fm)和PSII潜在活性(Fv/Fo)。测定前叶片预先暗适应15 min,通过对OJIP曲线的分析得出反映PSII供体侧、受体侧及PSII反应中心功能的参数。用Wk=(Fk-Fo)/(Fj-Fo)表示受体侧放氧复合体受伤害程度;用捕获的激子将电子传递到电子传递链中QA-下游其他电子受体的概率φEo=(Fm-Fj)/(Fm-Fo)来表示PSII受体侧电子传递的功能;用单位面积有活性反应中心数量RC/CSm=Fm×φPo×(Vj/Mo),表示PSII反应中心变化。
参考Klughammer等[12-13]的方法进行叶绿素荧光光响应曲线的测定。采用Dual-PAM100荧光仪(Walz,德国)进行饱和脉冲分析,在人工光源(800 μmol/(m2·s))下适应1 h,测定叶片在稳定光源下的荧光参数,之后测定叶片暗适应15 min后的初始荧光产量(Fo)和最大荧光产量(Fm),其中Fv=Fm-Fo。处理结束后,进行荧光光响应曲线的测定,照射样品30 s后打开饱和脉冲光进行最大荧光产量(F'm)及光下最大量子产量(P'm)的测量,Pm为最大光氧化P700+。按照如下公式进行荧光参数的计算。
PSII实际光化学效率Y(II)=(F'm-Fs)/F'm。
非调节能量耗散的量子产率Y(NO)=1/[NPQ+1+qL(Fm/Fo-1)]。
可调节能量耗散的量子产率Y(NPQ)=1-Y(II)-1/[NPQ+1+qL(Fm/Fo-1)]。
PSI的有效量子产率Y(I)=1-Y(ND)-Y(NA)。
PSI供体侧限制引起的非光化学能量耗散的量子产量Y(ND)=1-P700red。
PSI受体侧限制引起的非光化学能量耗散的量子产量Y(NA)=(Pm-P'm)/Pm。
PSI激发能分配系数α=f/(1+f),PSII激发能分配系数β=1/(1+f),f=(F'm-Fs)/(F'm-F'o),用β/α-1表示PSI和PSII间激发能分配的不平衡性[14]。
2 结果与分析
2.1 淹涝和强光交互作用下叶片表观症状
淹涝胁迫16 d及淹涝和强光交互作用3 d后葡萄叶片外观出现明显的症状。在淹涝胁迫前期未表现出明显的症状,至淹涝胁迫处理16 d时,叶片出现萎蔫现象,表现出叶尖、叶缘或叶脉黄化且有轻微的焦枯现象。进行淹涝-强光交互作用后,叶片淹涝症状显著加剧,在交互胁迫处理3 d时,叶缘、叶尖焦枯现象严重,部分叶片出现脱落现象。
2.2 淹涝和强光交互作用对葡萄叶片PSII及PSI的影响
2.2.1 淹涝和强光交互作用对葡萄叶片PSII和PSI活性的影响
PSII最大光化学量子产量(Fv/Fm)可以反映PSII受胁迫程度。从图1A可以看出,淹涝处理3 d就已经出现显著性降低,比对照降低了11%;淹涝胁迫至14 d、16 d时Fv/Fm分别显著降低了23.2%、36.5%;淹涝胁迫至14 d后叠加强光胁迫处理进一步加剧了Fv/Fm下降幅度,处理至14 d、16 d与对照相比,显著下降了29.6%、51.1%。
最大光氧化P700+(Pm)可以反映PSI活性。由图1B可知,淹涝处理3 d,Pm开始降低,但是降低程度不显著,处理至9 d与对照相比降低了0.7%,未达到显著性差异。直至淹涝14 d、16 d时才与对照出现显著下降(8.8%、36.7%);淹涝胁迫至14 d后叠加强光胁迫处理后进一步加剧了Pm下降幅度,胁迫至14 d、16 d与对照相比,显著下降了16.6%、48.8%。说明淹涝至第14天导致‘极高’叶片PSI发生明显光抑制,且叠加强光胁迫后加剧了光抑制程度。
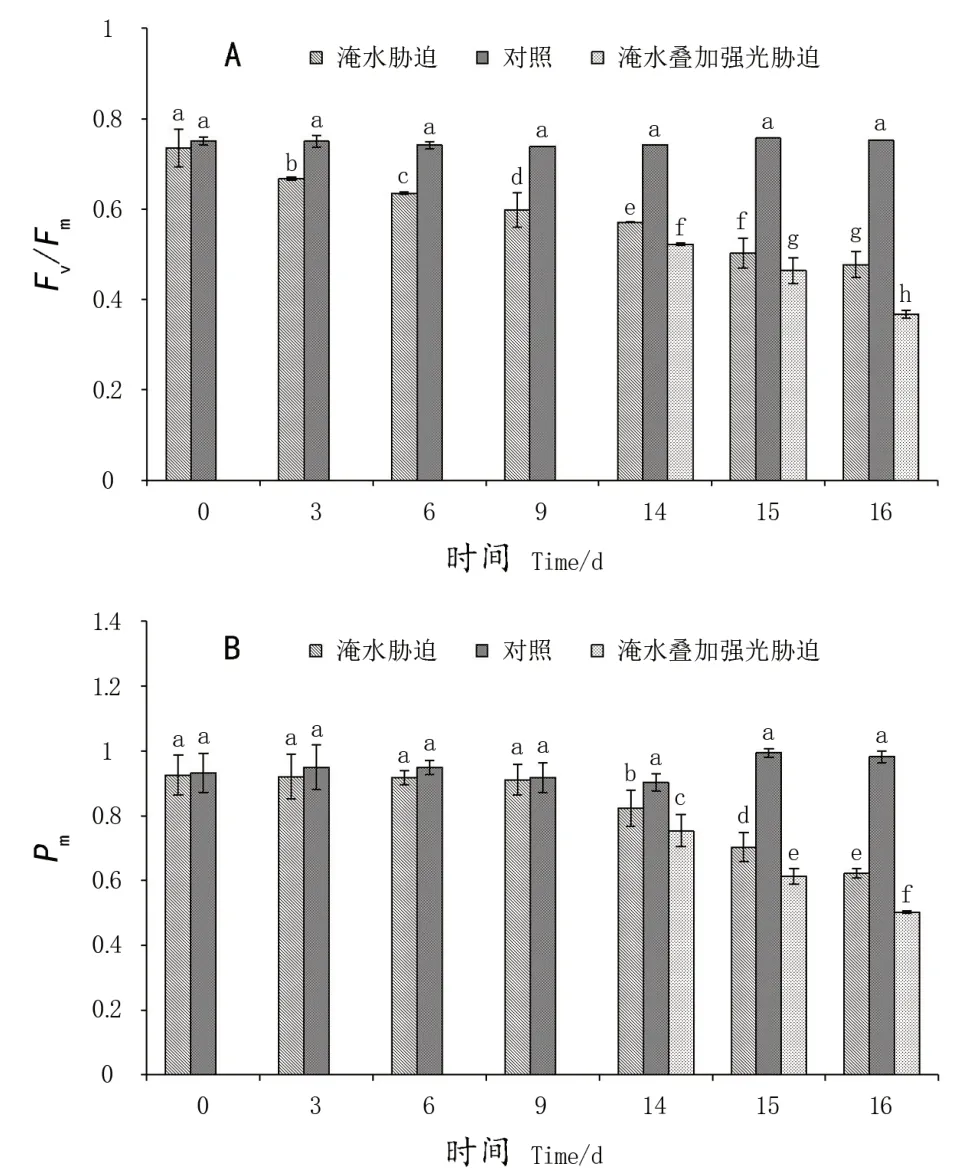
图1 淹涝和强光交互作用对葡萄品种Fv/Fm、Pm的影响Figure 1 Interaction effect of water-logging and high lightstress on Fv/Fm and Pm in grapevine leaves
2.2.2 淹涝和强光交互作用对葡萄叶片PSII供体侧、活性反应中心数量、受体侧的影响
K点相对可变荧光(Wk)表示PSII供体侧放氧复合体受伤害程度[15]。随着淹涝处理时间的增加,Wk值逐渐升高。与对照相比淹涝至14 d、16 d,显著增加了19.8%、38.6%;淹涝胁迫至14 d后叠加强光胁迫处理进一步加剧了其上升幅度,与对照相比,处理至14 d、16 d显著增加了23.3%、67.5%(图2A)。表明‘极高’叶片PSII供体侧受到了伤害,叠加强光胁迫后,‘极高’叶片PSII供体侧受到伤害加剧。
淹涝处理第3天时单位面积有活性反应中心数量(RC/CSo)就比对照出现显著性降低,淹涝14 d、16 d时单位面积有活性反应中心数量(RC/CSo)与对照相比降低了24.2%、26.1%;淹涝胁迫至14 d后叠加强光胁迫处理进一步加剧RC/CSo下降幅度,与对照相比,处理至14 d、16 d显著降低了32.7%、45.3%(图2B)。
由图3C可知,随着淹涝胁迫处理时间的延长,PSII受体侧电子传递的效率(φEo)逐渐降低。淹涝至第3天显著降低,至14 d、16 d,φEo与对照相比,显著降低了20.3%、30.3%;淹涝胁迫至14 d后叠加强光胁迫处理后下降幅度加剧,处理至14 d、16 d与对照相比显著降低了25.5%、46.1%。由此说明,淹涝胁迫3 d后就显著地抑制了PSII光合电子传递链,淹涝叠加强光胁迫进一步抑制了‘极高’叶片PSII光合电子传递链。
2.3 淹涝和强光交互作用对光系统能量分配及热耗散的影响
2.3.1 淹涝和强光交互作用对光系统能量分配的影响
淹涝以及淹涝叠加强光胁迫明显改变了PSII和PSI的能量分配。随着淹涝时间增加,PSII实际光化学效率Y(II)及不可调节的热耗散Y(NO)逐渐降低(图3D和3F),可调节的热耗散Y(NPQ)逐渐升高(图3E),淹涝至第3天便出现显著性变化。淹涝胁迫至14 d后叠加强光胁迫处理后使得PSII的Y(II)下降速率加剧,Y(NPQ)变为下降趋势,Y(NO)变为上升趋势,表明PSII活性受到抑制明显,且叠加强光胁迫后,依赖光保护的热耗散机制并不能耗散掉所有的光能,随着强光胁迫时间的延长,过剩光能的比例越来越高,叶片光抑制加剧。
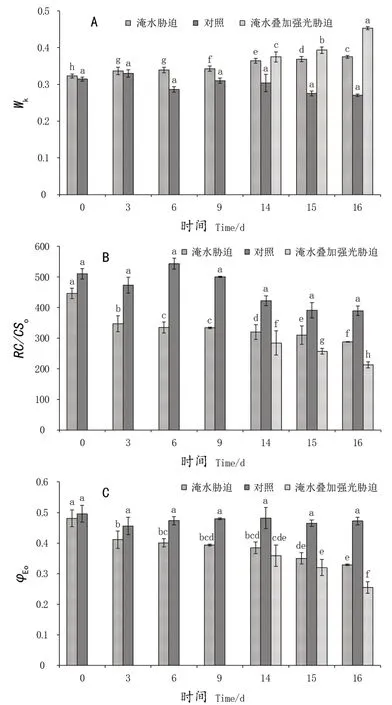
图2 淹涝和强光交互作用对葡萄叶片供体侧(Wk)、反应中心(RC/CSm)及受体侧(φEo)的影响Figure 2 Interaction effect of water-logging and high light stress on donor side (Wk),reaction center (RC/CSm) and receptor side (φEo) of PSII in grapevine leaves
随着淹涝至第6天PSI实际光化学效率Y(I)及氧化态P700+显著降低(图3A和3C),PSI受体侧在淹涝至第6天受抑制程度便显著增加(图3B),淹涝胁迫至14 d叠加强光胁迫处理后,进一步加剧了Y(I)及氧化态P700+降低速率,PSI受体侧受抑制程度加剧。进一步表明淹涝叠加强光胁迫导致了PSI光抑制的发生。
淹涝胁迫第3天,‘极高’叶片PSII和PSI间激发能分配不平衡性(β/α-1)显著性增大,淹涝至14 d、16 d时处理的β/α-1比对照显著增加了53%、230.6%;淹涝胁迫至14 d叠加强光胁迫处理后,β/α-1上升速率加剧,淹涝叠加强光处理至14 d、16 d分别比对照升高了73.5%和249%(图4)。
2.3.2 淹涝和强光交互作用对热耗散的影响
非光化学淬灭(NPQ)表示PSII天线色素吸收的光能不能用于光合电子传递而是通过光保护途径以热的形式耗散掉的光能所占的比例[5,16-17]。从图5中可以看出,淹涝胁迫至第6天时,NPQ出现显著性上升。淹涝胁迫至14 d叠加强光胁迫后NPQ进一步显著上升,表明光系统通过提高热耗散来进行光保护。
3 讨论与结论
叶绿体是植物进行光合作用的细胞器,对叶片的叶绿素荧光参数进行分析能够反映胁迫条件下植物光合器官的伤害程度。当植物吸收的光能超过所能利用的光能时,过剩光能便会导致光合机构的光抑制,甚至光破坏[18-19]。本试验发现淹涝后PSII最大光化学效率呈线性下降,但在淹涝胁迫后期下降趋势渐缓。淹涝强光交互作用下,PSII的最大光化学效率较同时期单因素淹涝胁迫低,且随着淹涝强光胁迫时间的增加,下降显著。这说明弱光环境PSII反应中心的压力较小,因此仅仅在淹涝胁迫下,植株开始逐渐适应高湿、弱光的环境,但是突然将植株移至高光强下,对已经受到抑制的PSII光合反应中心造成压力,使得过剩激发能增加,这些能量以电子的形式存在于电子传递链中并形成活性氧,过量的ROS既能直接破坏PSII[20],也可以抑制D1蛋白再循环[21],最终导致光破坏。
有研究表明,PSII到PSI的电子传递速率的降低保护PSI免受进一步的光抑制[22]。但也有研究表明,当PSII的活性被抑制至一定程度,PSII到PSI的电子流被严重阻断,PSI的受体侧无法正常的接受电子,使得PSI的光合作用无法正常完成[23]。本试验发现,淹涝胁迫对PSI的最大光氧化P700+(Pm)前期影响较小,在处理6 d才出现显著性影响,淹涝强光交互胁迫处理后,Pm显著下降。造成这种现象的原因可能有两方面:一是在淹涝胁迫下,PSII活性受到了抑制,降低了PSII到PSI的电子传递速率(图2C),使得PSI的活性虽然受到了一定程度的抑制,但是影响不显著。淹涝强光交互作用后,叶片两个光系统间激发能分配不平衡性显著增加(图4),PSII活性中心受抑制严重,PSI和PSII的光能分配出现严重失衡,叶片吸收的光能不能有效地从PSII传递到PSI,导致由PSII传递给PSI的电子大量减少,PSI不能有效的进行光合作用;二是在淹涝强光胁迫下,产生大量无法有效利用的光能,这些能量以电子的形式存在于电子传递链中并形成活性氧,过量的活性氧会攻击PSI活性中心,抑制PSI的活性。二者协同作用,造成PSI最大光氧化P700+受到抑制。
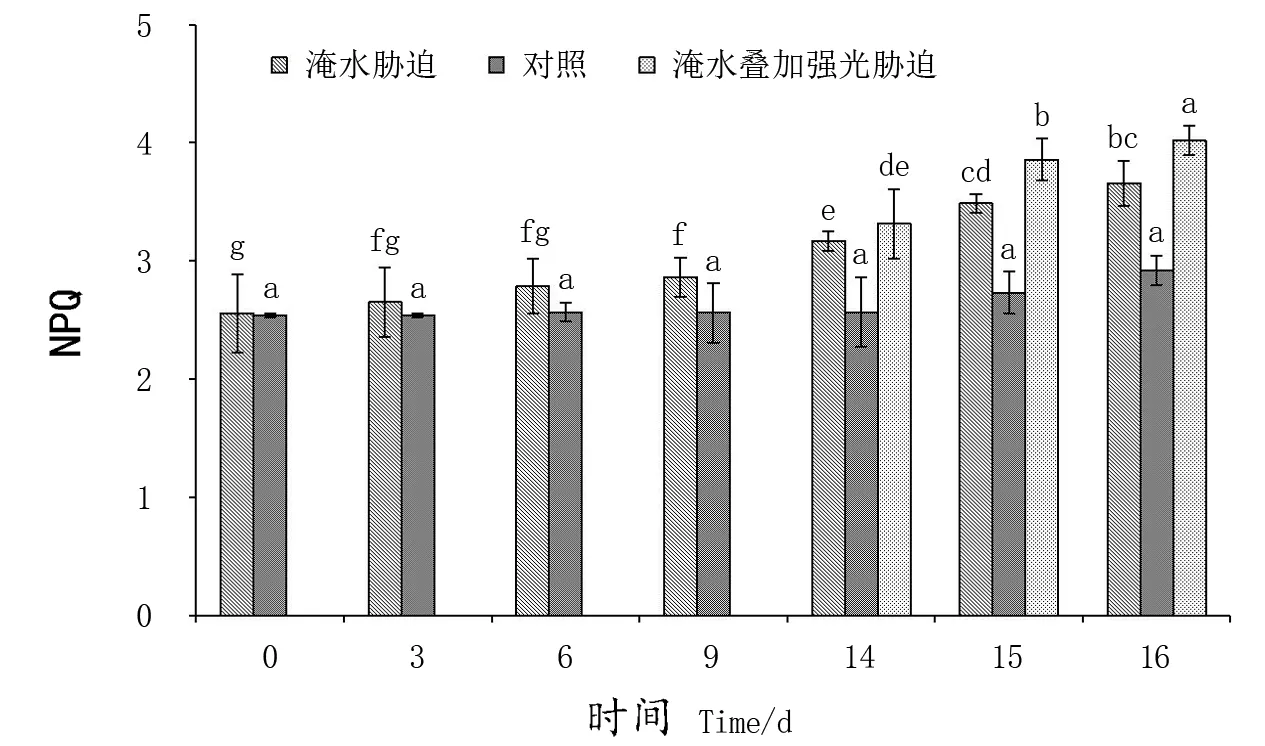
图5 淹涝和强光交互作用对葡萄品种非光化学淬灭(NPQ)的影响Figure 5 Interaction effect of water-logging and high light stress on non-photochemical quenching (NPQ) in leaves
NPQ表示PSII天线色素吸收的光能不能用于光合电子传递,而是通过光保护途径以热的形式耗散掉的光能所占的比例,反映PSII处理过剩光能的能力。因此,NPQ可作为光保护能量耗散的重要指标。试验结果表明,NPQ随着淹涝及淹涝后强光胁迫时间的增加不断上升,说明淹涝强光胁迫下,光系统通过提高热耗散来进行光保护。且淹涝强光交互胁迫3 d后使得PSII的可调节能量耗散的量子产率Y(NPQ)变为下降趋势,不可调节能量耗散的量子产率Y(NO)变为上升趋势,表明淹涝胁迫处理的叶片还能通过热耗散来进行自我调节,但淹涝强光交互胁迫后,依赖光保护的热耗散机制并不能耗散掉所有的光能,随着强光胁迫时间的延长,过剩光能的比例越来越高,叶片光抑制加剧。
综上所述,淹涝胁迫使葡萄叶片光系统II和光系统I的活性产生了抑制作用;葡萄叶片在淹涝过程中受到强光照射后,强光导致了PSII的光抑制和反应中心失活,进一步加剧了PSI的抑制。
猜你喜欢
全科护理(2022年14期)2022-05-16
中国医药导报(2021年35期)2022-01-20
现代盐化工(2021年3期)2021-08-27
长江丛刊(2020年13期)2020-11-19
实用器官移植电子杂志(2020年4期)2020-08-31
婚姻与家庭·婚姻情感版(2019年6期)2019-06-03
发明与创新·大科技(2017年7期)2017-07-17
中国经济周刊(2016年18期)2016-05-14
新课程·下旬(2015年10期)2015-10-21
中国美容医学(2004年1期)2004-08-11
