
海南岛甘什岭热带低地雨林棕榈藤空间点格局分析
2019-12-31 08:02徐瑞晶漆良华程昌锦
生态学报 2019年22期
胡 璇,徐瑞晶,2,漆良华,*,彭 超,丁 霞,程昌锦,郭 雯,张 建
1 国际竹藤中心,北京 100102 2 海南三亚竹藤伴生林生态系统国家定位观测研究站,三亚 572000
种群分布格局是种群个体在水平空间上的配置或分布状况[1],受种群特性、种群关系和环境条件的综合影响[2],是多种生态过程对群落中各组分在不同空间和时间尺度上综合 作用的结果[3]。分析种群分布格局有助于揭示种群的生物学特性、生态学过程及其与环境因子之间的关系[3-5]。
植物群落的空间分布是多种生态过程对群落中各组分在不同空间和时间尺度上综合作用的结果,具有一定的规律[3],植被类型、植物种类和数量、种间变异、环境以及外力干扰等都会对群落构建产生影响[6-7],且存在于整个群落生态过程中。分析各组分在群落中的空间分布有利于揭示物种在群落演替动态过程中的作用和潜在规律,具有重要的生态学意义。
关于空间分布格局的探索众学者已在热带雨林、针阔混交林、沙漠荒地[8-10]等群落物种中进行了大量研究,其中热带雨林一直为研究的热点,研究对象多集中于乔木优势种和灌草,但对藤本植物研究极少[11]。棕榈藤是省藤族(Calameae)藤类植物,隶属于棕榈科(Palmae)中省藤亚科(Calamoideae),是热带森林的重要组分。其被俗称为“藤条”的藤茎是重要的非木材林产品之一,市场需求量大,因此棕榈藤具有重要的经济价值和发展前景,国际上对棕榈藤资源的研究和开发也较为重视。目前,对于棕榈藤的相关研究多集中于分布特征以及生活习性[12-13]等方面,而对于其在群落中的分布及种间关系鲜有报道,仅Souza和Martins[14]针对棕榈植物(palms)进行了一定涉及,相关研究亟待加强。空间分布格局的研究方法很多,但依靠单一尺度的若干样方研究植物种群(群落)的空间分布格局的传统方法,只能对特定空间尺度上的分布格局得到具体认识[15-17]。1977 年Ripley[18]提出了空间点格局分析法,其后,O-ringO(t)、G(t)等函数分析法相继在RipleyK(t)函数分析法的基础上发展而出[19-20]。点格局分析方法由于克服了单一尺度下分析种群分布格局的不足,近年来已为国内外不少学者所运用,该方法可以分析各种尺度下的种群格局和种间关系,检验能力较强[5]。因此以海南岛甘什岭热带低地雨林白藤(Calamustetradactylus)、大白藤(C.faberii)、华南省藤(C.rhabdocladus)、黄藤(Daemonoropsmargaritae)和小钩叶藤(Plectocomiamicrostachys)5种棕榈藤种群为研究对象,采用点格局分析方法对棕榈藤种间和各生长阶段下的空间分布格局及关联性进行分析,以期深入认识各棕榈藤种群的空间格局特征及其随尺度变化的规律,为海南岛甘什岭热带雨林的生物多样性保护与维持、退化生态系统的恢复等提供理论依据和参考。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于海南省甘什岭自然保护区内的试验区,地处三亚市与保亭县南部交界处109°34′—109°42′E,18°20′—18°21′N,面积2103.44 hm2,其核心区和缓冲区分别占地1166.07 hm2、678.50 hm2,试验区占地为258.87 hm2。研究区为低山丘陵地貌,海拔50—681 m,土壤母质为花岗岩,岩层裸露率10%,土壤含砂量为20%左右。研究区干湿季分明,属热带海洋季风气候,年降雨量约为1800 mm,5—10月为雨季,其余时间为旱季,年平均气温25.4℃,年平均日照时数约为2563 h[21]。试验区属热带低地次生雨林,主要树种有无翼坡垒(Hopeareticulate)、青皮(Vaticamangachapoi)、阿芳(Alphonseamonogyna)及琼南柿(Diospyroshowii)等。
1.2 研究方法
1.2.1样地设置与调查
(1)样地设置
2015年11月至2016年1月,对海南岛甘什岭自然保护区的试验区进行实地调查,选择一块具有典型性、代表性的热带低地雨林,设置100 m×100 m样地1个。样地面积1 hm2,基准点地理坐标为109°40′18.76″E,18°23′06.56″N。样地总体上西高东低,海拔约为261 m,坡度0—26°。采用相邻格子样方法,将样地分成25个20 m×20 m的大样方。
(2)棕榈藤调查
分藤种对幼苗(<0.5 m)、幼藤(0.5—2.5 m)、成熟藤(>2.5 m)[22]以及株丛进行调查,株丛以萌蘖丛植株≥1株为基准。调查过程中对每株棕榈藤进行插签标记,防止漏测或重测,分别记录其种类和个体数量。生长早期的棕榈藤从基部至展开叶最高处的垂直高度作为植株高度,抽茎后则以抽茎基部至茎顶端为长度;对丛生棕榈藤进行分株测量,记录高度或长度。本研究中,白藤、大白藤、华南省藤、黄藤和小钩叶藤5种棕榈藤幼苗形态差异较明显,通过叶片形态和羽片数量对其进行种类鉴定。
1.2.2数据处理与统计分析
采用单变量和双变量的O-ring分析方法分析海南岛甘什岭热带低地雨林白藤、大白藤、华南省藤、黄藤和小钩叶藤5种棕榈藤种群种内和种间的空间分布和关联性。O-ring分析方法与以往国内外有关物种空间格局研究时多采用的Ripley(K)函数[3]相比,避免了积累效应,在做大尺度格局分析时,能避免受到小尺度积累效应的影响,可以更真实地反映种群任意尺度的空间分布格局,因而近年来得到了广泛的应用[23-24]。
在单变量O-ring分析种,用成对相关函数g(r)来计算种群多个特定尺度上的空间格局,其计算公式如下:
g(r)=(2πr)-1dK(r)∕dr
若g(r)高于置信区间上限,则该植物在该距离为聚集性分布;若g(r)在置信区间之间,则为随机分布;若g(r)低于置信区间的下限,则为均匀分布。
在成对相关函数进行双变量点格局分析时,函数为:
g12(r)=(2πr)-1dK12(r)∕dr
若g(r)高于置信区间上限,则种间在距离r处显著正关联;若低于置信区间的下限,则种间在距离r处显著负关联;若在置信区间之间,则表明两个种之间相互独立或没有显著关联性[25]。
采用Programita 软件(2008版)完成数据分析过程,空间尺度大小为0—50 m,经100次Monte-Carlo模拟得到99%的置信区间[26]。此外,采用Excel 2007和R 15.0对数据进行前期的基础处理和分析及作图。
2 结果与分析
2.1 棕榈藤的空间分布格局
2.1.1不同藤种棕榈藤的种群密度分布
如表1所示,样地内不同种的棕榈藤种群密度不同,其中华南省藤和黄藤种群密度较大,分别为241株/hm2和233株/hm2,其占棕榈藤总株数的比例分别为34.13%和33.00%;白藤和大白藤种群密度次之,均为83株/hm2,其占棕榈藤总株数的比例为11.76%;小钩叶藤的种群密度最小,为66株/hm2,其占棕榈藤总株数的比例为9.35%。

表1 棕榈藤种群密度及分布状况Table 1 Population density and distribution of rattan
表中各棕榈藤的密度是通过每丛种萌蘖出的株树来算的,故单位为“株/hm2”
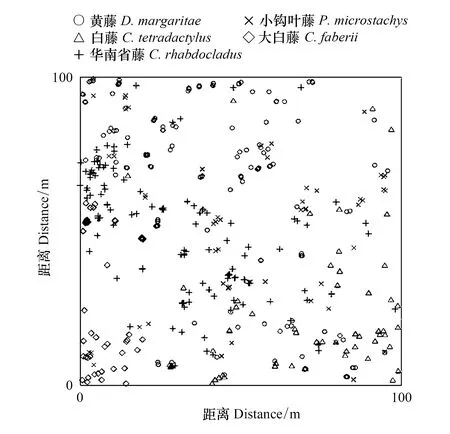
图1 5种棕榈藤在1 hm2样地内的分布点图Fig.1 Mapped point pattern of 5 kinds of rattan in 1 hm2 plot
通过表1、图1和图2可看出,不同棕榈藤在样地中的分布情况不同,其中黄藤、白藤、华南省藤和大白藤较为明显,聚集程度较高;小钩叶藤分布较为随机。白藤在样地内的聚集分布程度最为明显,主要分布在坐标为40≤x≤100,0≤y≤40的范围内;黄藤、华南省藤和大白藤的聚集程度次之。黄藤在样地内大多聚集分布在坐标为0≤x≤60,70≤y≤100的范围内;华南省藤大多聚集在坐标为0≤x≤40,10≤y≤70的范围内;大白藤大多聚集在坐标为0≤x≤20,40≤y≤60的范围内。小钩叶藤的分布情况较为随机,样地中各坐标范围内均有分布。不同藤种的棕榈藤在样地内的分布状况有差异,这可能是由于不同藤种对环境因子的响应不同造成的。
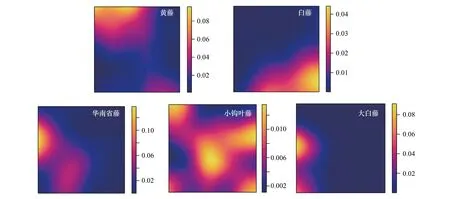
图2 5种棕榈藤在1 hm2样地内的分布热图Fig.2 Distribution thermal map of 5 kinds of rattan in 1 hm2 plot
2.1.2不同藤种棕榈藤的空间分布格局
对不同的棕榈藤进行独立的分布格局检测(图3),其中,黄藤在总体尺度上表现出高度聚集,仅在2.5、5.5—6.5、13.5、16.5、22.5、36.5、47.5—49.5 m尺度上表现出随机分布,在10.5、19.5、41.5—44.5 m和50 m尺度上表现出均匀分布;白藤在0—27.5 m尺度上表现出明显的聚集分布,仅在5.5 m尺度上表现出随机状态,在28.5—48.5 m尺度上聚集分布和随机分布交替出现,仅在49.5 m尺度上表现出均匀分布;华南省藤在0—13.5 m的小尺度上表现出非常明显的聚集分布,此后,随尺度增加逐渐出现聚集分布、均匀分布和随机分布交替出现的过渡区间,在46.5 m后的大尺度上呈现较为稳定的均匀分布状态;小钩叶藤仅在0—1.5 m的小尺度上表现出聚集分布,其后随着尺度的增长几乎全表现为随机分布,只在4.5 m尺度上表现出暂时的聚集分布,在31.5 m和37.5 m尺度上表现出均匀分布;大白藤在0—18.5 m的小尺度上呈现出非常明显的聚集分布,仅在16.5 m时暂时表现出随机分布,在19.5—32.5 m的区间随机分布和均匀分布交替出现,其后,在33.5—50 m尺度上聚集现象再次变得明显,仅在34.5 m、46.5—47.5 m尺度上短暂地表现出随机分布的状态。可见,不同藤种的棕榈藤随着尺度的变化,其分布格局也各有不同,小尺度上5种棕榈藤都呈现出聚集分布的状态,其后随着尺度的变化,种群的聚集程度逐渐减弱,最终呈现出均匀分布或随机分布,或聚集分布、均匀分布、随机分布三种状态交替出现的现象。这可能是由于不同藤种对环境资源,如光、水、空间及营养元素的需求和响应不同,随着种群密度的增加,种内和种间的相互作用增强,从而造成了自疏或它疏作用。5种棕榈藤中小钩叶藤在1.5 m尺度后迅速转变为随机分布,与其他4种藤分布情况差异较大,这可能与其自身的生存策略及其对环境的适应性有关,未来应加强对棕榈藤和环境因子关系的研究。

图3 5种棕榈藤点格局Fig.3 Spatial patterns of 5 kinds of rattan palms
2.2 藤种间的空间关联
棕榈藤的分布热图(图2)及不同棕榈藤的空间关联性分析(图4)表明:不同藤种间空间随尺度变化的关联性各不相同,差异较大。其中黄藤和白藤更多表现出不相关,仅在0—0.5 m、14.5 m、24.5 m尺度上表现出正相关,其他尺度上不相关和负相关情况穿插出现,随着尺度的增加,负相关现象越来越明显;黄藤和华南省藤0—20.5 m的小尺度下多表现出负相关的现象,仅在个别尺度上偶尔穿插正相关和不相关,其后,在22.5—50 m的大尺度上表现出不相关和正相关交替出现的现象,并且正相关现象更为显著;黄藤和小钩叶藤在整体尺度上更多表现出显著的不相关,仅在极个别尺度上表现出瞬时的正相关或者负相关;黄藤和大白藤在0—19.5 m的小尺度上表现出显著的负相关,仅在个别尺度上表现出不相关,其后,在20.5—50 m的大尺度上正相关、不相关和负相关交替出现,但更多的还是正相关;白藤和华南省藤随着尺度的增加多表现出负相关,仅在0—1.5 m、13.5—14.5 m的个别尺度上短暂地表现出正相关;白藤和小钩叶藤在0—50 m尺度上大部分表现出不相关,仅在个别尺度上表现出正相关或负相关;白藤和大白藤在空间分布上表现出显著的不相关;华南省藤和小钩叶藤大部分呈不相关,仅在4.5—13.5 m的小尺度上表现出正相关、不相关和负相关穿插出现的情况,其后,随着尺度的增长,逐渐转变为不相关和负相关交替出现,总体还是不相关现象居多;华南省藤和大白藤总体上呈现出显著的正相关关系,在3.5—33.5 m的尺度上正相关现象明显,之后随着尺度的增加,逐渐出现正相关和不相关交替出现的状况,偶尔表现出负相关;小钩叶藤和大白藤在0—37.5 m尺度上表现出负相关和不相关交替出现的现象,随着尺度的增加,表现出短暂的正相关现象,其后又逐渐趋于不相关。总体看来,黄藤和白藤、白藤和华南省藤、白藤和大白藤之间表现出明显的负相关关系,说明它们种间有一定的互斥作用;华南省藤和大白藤在小尺度上表现出明显的正相关关系,之后,随着尺度的增加逐渐趋于不相关,这说明在小尺度上它们之间存在着某种互利关系。在今后的研究中,棕榈藤的共存机制值得探究。

图4 不同棕榈藤藤种间的空间关联性Fig.4 Associations of different species of rattan palms
3 结论与讨论
我国为棕榈藤分布的北缘地区,棕榈藤分布较为丰富。众学者先后对中国棕榈藤植物的分类、地理分布及区系等进行了深入研究,但由于棕榈藤植物野外调查采集困难、种类界定较为复杂,棕榈藤植物的种类划分一直存在着较大争议[22,27-28],2015年王慷林[29]在前人研究的基础上对棕榈藤特征进行了重新梳理,确定我国天然分布共有3属40种(含变种)。棕榈藤主要分布在我国约24°N以南的热带和亚热带地区,并形成以西南地区云南西双版纳和东南地区海南岛为中心的两大分布区,从属水平上来看海南岛的棕榈藤分布最多[30]。棕榈藤作为重要的非木材林产品,具有很高的经济价值,对于其的相关研究亟待加强。
植物种群的空间格局分布决定于植物本身的生物学特性和环境两方面共同作用结果[18],体现了种群结构的合理性,也是种群和群落动态和稳定性的标志[31]。众多学者对同一群落内不同植物种群的空间分布格局及其空间关联性进行了研究,对更深层次地理解植物本身的生态学特性及种群动态和稳定性变化具有重要意义[25,32]。
本研究结果充分体现了种群空间格局的尺度依赖性[23]。海南岛甘什岭热带低地雨林棕榈藤种群的空间格局与空间尺度有着密切的关系,在小尺度上,5种棕榈藤种群都更倾向于聚集分布,这可能是因为聚集分布有利于其相互庇护,从而抵御外界环境影响,以提高存活几率;随着尺度的增长,各棕榈藤种群主要趋向于随机分布,形成不同的空间分布格局,这可能与种间竞争过程中种群自调节机理不同有关,主要是物种与环境长期相互适应和相互作用的结果[33];但在大尺度下,华南省藤表现出不太明显的聚集分布,大白藤表现出小程度的聚集分布,这可能与其本身的生物学特性和环境适应性有关。
不同棕榈藤藤种间的空间关联性分析结果显示,各藤种间的空间关联情况不一样,没有统一的结果,其中在整体尺度上表现出明显负关联的是白藤和黄藤、白藤和大白藤,这可能是因为它们对生存空间及养分的需求相同或相似,存在明显的竞争关系[34];呈现出明显正关联的是华南省藤和大白藤,这可能是由于它们在生长过程中相互庇护,用于抵御动物捕食和热带雨林内多变的外部环境;呈现出明显不相关的是黄藤和小钩叶藤以及白藤和小钩叶藤,这说明它们对环境和生存空间的依赖没有冲突性,能做到共存。鉴于各藤种间的空间关联情况不一样,热带雨林内棕榈藤的物种共存机制有待进一步研究。
目前,对于棕榈藤的相关研究多集中于分布特征以及生活习性[12-13]等方面,对于其在群落中的分布及其与种间关系鲜有报道,仅Souza和Martins[14]针对棕榈植物(palms)进行了一定涉及。这与棕榈藤的种类界定较为复杂和分布区域较小有关,但更重要的是,棕榈藤抽茎后刺太多,攀援现象也十分明显,测量和取样过程非常艰难。棕榈藤作为重要且优质的非木材林产品,对其进行研究很有必要。本文对海南岛甘什岭热带雨林自然分布的5种棕榈藤空间分布格局和关联性进行了分析,但对于棕榈藤生长、分布及其与环境因子的关系乃至棕榈藤的多物种共存机制方面还有待进一步深入研究。
猜你喜欢
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
天津医科大学学报(2021年1期)2021-12-05
小天使·一年级语数英综合(2020年11期)2020-12-16
蔬菜(2018年5期)2018-05-17
文理导航·科普童话(2017年4期)2018-02-10
散文诗(2017年17期)2018-01-31
中国医学影像技术(2017年11期)2017-01-16
考试周刊(2016年63期)2016-08-15
科技视界(2016年1期)2016-03-30
杂草学报(2015年2期)2016-01-04
