
中国北部小叶杨光合生理与叶片表型变异分析
2020-01-10 06:27李炳虹宋跃朋张德强张晓宇
福建农林大学学报(自然科学版) 2020年1期
李炳虹, 宋跃朋, 张德强, 张晓宇
(1.北京林业大学理学院;2.北京林业大学生物科学与技术学院,北京 100083)
小叶杨(PopulussimoniiCarr)为杨属落叶乔木,广泛分布于我国华北、东北、西北及西南地区,作为中国本土树种资源,小叶杨在林业、生态、经济等方面起着重要作用[1-3].因其具备生长快、寿命长、耐盐碱、抗干旱等特点,是生态保护和防护林工程的首选树种之一,又由于分布广泛,小叶杨存在多个变种,如垂枝小叶杨、秦岭小叶杨等,变种资源极其丰富[4-5].然而,随着气候变迁、土地荒漠化、盐碱化的加剧,小叶杨的生长环境受到了严重破坏,为保护基因资源并加强小叶杨生长发育及遗传改良研究,探究小叶杨光合生理及叶片表型变异变得尤为重要.
植物遗传变异是指在生存进化过程中包含表观变异和遗传物质改变的稳定变异[6].而叶片表型作为重要的遗传变异特征,代表着植物对自然环境的长期进化与适应性.目前,关于小叶杨的研究主要集中于人工林生长[7-9]、耐抗性分析[10-11]、基因克隆表达[12-13]等方面,而关于小叶杨光合生理及表型特征变异的研究则相对较少,且大多是关于小叶杨杂交系的对比分析,如李世峰等[14]研究了美洲黑杨与小叶杨8个杂交组合的苗高、胸径等表型性状,结果显示杂交组合在苗高、胸径、侧枝数3个性状上有较高遗传力;刘友全等[15]对比分析了川西高原青杨的变种及变异分化特性;卫尊征等[16]以华北及东北地区5个市县的192株小叶杨样本为试验材料,对比了小叶杨生理及叶片表型的变异特性及其与地理、气候等因素的关联.但是关于小叶杨自然种群在整个地理分布层面上涉及光合生理及叶片表型变异对比分析的研究则较少,且由于目前环境变化导致小叶杨天然资源不断衰减,而国内尚未建立关于小叶杨遗传种质资源的系统研究体系.因此,从地区分布层面对小叶杨光合生理及叶片表型变异情况进行摸底探究将极大程度上促进小叶杨变种和变型资源的对比分析,有助于进一步完善小叶杨遗传多样性评价和建立完备的基因资源库.
有鉴于此,为深入了解小叶杨的光合生理与叶片表型变异多样性,本文针对9个地区小叶杨叶片的10个生理及表型性状展开对比,通过采用方差分析、多重比较、相关性分析、主成分分析与聚类分析方法,探究了小叶杨叶片不同性状的变异程度及相互关联,并对不同地区进行划分,以期为小叶杨资源的遗传多样性研究及优良树种选育提供依据.
1 材料与方法
1.1 试验材料
2007年,宋跃朋等[17]采集了中国北部1233株小叶杨材料,并在中国山东省关县的克隆植物园内对其根部进行无性繁殖.本研究针对其中425株典型个体进行对比探究.这425株小叶杨个体分别来自于9个地区(河北赤城县、陕西富县、陕西高陵县、青海互助县、宁夏中宁县、陕西麟游县、河南伊川县、山西忻州市和河北承德市).
1.2 光合生理特征与叶片表型特征测量
在试验中,设定便携式气体交换系统(Li-6400xt, LiCor)的光合光子通量密度为1 600 μmol·m-2·s-1,CO2浓度为400 μmol·mol-1,并于上午9:00—11:00进行光合生理特征测量.当净光合速率维持不变(±0.1)时,记录净光合速率、气孔导度、胞间CO2浓度和水分利用率的值.为确保数据精确,测量3个叶片的光合生理特征3次(主茎顶第四至第六片叶)并计算均值.再采用便携式激光叶面积仪(CI-202)来测量6个叶片表型特征(叶面积、叶长、叶宽、叶周长、长宽比和叶形因子).
1.3 数据分析方法
计算均值与标准差获得变异系数(CV=标准差/均值).方差分析[18]中,采用完全随机试验设计模型:
yij=μ+αi+εij,i=1,2,…,k,j=1,2,…,ni
式中:yij为第i个因素的第j个观察值,μ为总体均值,αi为第i个因素的主效应,εij为误差项.k为因素个数,ni为第i个因素的观察值总数.结合方差分析使用Tukey检验[19]进行各特征的多重比较.
相关性分析中,利用Person相关系数:ρX,Y=cov(X,Y)/σXσY,度量不同特征之间的关联强弱;主成分分析中,进行特征值分解来获取各特征贡献率;
聚类分析中则利用数据相似性采用基于欧氏距离的Ward法[20]对不同地区进行划分.所有数据分析工作均使用R软件来完成.
2 结果与分析
2.1 小叶杨光合生理及叶片表型变异分析
在种群水平上(表1),小叶杨叶片净光合速率、气孔导度、胞间CO2浓度、水分利用率、叶面积、长度、宽度、周长、长宽比、叶形因子的平均值分别为14.57 μmol·m-2·s-1、0.32 mol·m-2·s-1、260.0 μmol·mol-1、3.38 μmol·mmol-1、33.52 cm2、8.30 cm、5.83 cm、40.81 cm、1.47和0.41.其中,叶片面积的变异系数最大,高达0.78,变异范围为3.91~177.95 cm2;胞间CO2浓度的变异系数最小,为0.14,变异范围为144.24~400.38 μmol·mmol-1.此外,根据样本峰度和样本偏度值,除水分利用率、叶面积、宽度和长宽比外,其余性状均近似服从正态分布.
在不同地区之间,小叶杨叶面积、周长和叶形因子具有最大的平均变异系数(表2),分别为0.52、0.51和0.61.相较于光合生理特征(0.235),小叶杨的叶片表型特征具有更大的平均变异系数(0.395).
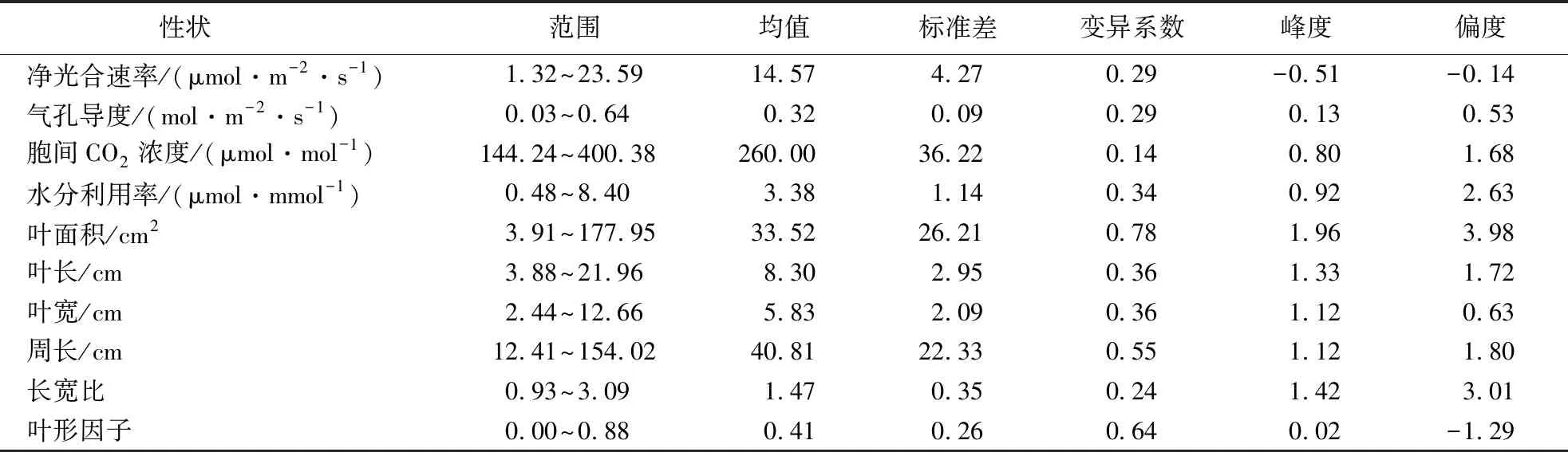
表1 小叶杨光合生理及叶片表型性状变异Table 1 Variation of photosynthetic physiological characteristics and leaf phenotypic traits of P.simonii

表2 各地区小叶杨叶片光合生理及叶片表型性状变异系数均值Table 2 Averages of variation coefficients of photosynthetic physiological characteristics and leaf phenotype of P.simonii from different regions
由此可见,在小叶杨种群中,光合生理特征保持相对稳定,而叶片表型特征则由于环境和地理因素的不同出现了较明显的变异.
2.2 方差分析与多重比较
方差分析结果表明(表3),不同地区之间,除水分利用率外,其余特征均存在极显著(P<0.001)差异.说明小叶杨群落的光合生理特征与叶片表型特征均存在丰富变异.多重比较(表4、5)结果显示,个别地区的某些特征相较于其它地区存在明显差异.例如,富县和伊川县地区小叶杨的叶面积均值高达54.39和57.42 cm2,而高陵县与中宁县的小叶杨叶面积均值仅有18.78和17.81 cm2,虽然富县与高陵县地区相近,但小叶杨叶面积相差较大,并且叶片长度均值也存在较大差距(10.80 cm>6.21 cm);此外,互助县地区小叶杨的叶周长均值为所有地区最大值63.97 cm,但叶形因子均值(0.17)却明显低于其它地区.说明不同地区之间小叶杨的叶片表型性状存在明显变异,且互助县地区的小叶杨叶片更细长,而其它地区近似为椭圆.

表3 不同地区小叶杨光合生理及叶片表型性状方差分析1)Table 3 Variance analysis of photosynthetic physiology and leaf phenotype on P.simonii from different regions
1)***代表在0.001水平上显著.
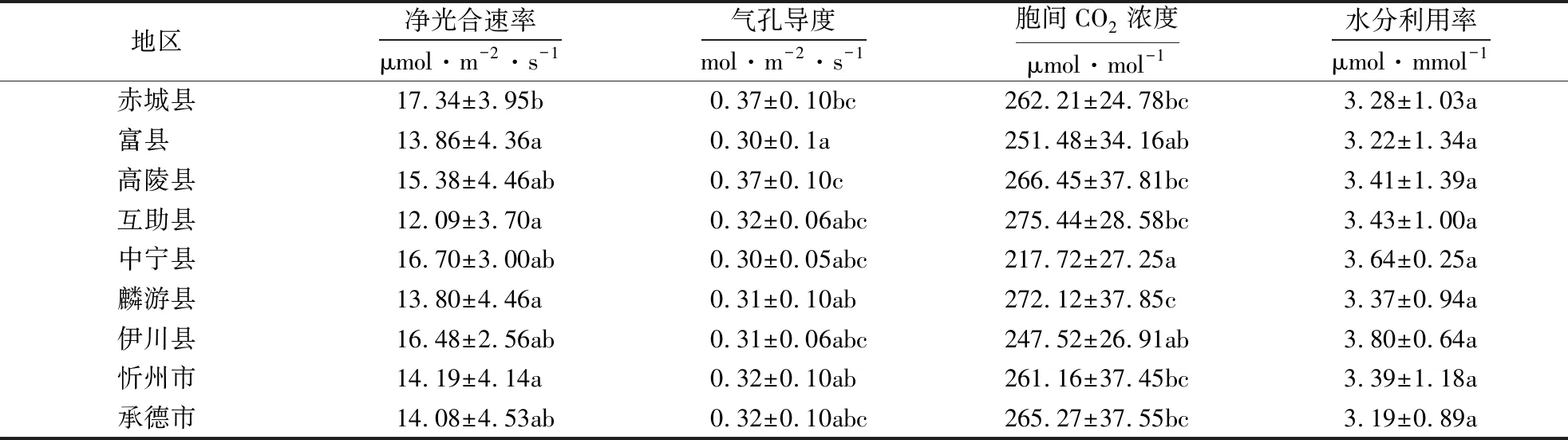
表4 小叶杨光合生理特征多重比较1)Table 4 Multiple comparison of photosynthetic physiological characteristics of P.simonii
1)不同小写字母代表差异显著(P<0.05).

表5 小叶杨叶片表型特征多重比较1)Table 5 Multiple comparison of leaf phenotypic traits of P.simonii
1)不同小写字母代表差异显著(P<0.05).
2.3 相关性分析
相关系数反映了变量之间线性关系的强弱,而正负则代表两个变量之间存在正相关或者负相关.表6为小叶杨群落的10个光合生理及叶片表型特征的Person相关系数.其中,光合生理特征之间在P=0.01水平上均显著相关,且净光合速率与气孔导度及水分利用率之间存在正强相关(0.6 表6 小叶杨光合生理特征与叶片表型特征相关系数1)Table 6 Correlation coefficients between photosynthetic physiology characteristics and leaf phenotypic traits of P.simonii 1)**代表在0.01水平上显著,*代表在0.05水平上显著. 主成分分析可用于缩减变量复杂关系,简化系统内核.通过对小叶杨光合生理及叶片表型特征进行主成分分析(表7),结果表明,前4个主成分可以解释小叶杨叶片总变异的86%,即1个四因素的模型可解释小叶杨叶片86%的变异.其中,第一、第二、第三和第四主成分的方差贡献率分别为0.29、0.26、0.18和0.13.且相关性分析表明,第一主成分与叶面积、叶长、叶宽的相关系数高达-0.96、-0.95和-0.92.说明叶片变异主要涉及叶面积和叶片长宽的变化,这与相关性分析结果是一致的. 表7 主成分贡献率Table 7 Principal component contribution rates 对整体数据进行标准化后并对各地区求平均,通过使用R语言mvstats包中的H.clust函数,根据欧氏距离对各地区小叶杨数据进行ward法系统聚类,结果如图1所示.可以清晰看出,来自不同地区小叶杨样本可按地区划分为3个子类,第1类来自富县和伊川县,第2类来自互助县、麟游县和忻州市,第3类来自于中宁县、承德市、赤城县和高陵县.从表型性状来看,富县和伊川县地区小叶杨具有最大的叶片面积、叶长度和宽度,而另外两类则相对较小,这与方差分析和多重比较结果是一致的;从地区分布来看,第1类和第3类(除中宁县外)类内地区相距较近,而第2类小叶杨样本虽然地区相差较远,但均分布多山脉地区,因此而聚为一类.值得注意的是,富县与高陵县虽地区相近,但小叶杨表型差异较大,样本也因此分布于不同子类,说明了这两者地区之间的小叶杨存在明显的变异情况. 得力于优良的环境适应性,小叶杨在我国众多地区均有分布,且由于不同地理及环境条件的影响,变种资源异常丰富.卫尊征等[16]分析了中国东北及华北地区小叶杨5个种源的17个生理及表型特征,结果表明:(1)在小叶杨的17个生理及表型特征中,有15个特征在种源内达到了差异极显著,其中,华北地区小叶杨叶片要明显大于东北地区,且山西宁武地区的小叶杨种源变异要大于吉林通榆地区;(2)小叶杨生理特征在种源间也存在显著差异,类胡萝卜素等特征的差异极有可能是由海拔因素而引起的;(3)相对于生理性状,小叶杨的表型特征稳定性较差,变异更加丰富;(4)小叶杨种源间的变异要稍低于种源内的变异,证明了种源内变异为主要变异,并且相关分析表明,生理及表型性状的变异与地理及环境因素显著相关.本文研究中,通过对比地区间小叶杨光合生理特征与叶片表型特征的平均变异系数,发现光合生理特征的平均变异系数为0.235,明显低于叶片表型特征的平均变异系数0.395,且光合生理特征的平均变异幅度0.18也明显低于表型特征的平均变异幅度0.45,证明了小叶杨生理特征的变异要低于表型特征的变异,与上述研究是一致的. 多重比较结果显示小叶杨表型特征在不同地区之间存在显著差异.由于群体间差异反映了小叶杨对不同地理情况的适应性,并且作为种内多样性的组成部分,群体间的变异也彰显了小叶杨对不同环境的广泛适应能力[21].小叶杨作为我国重要的乡土树种资源,遍布我国北部及西南地区,伴随各地区生态因素的不同,小叶杨在叶片表型特征方面也因环境而差异显著.例如,中宁县与高陵县属西北多山脉地区,气候偏干旱,叶片面积偏小以降低蒸腾速率减少水分流失[10,22];互助县地区由于海拔较高温度偏低,叶片长宽比高于其它地区,叶片偏细长且周长较大.聚类分析显示,河北承德和河北赤城归为一类,由于该两地区地理位置相距较近且小叶杨变种多为辽东小叶杨[23-24],叶片偏小并具有相似的光合及表型特性,与分类结果相符合;陕西富县和河南伊川地区同属黄土高原,生态条件差别不大且小叶杨变种多为秦岭小叶杨,因此归于一类;此外,值得注意的是,陕西富县、陕西高陵与陕西麟游虽属同一省份,但由于该地区属于小叶杨适生区[22],变种资源丰富,3个地区由于小叶杨样本的光合及表型差异而归于不同分类中.由上可见,正是由于小叶杨存在丰富的变异性,才使其能够在多个地区进行生存.同时,丰富的变异为小叶杨种质资源多样性提供了保障,也为小叶杨基因资源的研究提供了新鲜材料.加强遗传多样性研究将为小叶杨生长发育和优良树种选育提供坚实的理论基础.
2.4 主成分分析

2.5 聚类分析
3 讨论
猜你喜欢
中华医学图书情报杂志(2022年1期)2022-11-18
农学学报(2022年8期)2022-11-16
中国现代医生(2022年21期)2022-08-22
中国药学药品知识仓库(2022年9期)2022-05-23
热带农业工程(2021年5期)2021-12-06
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
天津医科大学学报(2021年1期)2021-01-26
三农资讯半月报(2020年2期)2020-03-09
江苏农业学报(2019年1期)2019-09-10
Coco薇(2017年5期)2017-06-05
