
水分胁迫对不同抗旱性燕麦品种光合及产量形成的影响
2020-03-05 03:55赵宝平徐忠山武俊英刘瑞芳刘景辉
麦类作物学报 2020年11期
赵宝平,任 鹏,徐忠山,武俊英,刘瑞芳,刘景辉
(1.内蒙古农业大学,内蒙古杂粮工程技术研究中心,内蒙古呼和浩特 010019;2.内蒙古呼和浩特园艺科技试验中心,内蒙古呼和浩特 010070)
燕麦(AvenasativaL.)属于粮饲兼用作物,同时具有降脂控糖、降低心血管疾病等功能,因而燕麦原料市场需求量较大,而且不断增长[1-3]。我国燕麦主要分布在西北等干旱半干旱地区。这些地区年降水量约在200~400 mm,农业种植以旱作为主,生育期间的干旱是限制燕麦等作物生长及高产的主要因素[4]。不同抗旱性燕麦品种的产量潜力存在较大差异[5]。因此,在水分胁迫下探究不同抗旱性品种的生理差异,对燕麦抗旱育种和栽培具有重要意义。
作物的光合作用是产量形成的物质基础,较高的光合同化能力有助于作物高产[6]。水分亏缺会导致作物叶片光合能力下降,影响产量[7]。拔节期到开花后10 d被认为是影响燕麦产量的临界期[8]。水分胁迫下燕麦叶片气孔关闭,导致净光合速率和蒸腾速率下降,提高瞬时水分利用效率[9]。水分胁迫下不同裸燕麦品种的气孔导度与光合速率均显著下降,但高产品种的光合能力降低与气孔关闭无关[10]。随着水分胁迫程度的增大,燕麦叶片气孔逐步关闭,叶绿体等超微结构受损伤程度加剧[11]。水分胁迫还会引发植物光系统Ⅱ(PSⅡ)的电子传递速率降低以及PSⅡ结构或光捕获体损伤[12]。但Ghotbi-Ravandi等研究认为,轻度水分胁迫下气孔导度是限制强抗旱性大麦品种光合的主要因素,而在重度水分胁迫下,除了气孔限制,PSⅡ结构破坏、电子传递速率减低等均影响了光合作用[13]。此外,水分胁迫下不同小麦品种的光合气体交换、叶绿素荧光参数[14]、叶绿素含量以及产量[15]均存在显著差异,说明作物光合作用过程研究不仅要考虑叶片气体交换和气孔特征,还要考察PSⅡ的结构和功能响应特征,从而揭示水分胁迫下作物光合调控机制并以此改善作物光合产量潜力[14]。叶绿素荧光技术由于可探究叶绿素蛋白体色素、激发能传递以及PSⅡ光量子传递效率等有用信息,近年来被广泛用于作物光合作用性能研究中[16]。在燕麦上,Marcińska等利用叶片气体交换和叶绿素荧光动力学参数等指标分析了不同燕麦品系的抗旱性差异[17],而将这两类指标结合分析不同水分胁迫程度下不同抗旱性燕麦品种光合和产量形成的生理差异的研究尚未见报道。为此,本试验研究不同水分胁迫下两个抗旱性不同的燕麦品种叶片叶绿素含量、光合特性、叶绿素荧光特性、产量及其构成因素,以期为燕麦节水高产栽培与抗旱育种提供理论依据与技术支持。
1 材料与方法
1.1 试验设计
1.1.1 供试材料
供试品种为河北省张家口农业科学院选育的水分敏感裸燕麦品种坝莜3号和内蒙古自治区农牧业科学院选育的抗旱皮燕麦品种蒙燕1号。
1.1.2 试验设计
试验在呼和浩特市科技园艺示范中心防雨棚内采用盆栽方式进行。设置2个品种和4个水分胁迫梯度,共8个处理,每个处理重复6次(3盆取样测定光合色素指标,3盆用来测产考种),在拔节和抽穗两个时期胁迫,共96盆。完全随机排布,并每周调换一次盆的位置。4个水分梯度分别为重度胁迫[30%田间持水量(FWC)]、中度胁迫(45%FWC)、适度控水(60%FWC)和正常供水(CK,75%FWC);待各处理达到所设置的水分梯度后,分别在拔节期(出苗后41 d)、抽穗期(出苗后62 d)开始,每天采用称重差值法实施7 d控制水分处理。处理结束之后恢复正常供水(≥75%FWC)。
盆栽土壤为沙壤土,田间持水量为16%。燕麦种植于高28 cm、直径22 cm的塑料盆,每盆装取自大田耕层土壤10 kg(干土重),并底施磷酸二铵和尿素各3 g,生育期不追肥。播种前每盆灌底墒水1.5 L,每盆播种50粒,三叶期每盆定苗25株。水分处理开始前和结束后维持正常供水直至成熟。
1.2 测定指标与方法
1.2.1 光合色素含量测定
用95%乙醇提取叶片光合色素,用分光光度计测定提取液在波长665、649和470 nm下吸光值,计算出叶绿素a、叶绿素b的含量及叶绿素a与叶绿素b的比值[11]。
1.2.2 叶片光合指标测定
采用Li-6400光合仪分别在燕麦拔节期、抽穗期胁迫结束后第一天测定最上部完全展开叶片的净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr),并计算水分利用效率(WUE=Pn/Tr)。测定时间为上午9:00-11:00,每处理测定5株,取平均值为该处理测定值。
1.2.3 叶绿素荧光动力学参数测定
用英国汉莎公司FMS-2叶绿素荧光仪在两个生育时期胁迫处理完成后当天进行叶绿素荧光动力学参数测定。选取植株上部受光一致的功能叶3片,叶片暗适应25 min后,测定初始荧光(Fo);随后强闪光(6 000 μmol·m-2·s-1),脉冲时间(0.8 s)测定最大荧光(Fm);测定充分暗适应的PSⅡ最大光化学效率(Fv/Fm)。接着照射饱和脉冲光(6 000 μmol·m-2·s-1),测定光下最大荧光(Fm′)、Ft和PSⅡ实际光化学效率(ΦPSⅡ)。根据各处理数据,计算光化学猝灭系数(qP)和非光化学猝灭系数(NPQ)。
qP=(Fm′-Ft)/(Fm′-Fo)
NPQ=(Fm-Fm′)/(Fm-Fo)
1.2.4 产量及产量构成因素测定
在燕麦成熟期,收获各处理盆中全部25株燕麦植株及籽粒,测定穗长、每穗小穗数、穗粒数(实粒数、每穗秕粒数)等,脱粒后测定穗粒重、千粒重、每盆产量等指标。
1.3 数据处理
试验数据采用SPSS 18.0及Excel 2010进行统计分析。
2 结果与分析
2.1 水分胁迫对燕麦叶片叶绿素含量的影响
在拔节期与抽穗期,正常供水(CK,75%FWC)和适度控水(60% FWC)处理下两个燕麦品种叶片叶绿素a(Chla)与叶绿素b(Chlb)含量均高于重度胁迫(30% FWC)处理(抽穗期蒙燕1号叶片Chla除外)(表1)。燕麦叶片叶绿素含量(Chla+b)随水分胁迫强度的增加而逐渐降低,且重度水分胁迫处理与CK差异显著。在拔节期,与CK相比,坝莜3号的叶片Chla+b值在重度胁迫、中度胁迫(45%FWC)、适度控水处理下分别降低了48.0%、27.6%和6.7%,蒙燕1号分别降低32.2%、21.0%和13.4%。此外,拔节期两个品种Chla/b值随着水分胁迫程度的增加均呈先增后降趋势,两个品种的Chla/b值在适度控水处理下均较高;抽穗期两个品种Chla/b值随水分胁迫强度的增加而增加,坝莜3号在不同水分处理间差异不显著,蒙燕1号的重度胁迫、中度胁迫和适度控水处理较CK分别增加了28.4%、64.8%、237.5%,差异均显著,且在重度胁迫下蒙燕1号的Chl a/b显著高于坝莜3号。
2.2 水分胁迫对燕麦叶片光合特性影响
两个燕麦品种的净光合速率(Pn)对水分胁迫的响应表现相似,拔节期和抽穗期均随水分胁迫程度的加剧而呈下降趋势。相同水分胁迫下,两个品种Pn除拔节期适度控水处理下蒙燕1号显著高于坝莜3号外,其他处理下二者差异均不显著。拔节期重度水分胁迫下坝莜3号和蒙燕1号的Pn分别较CK下降42.7%和34.2%;抽穗期分别下降27.8%和29.6%,拔节期水分胁迫对坝莜3号Pn的影响大于抽穗期(图1A)。蒸腾速率(Tr)变化趋势与Pn相似,随着水分胁迫程度的增加呈下降趋势。重度和中度胁迫下两个品种的Tr与CK差异均显著。拔节期水分胁迫对坝莜3号的Tr影响较大,而抽穗期水分胁迫对蒙燕1号影响较大(图1B)。
拔节期两个燕麦品种的气孔导度(Gs)均随着胁迫程度的加剧呈递减趋势。重度胁迫下,坝莜3号与蒙燕1号的Gs分别比CK降低55.1%和49.7%,差异均显著(P<0.05)。抽穗期水分胁迫对Gs影响不显著。两个品种Gs随着胁迫程度的增加均表现为先增后降,坝莜3号在适度控水条件下最高,蒙燕1号在中度胁迫下最高(图1C)。抽穗期水分胁迫对两个品种Ci影响均不显著,拔节期水分胁迫下蒙燕1号Ci显著低于CK,而坝莜3号的Ci值随着胁迫程度加剧呈不断增加趋势(图1D)。
随着水分胁迫程度的增加,拔节期与抽穗期蒙燕1号水分利用效率(WUE)均呈现先增后降趋势,适度控水处理下达到峰值,且高于坝莜3号。适度控水处理下蒙燕1号的WUE在拔节期与抽穗期分别较重度胁迫处理提高39.6%和 15.7%。在中度水分胁迫与适度控水处理下拔节期坝莜1号的WUE高于CK,重度水分胁迫下则表现相反。抽穗期坝莜3号的WUE在各水分胁迫处理下均低于CK,其中重度水分胁迫处理与CK差异达到显著水平(图1E)。
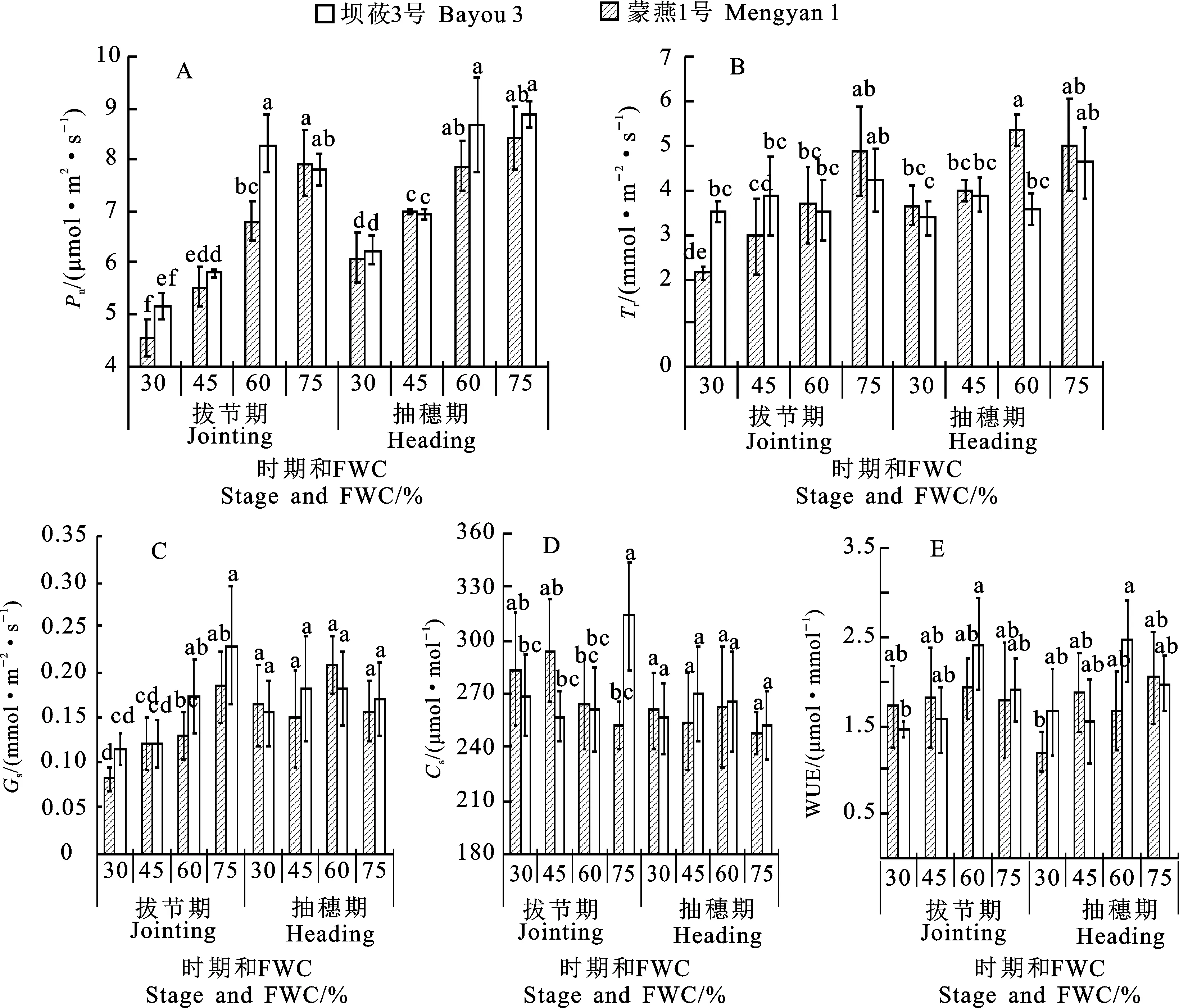
同一时期图柱上不同字母表示不同处理间在0.05水平上差异显著。图2同。
2.3 水分胁迫对燕麦叶片叶绿素荧光参数的 影响
2.3.1 初始荧光(Fo)、最大荧光(Fm)及最大光化学效率(Fv/Fm)
不同生育时期,两个燕麦品种叶片的Fo值均随着水分胁迫程度的增加呈上升趋势。重度胁迫下,坝莜3号和蒙燕1号的Fo值在拔节期较CK分别上升34.8%和8.2%,均显著高于其他处理;在抽穗期分别上升39.0%和20.3%(图2A)。Fm的变化与Fo的表现相反(图2B),两个燕麦品种的Fm均随着水分胁迫程度的加剧而下降。在CK和适度控水下,坝莜3号的Fm高于蒙燕1号;在重度和中度水分胁迫下,坝莜3号的Fm急剧下降,且在重度水分胁迫下显著低于蒙燕1号。
两个生育时期两个品种的Fv/Fm值均随水分胁迫程度增加均呈下降趋势(图2C)。与CK相比,适度控水处理的降幅较小,中度和重度胁迫处理的降幅较大,且各水分胁迫处理下抽穗期的Fv/Fm值均低于拔节期;在重度和中度水分胁迫处理下两个品种Fv/Fm值的降幅不同,蒙燕1号在拔节期和抽穗期重度水分胁迫下Fv/Fm值分别较CK下降4.8%和6.0%,而坝莜3号分别下降11.9%和14.5%;蒙燕1号的Fv/Fm值在两个时期均显著高于坝莜3号。
2.3.2 实际光化学效率(ΦPSⅡ)和电子传递速率(ETR)
不同生育时期燕麦叶片实ΦPSⅡ和ETR值均随水分胁迫程度的增加而下降。拔节期和抽穗期水分胁迫下,两个燕麦品种叶片的ΦPSⅡ均显著低于CK(图2D);重度水分胁迫下坝莜3号和蒙燕1号在拔节期、抽穗期分别较CK降低63.2%、65.0%和34.8%、29.2%,且相同水分条件下蒙燕1号的ΦPSⅡ均显著高于坝莜3号。两个品种的ETR随着水分胁迫程度的加剧均呈下降趋势,且坝莜3号的降幅大于蒙燕1号(图2E)。在拔节期和抽穗期重度水分胁迫下,蒙燕1号的ETR分别较坝莜3号高108.0%和122.8%。
2.3.3 光化学淬灭系数(qP)和非光化学淬灭系数(NPQ)
两个生育时期的水分胁迫均导致燕麦叶片qP下降,且降幅随着水分胁迫程度的增加而增大(图2F)。在抽穗期随着水分胁迫程度的加剧,两个品种的qP差异逐渐增大,在重度水分胁迫下蒙燕1号较坝莜3号高25.0%。而NPQ表现出与qP相反的趋势,其值随着水分胁迫加剧而逐渐增加(图2G),且在各水分胁迫处理下坝莜3号均高于蒙燕1号。
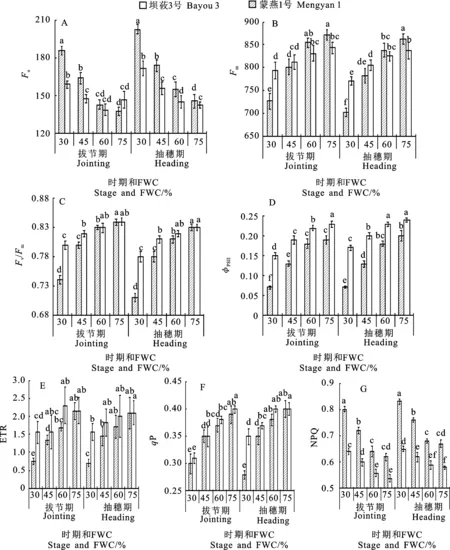
图2 不同水分胁迫下燕麦叶片叶绿素荧光参数
2.4 水分胁迫对燕麦产量及其构成因素的影响
两个生育时期的水分胁迫均对燕麦籽粒产量及其构成因素产生显著影响(表2)。随水分胁迫程度的增加,坝莜3号和蒙燕1号的穗长、单株小穗数均显著下降。拔节期重度水分胁迫下坝莜3号、蒙燕1号穗长分别较CK(75%FWC)减少26.3%、21.8%;抽穗期重度和中度水分胁迫下,坝莜3号穗长分别较CK下降25.3%和21.4%,而蒙燕1号分别下降20.3%和8.0%。水分胁迫对坝莜3号单穗小穗数的影响大于蒙燕1号。拔节期适度控水、中度和重度水分胁迫下坝莜3号单穗小穗数较CK分别下降26.4%、30.7%和 39.0%;而蒙燕1号分别下降10.0%、21.1%和 20.5%;抽穗期适度控水、中度和重度水分胁迫下坝莜3号较CK分别下降20.8%、32.9%和 41.7%;而蒙燕1号分别下降5.3%、17.2%和 29.0%。两个时期的水分胁迫对各品种的穗粒数影响均显著(表2)。拔节期各水分胁迫处理下蒙燕1号的穗粒数均显著高于坝莜3号,而在抽穗期除了重度胁迫下蒙燕1号的穗粒数较坝莜3号高32.1%之外,在其他水分处理下两个品种间差异均不显著。同一水分处理下蒙燕1号单穗实粒数显著高于坝莜3号;在抽穗期重度水分胁迫下坝莜3号和蒙燕1号的单穗实粒数分别较CK下降24.0%和26.0%,其他水分处理间差异不显著。拔节期重度水分胁迫下坝莜3号和蒙燕1号单穗秕粒数分别较CK增加134.2%和61.3%,其他处理间差异不显著;而在抽穗期坝莜3号单穗秕粒数在不同处理间差异不显著,但蒙燕1号的单穗秕粒数在重度胁迫下较CK增加72.8%。
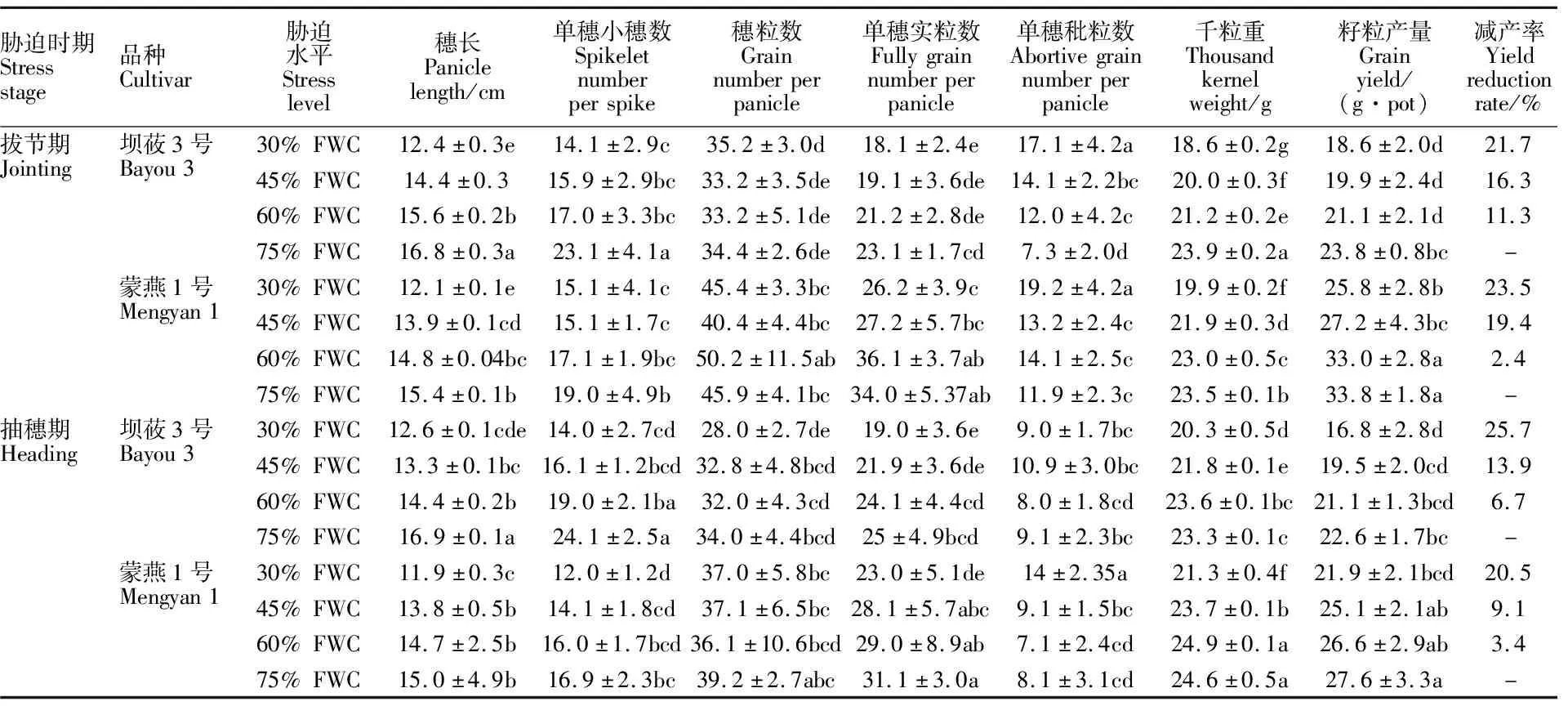
表2 水分胁迫下不同品种燕麦产量及其构成因素
两个生育时期随着水分胁迫加剧,千粒重均呈不断下降趋势。拔节期坝莜3号的千粒重在适度控水、中度胁迫和重度胁迫下分别下降 11.2%、16.5%和22.2%,而蒙燕1号分别下降 2.0%、6.9%和15.2%;在抽穗期,适度控水处理的千粒重与CK差异不显著,重度水分胁迫下蒙燕1号和坝莜3号的千粒重分别较CK下降 13.6%和 12.9%。两个生育时期随水分胁迫程度的加剧,两个品种的产量降幅逐渐增大,在拔节期适度控水处理下,坝莜3号和蒙燕1号分别减产 11.3%和2.4%;抽穗期坝莜3号在中度和重度水分胁迫下分别减产13.9%和25.7%;而蒙燕1号减产率分别为 9.1%和20.5%,且中度水分胁迫下产量与CK差异不显著。
3 讨 论
本研究表明,不同生育时期、不同水分胁迫程度下两个燕麦品种光合特性、叶绿素荧光参数和产量形成的响应均存在差异。叶绿素含量的高低在一定程度上能反映叶片的光合能力[13]。在本试验中,中度和重度水分胁迫下,两个品种叶片Chla、Chlb、Chla+b值均显著低于CK,且水分胁迫对水分敏感型品种坝莜3号影响大于抗旱品种蒙燕1号,表明干旱条件下抗旱燕麦品种的叶片光合色素具有较强保持能力。Chla/b值反映了植物对干旱胁迫的敏感性及抗旱性[18]。研究表明,干旱胁迫下金花茶幼苗叶片Chla/b值显著降低[19]。本研究中,在拔节期水分胁迫下两个品种的Chla/b值均降低,说明此期水分胁迫燕麦通过减少捕光蛋白色素复合物含量,保护PSⅡ以适应水分胁迫[20];在抽穗期重度水分胁迫下蒙燕1号Chla/b值显著高于坝莜3号,说明抗旱性强的品种可能具有更小的捕光蛋白色素复合体,以减轻水分胁迫对PSⅡ的损伤[21]。
影响植物光合作用的因素可分为气孔因素和非气孔因素[22]。本试验中,两个时期不同水分胁迫处理下燕麦叶片Pn均低于CK;蒙燕1号拔节期各水分胁迫处理的Ci均低于CK,而抽穗期均高于CK,表明拔节期蒙燕1号Pn的下降是由气孔因素引起的,而抽穗期可能是由非气孔因素引起的;而坝莜3号两个生育时期的Pn在水分胁迫下均下降,Ci均高于CK,说明该品种两个时期水分胁迫Pn的下降可能受到非气孔因素影响;同时,水分胁迫下PSⅡ实际光化学效率下降,说明其光合羧化效率降低[23]。本研究发现,适度控水下抗旱性强的蒙燕1号具有较高的叶片水分利用效率(WUE),这与Yang得出适度干旱或干湿交替灌溉有利于提高作物WUE的研究结果相似[24]。
本研究发现,不同水分胁迫下两个品种叶片叶绿素荧光动力学参数表现不同。在中度与重度水分胁迫下,坝莜3号叶片Fo显著高于蒙燕1号,而Fm和Fv/Fm均显著低于蒙燕1号,Fo的增加说明水分敏感型燕麦品种在水分胁迫下 PSⅡ 反应中心受破坏程度和光化学反应受抑制程度更大[9],导致植株光合速率降低(图1A);Fv/Fm的降低说明光捕获体向PSⅡ的电子传递能力下降,可能与上述水分胁迫下Chla/b值降低、捕光蛋白色素复合物含量下降有关;而蒙燕1号在胁迫下Fv/Fm和Chla/b值更高,说明其抗旱性主要体现在光捕获蛋白含量较高和PSⅡ活性较强[18,25]。同时,水分胁迫下ETR下降,导致激发能过量,使得光抑制作用加强[26]。此外,抽穗期各水分胁迫处理下Fv/Fm均低于拔节期,表明水分胁迫下抽穗期叶片PSⅡ活性中心受损程度大于拔节期。
水分胁迫下坝莜3号叶片实际光化学效率ΦPSⅡ显著低于蒙燕1号,表明水分胁迫导致水分敏感性品种ΦPSⅡ下降幅度更大[27]。光化学淬灭系数qP反映了PSⅡ的电子传递活性,qP的减少意味着光抑制作用增加,因此逆境条件下作物维持较高的qP值以避免光抑制的作用至关重要[14]。本试验试中两个燕麦品种随着水分胁迫程度的加剧,叶片qP逐渐降低,但蒙燕1号的qP在重度水分胁迫下显著高于坝莜3号,说明抗旱品种能够在水分胁迫下减轻光抑制作用。非光化学猝灭系数NQP反映了叶片吸收的光能以热耗散的形式散失的程度。本研究发现,水分胁迫加剧使得NQP值逐渐增加,而且坝莜3号显著高于蒙燕1号,可能是由于水分敏感型品种吸收的光能有很大比例没有被光合利用,而以热能形式(NPQ)散失[28]。
作物产量损失程度受水分胁迫时期、程度影响很大[29-30],并且不同基因型品种对水分胁迫的生理响应不同[31]。本试验中,拔节期适度控水条件下坝莜3号减产达11.3%,在抽穗期中度和重度水分胁迫下坝莜3号减产13.9%~25.7%。说明坝莜3号在拔节期对水分更加敏感。拔节期正值燕麦幼穗分化期,水分胁迫影响小穗分化、小花发育,进而导致小穗数减少、秕粒数增加甚至籽粒败育[32];水分胁迫下坝莜3号在该生育时期Pn及ΦPSⅡ、Fv/Fm均显著下降,说明水分胁迫主要通过非气孔因素限制叶片净光合速率,抑制PSⅡ活性,光合产物减少导致小穗数、粒重下降,最终影响产量。而蒙燕1号在两个生育时期适度控水处理下减产2.4%~6.7%,说明抗旱强蒙燕1号可适应适度干旱而不影响产量,在中度或重度水分胁迫下减产9.1%~23.5%,减产率低于坝莜3号(表2),在抽穗期中度胁迫下减产9.1%。这可能与其具有较高的Fv/Fm和Chla/b值,减轻了水分胁迫对光系统Ⅱ的损伤和光抑制作用,提高电子传递能力,进而提高其光合羧化效率[13],说明其在水分胁迫下光抑制作用较轻,可为穗部提供光合同化物,以保证小穗小花发育和结实,进而减少产量损失[33]。
4 结 论
两个生育时期水分胁迫均显著影响了燕麦叶绿素含量、光合速率以及荧光动力学参数,并且两个品种间存在显著差异。水分敏感品种坝莜3号叶片Pn及ΦPSⅡ、Fv/Fm受水分胁迫抑制作用显著大于抗旱品种蒙燕1号,导致穗长、小穗数、千粒重显著减少,显著增加秕粒数,减产幅度较大,甚至在拔节期适度控水即显著减产。抗旱品种蒙燕1号Fv/Fm和Chla/b值在水分胁迫下相对较高,减轻了水分胁迫对PSⅡ的损伤和光抑制作用,在适度控水下产量下降不显著,中度和重度胁迫减产幅度小于坝莜3号。这说明抗旱性品种通过维持较高的光化学效率,以促进穗粒发育,进而减少产量损失。不同生育时期水分胁迫对两个品种燕麦光合及产量影响效果不同,整体来看拔节期受水分胁迫影响程度更大。
猜你喜欢
中老年保健(2022年5期)2022-11-25
核农学报(2022年11期)2022-10-29
成都信息工程大学学报(2022年2期)2022-06-14
今日农业(2021年6期)2021-11-27
文萃报·周五版(2021年37期)2021-10-09
今日农业(2021年7期)2021-07-28
电脑报(2020年12期)2020-06-30
农民致富之友(2017年15期)2017-08-20
延河(2017年7期)2017-07-19
农产品市场周刊(2017年17期)2017-06-02
