
芝麻种子萌发动态及其代谢生理变化研究
2020-03-15 03:26孙建周红英乐美旺颜廷献饶月亮颜小文梁俊超叶艳英
中国农业科技导报 2020年8期
孙建, 周红英, 乐美旺, 颜廷献, 饶月亮, 颜小文, 梁俊超, 叶艳英
(江西省农业科学院作物研究所,江西省油料作物生物学重点实验室,国家油料改良中心南昌分中心,农业农村部长江中下游作物生理生态与耕作重点实验室,南昌 330200)
芝麻(SesamumindicumL.)是重要优质油料作物,种子含油量高,不饱和脂肪酸占85%左右,富含芝麻素(sesamin)、芝麻林素(sesamolin)、维生素E等天然抗氧化类物质[1-3],是重要的优质食用油源和营养保健食品[4-5]。萌发生理是种子生物学的重要内容之一,在很多植物中开展了大量相关研究,包括种子萌发的逆境生理、营养及化学成分变化和代谢生理等方面。关于芝麻种子萌发过程的相关研究多集中在营养及化学成分[6-7]、芽苗生长与生理[8-9]等方面,关于代谢生理方面的报道不多,韩亚飞等[10]在30 ℃条件下对芝麻种子萌发0~48 h的总糖、粗脂肪、粗蛋白和氨基酸组成等变化进行研究,结果显示,总糖含量先降后升,粗脂肪含量逐渐下降,粗蛋白含量变化不明显,不同类型氨基酸占比变化不同。张瑞等[11]研究了35 ℃暗培养条件下芝麻种子萌发 0~72 h 的粗脂肪、粗蛋白、总糖和脂肪酸组成等的变化,结果显示,粗脂肪含量在36 h后降低幅度较大,粗蛋白含量变化较小,总糖含量不断增加,脂肪酸组成之间变化幅度较小。张丽霞等[12]研究了芝麻种子在28 ℃条件下萌发0~96 h的脂肪酸组成、氨基酸组成、粗脂肪、粗蛋白等主要成分的动态变化,发现粗脂肪含量在萌发的36 h以前变化较小,粗蛋白含量则呈现先降后升再降的变化,84 h后下降明显,主要脂肪酸组分的变化不大,17种氨基酸变化各不相同,酸性氨基酸占比随着种子萌发不断增加,中性和碱性氨基酸不断减少。这些研究主要针对芝麻种子萌发过程的营养成分变化,尚未从代谢生理的角度对芝麻种子主要贮藏物质参与代谢的先后顺序进行研究。因此,本研究拟检测芝麻萌发动态的形态学和水分需求变化,确定芝麻种子萌发的关键时间点,并进一步测定芝麻种子萌发过程中与代谢相关的可溶性糖、可溶性蛋白、氨基酸、游离脂肪酸及脂肪酶等生理指标变化,明确种子主要贮藏物质参与代谢的先后顺序,在萌发动态和代谢生理上丰富芝麻种子生物学相关内容,并为芝麻种子萌发代谢机理的进一步研究提供参考。
1 材料与方法
1.1 试验材料
供试材料为赣芝9号和赣芝12号,其中赣芝9号为黑芝麻品种,粗脂肪含量为50.5%,粗蛋白含量为19.7%;赣芝12号为白芝麻品种,粗脂肪含量57.1%,粗蛋白含量20.6%。种子均由江西省农业科学院作物研究所芝麻创新团队提供,选取正常成熟、籽粒饱满的芝麻种子备用。
1.2 试验设计
试验分两步开展,均在黑暗状态的25 ℃恒温培养箱中进行。第一步用于观察芝麻种子萌发动态,测定吸水率、吸水速率、粗脂肪含量和脂肪酸组成,将精选的干净种子放置在铺有3层滤纸的发芽盘中,每盘称取10 g平铺于滤纸上,确保种子籽粒之间不重叠,每盘等量地加入充足的蒸馏水以供萌发,分别在0、3、6、9、12、15、18、21、24、30、36、42、48、54、60、66和72 h取样,每个时间点取样3盘,即3次重复。每个样品取出后测定鲜重,并于60 ℃烘箱烘干至恒重,称量干重用于粗脂肪、脂肪酸组成的测定。根据第一步的发芽动态、吸水率和吸水速率测定结果,确定0、6、12、24、48、72 h共6个时间点进行第二步试验,将精选的干净种子放置在铺有3层滤纸的发芽盒中,发芽盒长11 cm、宽11 cm、高6 cm,每盒放置种子200粒,同样于25 ℃暗培养,每个时间点取样3盒,即3次重复,将取样样品用蒸馏水冲洗干净并用滤纸吸干水分备用,用于测定可溶性糖含量等生理指标。
1.3 测定指标与方法
吸水率为种子吸收水分的质量占种子干重的百分比,吸水速率为每g干种子每h吸收的水分质量。粗脂肪含量采取索氏抽提法(GB/T 14772-2008)[11]测定,采取气相色谱法[11]测定棕榈酸、硬脂酸、油酸和亚油酸4个主要脂肪酸组成成分。可溶性糖含量采用蒽酮比色法[13]测定,氨基酸含量采用水合茚三酮法[13]测定。可溶性蛋白含量采用BCA法[14]测定。脂肪酶活性通过铜皂-分光光度法测定脂肪酸生成速率并进行计算[15]。游离脂肪酸含量采取铜皂比色法[16]测定。所用试剂均购自苏州科铭生物技术公司。
1.4 数据分析
Microsoft Excel 2007和SPSS 18.0软件进行数据整理、相关性分析及作图。
2 结果与分析
2.1 种子萌发动态及其水分变化
芝麻种子萌发动态见图1,可见,25 ℃暗培养情况下,12 h芝麻种子实现萌动露白,24 h进入胚根快速伸长阶段,48 h胚根和胚芽同步快速伸长生长,72 h子叶开始展开完成种子萌发过程,继而进入芽期幼苗生长阶段。
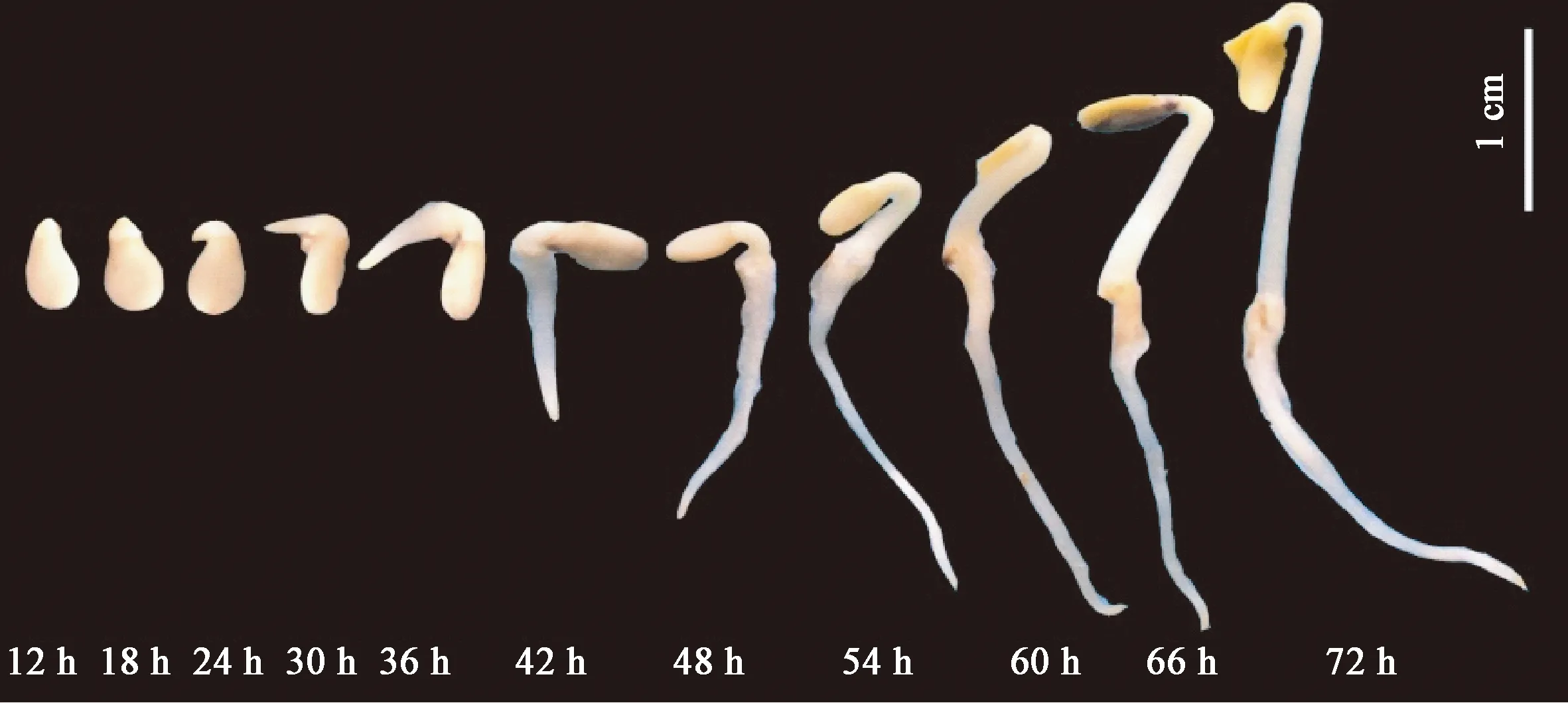
图1 芝麻种子萌发过程的形态学观察Fig.1 Germination morphology of sesame seed
芝麻种子萌发过程的吸水率和吸水速率结果(图2)显示,种子吸水率呈先急速上升后平稳增加接着再次快速上升的规律,呈现两个快速吸水期,赣芝9号和赣芝12号3 h吸水率分别达到50.77%和44.19%,并在6 h后进入较为平稳增加阶段,24~54 h,吸水率开始进入第二次快速上升阶段,赣芝9号和赣芝12号分别迅速增加到162.55%和153.92%,接着增速有所减缓。吸水速率的变化呈现出由高吸水速率(赣芝9号和赣芝12号分别为169.23和147.12 mg·h-1·g-1)快速下降,9 h降到最低,随后开始平稳上升,至 54 h 达到第二个峰值(赣芝9号和赣芝12号分别为51.51和45.79 mg·h-1·g-1),然后再次下降。两个品种萌发过程中的吸水率和吸水速率的变化趋势表现一致,但也存在较小差异,这可能与品种的种皮结构、物质组成和萌发特点等差异有关。
结合种子萌发形态和种子吸水率、吸水速率结果,可以确定25 ℃暗培养条件下,芝麻种子于浸水6 h时吸收的水分超过自身干重,基本满足对水分的需求,达到种子萌发阶段的“吸胀”,此前吸水率急剧上升,吸水速率最高。12 h进入种子萌发的第二阶段“萌动”,即“露白”,对水分的需求趋于平稳,吸水率增加较慢,吸水速率在较低水平变化。随后在24 h开始进入胚根伸长时期, 48 h后胚根和胚芽的下胚轴开始同步进入快速伸长生长时期,对水分的需求开始快速增加。72 h 时子叶基本展开,完成了种子萌发的第三阶段“发芽”,随后进入幼苗生长阶段。
鉴于此,本研究下一步开展的代谢生理研究将围绕干种子(0 h)、吸胀(6 h)、萌动(12 h)、胚根开始伸长(24 h)、胚芽的下胚轴开始伸长(48 h)和基本完成萌发(72 h)共6个时间点进行。
2.2 种子萌发过程的可溶性糖含量变化
可溶性糖是芝麻种子贮藏的主要碳水化合物之一,也是种子萌发过程中脂肪代谢的最终产物之一。图3显示,芝麻干种子中贮藏有一定量的可溶性糖,吸胀(6 h)时可溶性糖含量急剧下降,差异达显著水平。随着萌发过程的推进,可溶性糖含量呈先降后升的变化趋势,品种间变化趋势基本一致。种子吸胀(6 h)到胚芽下胚轴开始伸长生长(48 h),可溶性糖被不断消耗,含量不断降低,在完成发芽(72 h)时含量又有所增加,差异达到显著水平。
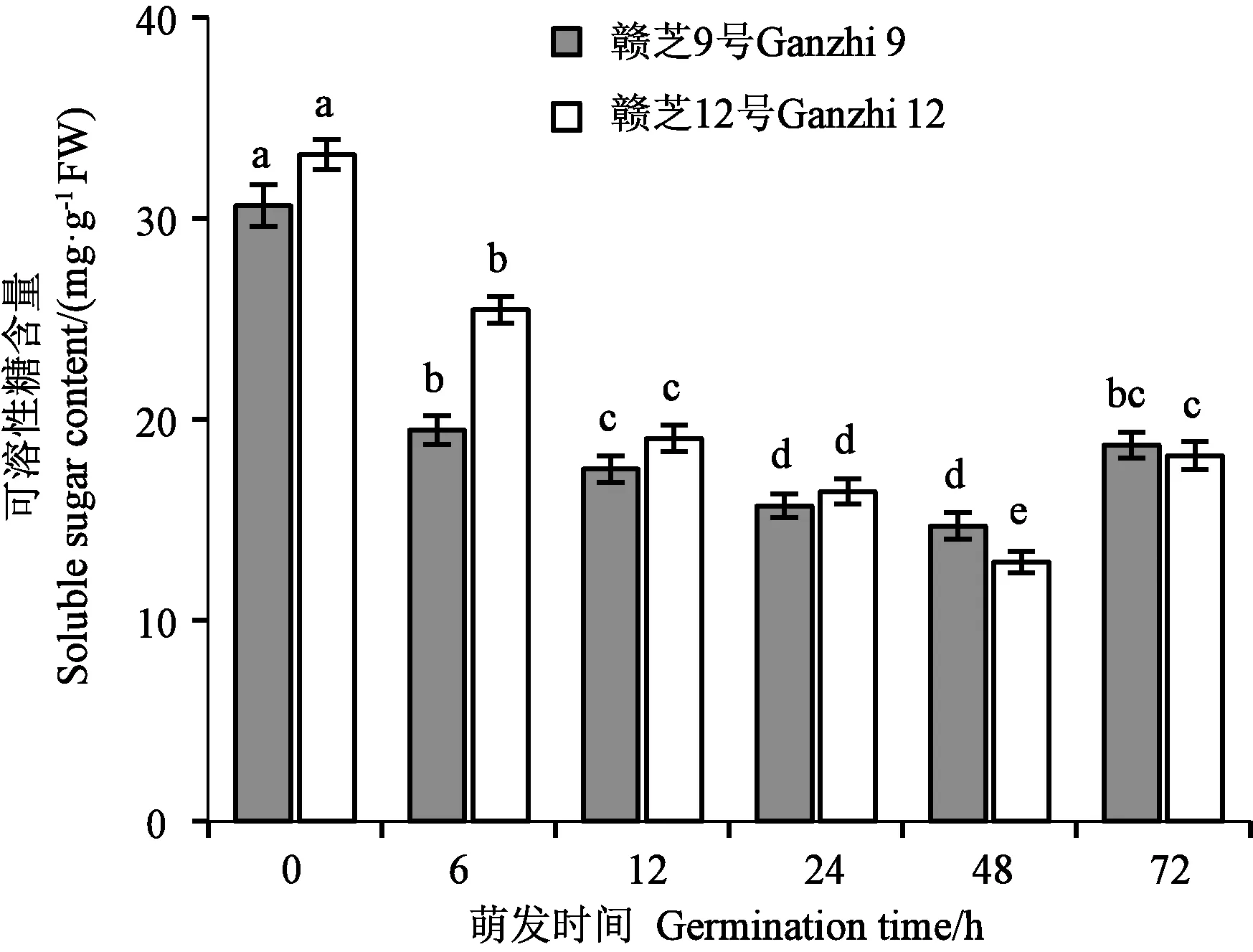
注:不同小写字母表示同一品种不同萌发时间的差异在P<0.05水平具有统计学意义。Note: Different lowercase letters of the same variety indicate significant difference between different germination times at P<0.05 level.图3 芝麻种子萌发过程的可溶性糖含量变化Fig.3 Changes of soluble sugar content during germination of sesame seed
2.3 种子萌发过程的蛋白质代谢
蛋白可溶化是贮藏蛋白进入代谢过程的第一步。2个供试品种种子萌发过程中,可溶性蛋白含量均呈先降后升的变化规律(图4),吸胀(6 h)时可溶性蛋白含量较干种子显著下降,然后随着种子的萌发进程,可溶性蛋白含量不断升高,6~72 h不同萌发时间的可溶性蛋白含量差异均达到显著水平。
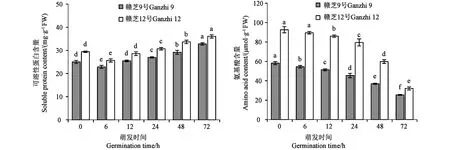
注:不同小写字母表示同一品种不同萌发时间的差异在P<0.05水平具有统计学意义。Note: Different lowercase letters of the same variety indicate significant difference between different germination times at P<0.05 level.图4 芝麻种子萌发过程中可溶性蛋白含量和氨基酸含量的变化Fig.4 Changes of soluble protein content and amino acid content during germination of sesame seed
作为蛋白质的代谢产物,氨基酸是参与生命活动的重要因子。芝麻种子萌发过程中,氨基酸含量呈现不断下降趋势(图4),同一品种不同萌发时间的氨基酸含量差异均达到显著水平。供试品种间的变化趋势基本一致,但品种间的含量存在较大差异,赣芝12号的氨基酸含量明显高于相同萌发时间的赣芝9号。
2.4 种子萌发过程的脂肪代谢
萌发过程中种子粗脂肪含量结果(图5)显示,从干种子(0 h)、吸胀(6 h)、萌动(12 h)到胚根伸长(24 h),种子的粗脂肪含量没有显著变化。胚芽伸长(48 h)后,粗脂肪含量开始显著下降。在种子萌发过程中,游离脂肪酸含量表现为先急剧下降后不断升高再快速下降的趋势。在干种子(0 h)中存在一定含量的游离脂肪酸,在种子吸胀(6 h)时游离脂肪酸急剧下降到最低水平,其后随着种子萌发进程的推进,游离脂肪酸含量不断上升,在胚芽伸长(48 h)时达到最高值,然后在发芽(72 h)时再次快速下降。脂肪酶活性结果(图5)显示,种子萌发过程中脂肪酶活性不断增加,品种间总体变化趋势基本一致,但品种间存在差异。在吸胀(6 h)和萌动(12 h)时两个供试品种间基本没有差异,胚根伸长(24 h)后,赣芝12号的脂肪酶活性大幅度增加,而赣芝9号的脂肪酶活性增加较缓,这可能与品种的粗脂肪含量有关,高含油量的赣芝12号较低含油量的赣芝9号在种子萌发的脂肪代谢中需要更高活性的脂肪酶。
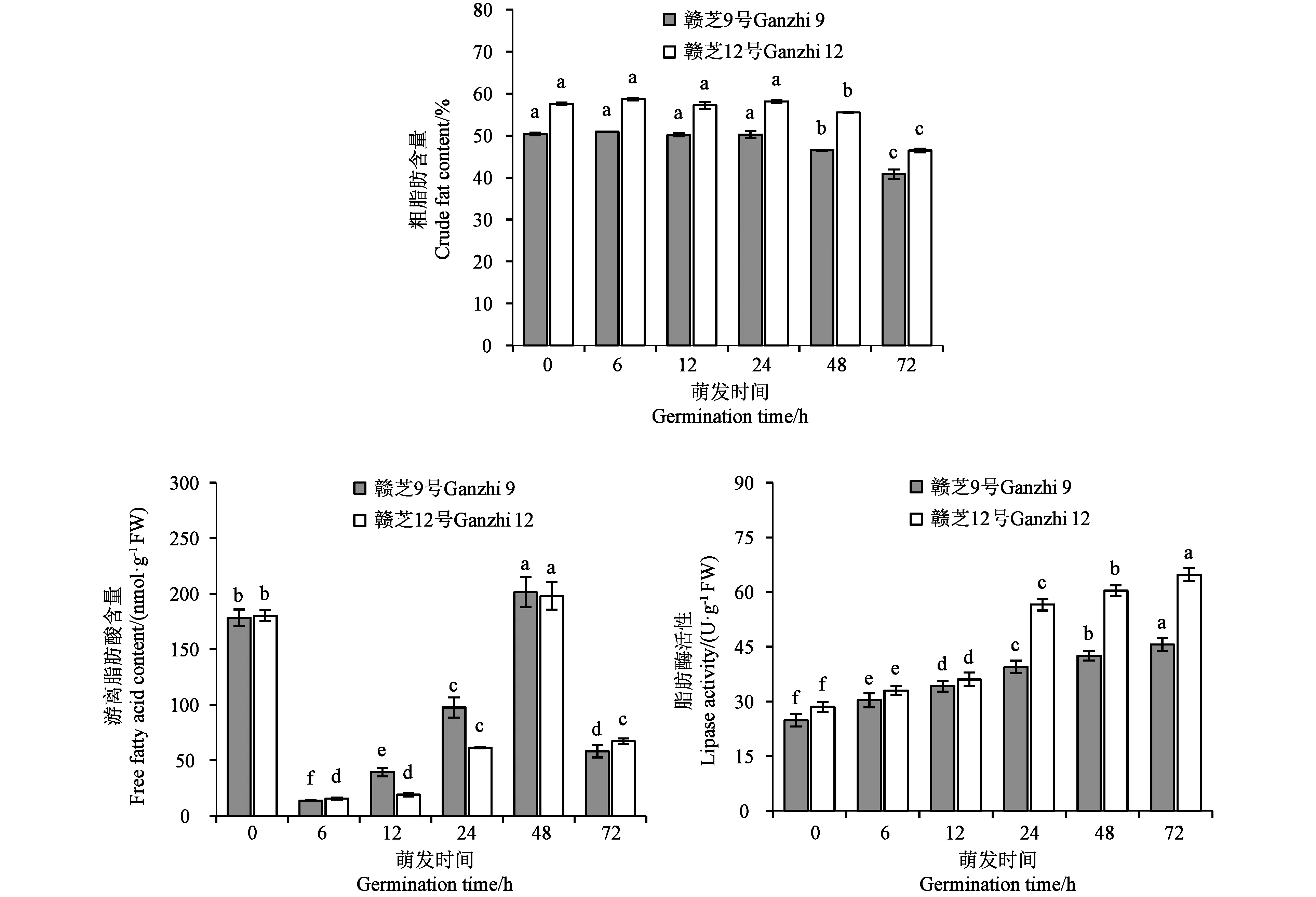
注:不同小写字母表示同一品种不同萌发时间的差异在P<0.05水平具有统计学意义。Note: Different lowercase letters of the same variety indicate significant difference between different germination times at P<0.05 level.图5 芝麻种子萌发过程中粗脂肪含量、游离脂肪酸含量和脂肪酶活性的变化Fig.5 Changes of crude fat content, free fatty acid content and lipase activity during germination of sesame
对种子萌发过程中的主要脂肪酸组成进行分析,结果(图6)显示,棕榈酸、硬脂酸、油酸和亚油酸4种主要脂肪酸组成基本保持稳定,变化较小,这与前人的研究结果[11-12]一致。棕榈酸在种子发芽(72 h)时有所降低,但降低幅度较小,分别为1.56%(赣芝9号)和1.94%(赣芝12号)。相关性分析显示,油酸与亚油酸之间存在显著负相关关系(r=-0.522,P<0.01)。
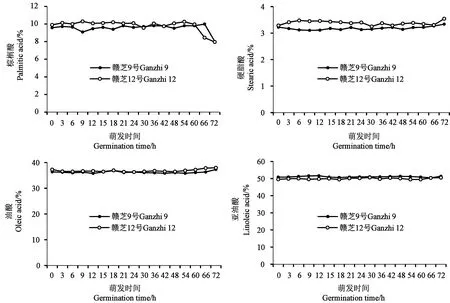
图6 芝麻种子萌发过程的主要脂肪酸组成变化Fig.6 Changes of main fatty acid composition during germination of sesame seed
2.5 相关分析
对芝麻种子萌发过程中与代谢相关的可溶性糖、可溶性蛋白、氨基酸、粗脂肪、游离脂肪酸和脂肪酶活性共6个生理指标进行相关分析,结果(表1)显示,可溶性糖含量与脂肪酶活性呈显著负相关(P<0.05),粗脂肪含量与氨基酸含量呈极显著正相关(P<0.01),脂肪酶活性与可溶性蛋白呈极显著正相关(P<0.01)。蛋白质代谢中,可溶性蛋白含量与氨基酸含量呈负相关,与蛋白质代谢生成氨基酸有关。脂肪代谢中,脂肪酶活性与游离脂肪酸含量呈正相关,与粗脂肪含量呈负相关,粗脂肪含量与游离脂肪酸含量正相关,但三者相互间的相关系数均较小,可能与芝麻种子萌发0~72 h内脂肪代谢量较低有关。
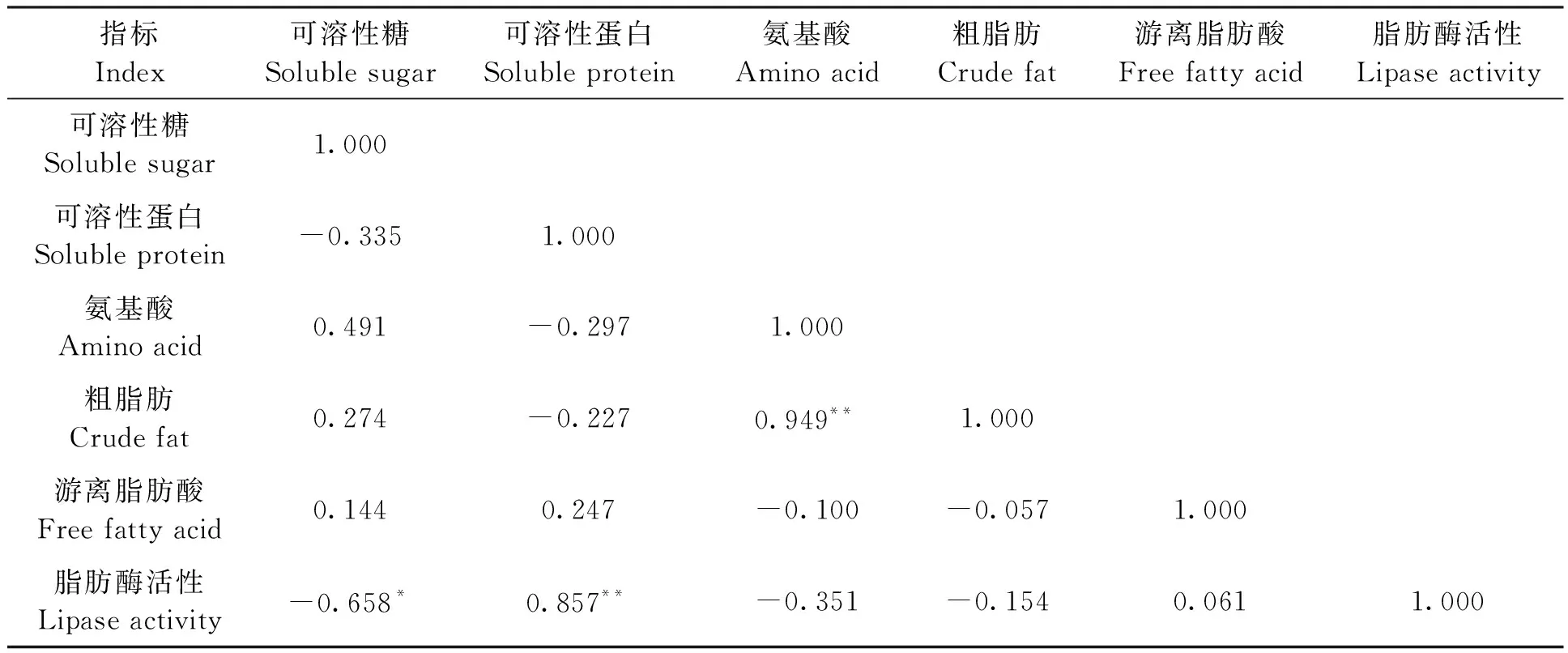
表1 芝麻种子萌发过程代谢生理指标的相关分析Table 1 Correlation analysis of physiological indexes of metabolism during germination of sesame seed
3 讨论
种子萌发是植物重要的生命过程之一,可以分为吸胀、萌动、发芽和成苗4个阶段,涉及一系列的生理和形态变化,并受环境条件的影响[17]。本研究在25 ℃暗培养条件下,通过形态学、水分吸收和代谢生理指标的检测,明确了芝麻种子吸胀、萌动、胚根快速伸长、胚芽快速生长和完成发芽的时间点,并对芝麻种子的萌发低谢机理进一步研究。芝麻种子吸胀时间的确定为芝麻生产中浸种处理提供理论参考,即25 ℃时浸种时间不宜超过6 h,温度较高时可适当缩减浸种时间[10-11]。
碳水化合物(糖)、蛋白质和脂肪是芝麻种子中的主要物质成分,占种子质量的90%以上[18],也是参与芝麻种子萌发代谢的主要物质。芝麻种子中糖类主要是葡萄糖、果糖、蔗糖、车前糖等可溶性糖[18-19]。种子萌发过程中,可溶性糖中的葡萄糖等直接被代谢利用,蔗糖等则被快速分解为小分子物质被利用,为种子萌发和胚生长提供能量[17,20]。本研究中芝麻萌发过程中,可溶性糖含量在0~48 h之间不断减少,在完成发芽的72 h时则表现为增加,这与韩亚飞等[10]的研究一致,也与其他脂肪类种子的研究结果[20-21]相同。种子萌发72 h时,可溶性糖含量增高,可能是脂肪代谢产生糖类的原因[17]。因此,糖类是芝麻种子萌发过程中最先被代谢利用的物质,为萌发和生长提供能量,而脂肪代谢明显迟于糖代谢。
蛋白质在种子萌发过程中具有不可替代的作用,其代谢产物氨基酸参与新蛋白合成或为呼吸氧化提供碳架[17,22-23]。种子中的蛋白质主要以贮藏蛋白形式存在,其参与代谢的第一步是转化为可溶性蛋白。本研究中芝麻种子萌发过程中,吸胀(6 h)时可溶性蛋白较干种子有所降低,随后随着萌发进程的推进不断上升,变化趋势与花生[24]和刺萼龙葵[25]等的研究结果相似。可能是种子吸胀引起细胞膜损伤导致可溶性蛋白透过膜外渗,从而表现为可溶性蛋白含量降低,随后贮藏蛋白不断实现水溶化,导致可溶性蛋白含量增加[20]。
作为种子中高能量贮藏物,脂肪在酶的作用下分解出的脂肪酸转化为糖供植物利用[20]。本研究结果显示,芝麻种子萌发过程中粗脂肪含量在24 h前基本没有变化,48 h后才开始不断下降。游离脂肪酸含量在吸胀(6 h)时大幅度下降,这可能与吸胀引起的细胞膜损伤导致的游离脂肪酸外渗有关[17,20],随后不断上升,于48 h达到最高值后再次下降。随后脂肪分解,游离脂肪酸含量不断上升,在48 h之后游离脂肪酸进入代谢过程,其含量再次下降。综合来看,粗脂肪含量24 h之后开始减少,游离脂肪酸含量48 h达到高峰后快速降低,脂肪酶活性不断增强,以及可溶性糖含量于72 h再次升高,可以认为芝麻种子萌发过程中脂肪的代谢发生较晚。
综上所述,在芝麻种子萌发过程中,糖类物质最先参与代谢,其后是蛋白质,最后才是脂肪,这与其他油料作物种子萌发代谢顺序基本一致[17,20]。本研究结果阐述了芝麻种子萌发过程中物质代谢先后顺序,为萌发过程中相关基因表达及代谢组学等进一步研究提供了理论参考。
猜你喜欢
中华胰腺病杂志(2022年4期)2022-08-23
现代农业科技(2022年12期)2022-07-02
农村青少年科学探究(2022年3期)2022-05-13
中老年保健(2021年3期)2021-08-22
求知导刊(2019年15期)2019-08-30
小学生优秀作文(高年级)(2018年9期)2018-09-14
广西科技大学学报(2018年2期)2018-09-10
绿色科技(2016年20期)2016-12-27
湖北农业科学(2014年21期)2014-12-22
杂文选刊(2007年13期)2007-02-11
