
紫薇金叶品种金幌叶色变化响应高光照的生理特性
2020-03-27 09:30乔东亚王鹏王淑安张恩亮高露璐李亚杨如同
江苏农业学报 2020年1期
乔东亚 王鹏 王淑安 张恩亮 高露璐 李亚 杨如同


摘要:以紫薇品种金幌和粉晶为材料,对其由低光照[50~120 μmol/(m2·s)]转向高光照[1 200~2 000 μmol/(m2·s)]后168 h内叶片叶绿素含量、光合特性、叶绿素荧光参数、叶绿素合成前体物质含量及过氧化物酶活性等进行了测定和分析。结果表明:(1)与低光照相比,高光照处理168 h后,金幌叶片中叶绿素a、叶绿素b含量、超氧化物歧化酶活性、净光合速率和蒸腾速率显著低于粉晶。(2)在低光照时,金幌叶片中的5-氨基乙酰丙酸、胆色素、尿卟啉和粪卟啉等4种前体物质含量均显著高于粉晶,其中粪卟啉含量是粉晶中的2倍。(3)高光照处理168 h后,金幌叶片中非光化学猝灭(NPQ)参数值显著高于粉晶,而PSⅡ原初光化学效率(Fv/Fm)和非循环光合电子传递速率(ETR)则与粉晶差异不显著。说明金幌叶色变化的直接原因是叶绿素含量,特别是叶绿素b含量变化。叶绿素b含量的改变则是由于叶绿素合成前体物质,特别是粪卟啉的累积造成的。叶绿素含量的改变同时导致了金幌光合能力的降低及光合系统PS II的稳定性的改变。
关键词:紫薇;叶色;光响应;叶绿体
中图分类号:S685.99文献标识码:A文章编号:1000-4440(2020)01-0180-07
Abstract:In this study, the acclimation response of crape myrtle (Lagerstroemia indica) Jinhuang and Fenjing plants was assessed during 168 h after transferring from low light [LL, 50-120 μmol/(m2·s)] to high light [HL, 1 200-2 000 μmol/(m2·s)] by analyzing the content of chlorophyll, photosynthetic properties, chlorophyll fluorescence parameters, chlorophyll synthesis precursors contents and peroxidase activity. The results showed that compared with low light treatment, the contents of chlorophyll a and chlorophyll b, superoxide dismutase activity, net photosynthetic rate and transpiration rate of leaves in Jinhuang were significantly lower than those in Fenjing after 168 hours of high light treatment. Under low light conditions, the contents of four precursors including 5-aminolevulinic acid, bile pigment, urinary porphyrin and coproporphyrin in Jinhuang leaves were significantly higher than those in Fenjing, and the content of coproporphyrin was twice as high as that in Fenjing. After 168 hours of high light treatment, the value of non-photochemical quenching (NPQ) in Jinhuang leaves was significantly higher than that in Fenjing, while the differences of initial light energy conversion efficiency of PS II(Fv/Fm) and electron transport rate(ETR) were not significant. This indicates that the change of leaf color in Jinhuang is directly caused by the change of chlorophyll content, especially the content of chlorophyll b. The change of chlorophyll b content is caused by the accumulation of chlorophyll synthesis precursors, especially coprophyrin. The changes of chlorophyll content also lead to the decrease of photosynthetic capacity and PS II stability in Jinhuang.
Key words:Lagerstroemia indica;leaf color;photoresponse;chloroplast
紫薇(Lagerstroemia indica),千屈菜科(Lythraceae)紫薇屬(Lagerstroemia)植物,具有花期长、花色丰富等特点,素有“满堂红”、“百日红”之称,为中国乃至世界上重要的夏季观花植物。紫薇金叶品种金幌是由紫薇品种粉晶的金色芽变培育的彩叶紫薇品种,与传统观花紫薇相比,其叶色金黄,独具特点,具有较高的观赏价值[1]。通过前期对叶色变异机理的研究,发现金幌叶色变异与光照度有关[2],但是关于其对高光照响应的生理机制还不清楚。
叶色突变体的表型与叶片中细胞色素的类型和各种色素的含量以及它们之间的比例有关,同时光照时间及强度等环境因素也影响其表型。不同光照度处理下,金叶与正常叶的叶绿素含量有差异[3-5]。金边大叶黄杨(Euonymus japonicus Ovatus Aureus)在低光照度条件下生长时叶片叶绿素含量显著高于全光条件下[6]。随着光照度的增加,中华金叶榆叶片叶绿素含量发生很大变化,类胡萝卜素、叶绿素及花色素苷含量间的比值最终影响中华金叶榆叶片颜色的表达。
高光照下,叶色黄化突变体很容易出现灼伤现象。玉簪的黄化品种(Hosta Golden Edger)和花叶品种(Hosta Antioch)等在光照过强时,会出现不同程度的焦边、灼伤,甚至地上部枯死等现象[7-8]。非光化学猝灭系数越高的鸡爪槭品种日灼现象越严重,实际光化学效率和电子传递速率(ETR)与日灼抗性间关系密切,易受日灼伤害的品种有较低的实际光化学效率和ETR值[9]。
本研究以紫薇金叶品种金幌和绿叶品种粉晶为材料,测定2个品种由低光照转向高光照后不同时间叶片中叶绿素含量、叶绿素合成前体物质含量、光合参数、叶绿素荧光参数及过氧化物酶活性等指标,以期解析金幌叶色变化响应高光照的生理特征。
1材料与方法
1.1试验材料
紫薇金叶品种金幌及对照品种粉晶的2年生扦插苗各40株定植于江蘇省中国科学院植物研究所紫薇种质资源圃,植株健壮,株距50 cm×50 cm,正常水肥管理。7月初,金幌及粉晶单株在光照度50~120 μmol/(m2·s)低光照(遮荫)下处理20 d,然后在高光照(1 200~2 000 μmol/(m2·s))下处理7 d。
1.2叶片色素含量测定
选取金幌和粉晶当年生枝条第3~4节部成熟叶片,混合取样,测定色素含量[9]。称取0.10 g 叶片放入10 ml 试管中,加入10 ml 95%的乙醇,封口后闭光放置48 h,直到叶片完全变为白色。然后用分光光度计测定样品OD 值,3次重复。取样时间为高光照处理第0 d、2 d、4 d和7 d。
1.3光合特性相关指标的测定
选取当年生枝条第3~4 节部成熟叶片,在外界光照度1 200~2 000 μmol/(m2·s)、叶室光照度1 200 μmol/(m2·s)、气温28 ℃条件下,使用LI-6400 便携式光合仪测定净光合速率、气孔导度、蒸腾速率等光合参数。测定时间为高光照处理第0 d、2 d、4 d和7 d的上午10∶00-11∶00,6次重复。
1.4叶片叶绿素荧光参数的测定
参考Brian等[10]的方法,利用OS-5P+便携式脉冲调制叶绿素荧光仪,在自然光照条件下测定当年生无病虫害枝条第3~4 节部成熟叶片的PSⅡ原初光化学效率(Fv/Fm)、非循环光合电子传递速率(ETR)和非光化学猝灭(NPQ)等参数。测定时间为高光照处理第0 h、2 h、4 h和7 h,6次重复。
1.5叶绿素合成前体物质含量测定
取当年生第3~4 节部成熟叶片1.00 g,去掉主脉,剪碎。 氨基乙酰丙酸(ALA)含量的测定与胆色素原(PBG)含量的测定参照Bogorad[11]的方法。参照 Bogorad [11]的方法,在Perkin Elmer Ls荧光分光光度计上测定E400F633、E400F622和E400 F640,计算原卟啉(ProtoIX)的含量;测定E420F678,计算镁原卟啉(Mg-ProtoIX)含量;测定E440F640、E400F633,计算原叶绿素酯(Pchlide)含量。样品采集时间为高光照处理第0 h、2 h、4 h和7 h ,3次重复。
1.6过氧化物酶活性测定
在高光照处理第0、2、4和7 d的上午10∶00-11∶00 时,选取当年生无病虫害枝条第3~4节部成熟叶片,混合取样,测定过氧化物酶活性。超氧化物歧化酶(SOD)活性利用总超氧化物歧化酶测定试剂盒(A001-3,南京建成生物科技有限公司产品)测定,过氧化氢酶(CAT)活性利用过氧化氢酶试剂盒(A007-1,南京建成生物科技有限公司产品)测定,过氧化物酶(POD)活性利用过氧化物酶试剂盒(A804-3,南京建成生物科技有限公司产品)测定,3次重复。
1.7数据处理
图片数据使用Adobe Photoshop CS6(Adobe Inc., California, USA)进行处理。采用Excel 2016进行数据整理、作图,用SPSS 22.0(SPSS Inc., Chicago, USA)进行方差分析,LSD法进行多重比较,所测数据以平均值±标准差(SE)表示。
2结果与分析
2.1光照度增加导致紫薇金幌叶绿素b含量的降低及类病斑的形成
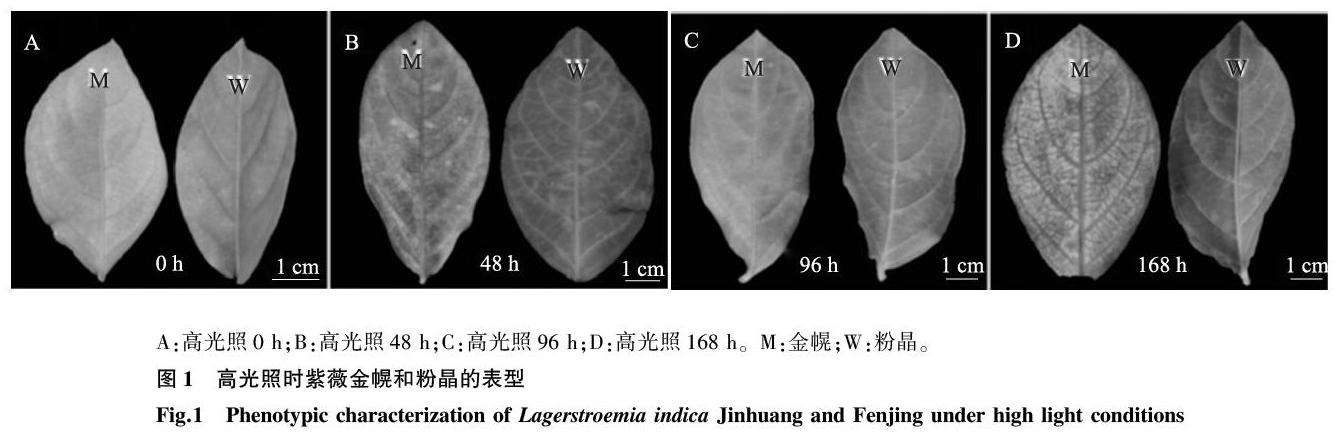
在低光照处理结束时(高光照0 h)时,金幌叶色淡黄,粉晶叶色黄绿,高光照处理168 h后,金幌叶片叶肉部分出现白化现象,黄色部分颜色变浅,粉晶恢复绿色(图1A、图1D)。为了阐明叶色变化原因,我们测定了金幌和粉晶的色素含量变化(图2)。金幌和粉晶在低光照处理20 d后,仅叶绿素a含量在两者叶片中有显著差异。高光照处理48 h后,粉晶叶片中叶绿素a、叶绿素b和类胡萝卜素含量均高于金幌,并且叶绿素a含量在两者之间有显著差异。高光照处理96 h后,金幌叶片中叶绿素a/叶绿素b显著高于粉晶,并且叶绿素a、叶绿素b含量显著低于粉晶。高光照处理168 h后,金幌叶片中叶绿素a、叶绿素b含量显著低于粉晶,类胡萝卜素含量和叶绿素a/叶绿素b均高于粉晶;粉晶叶片中叶绿素a、类胡萝卜素含量均增加,叶绿素b含量保持稳定。但金幌叶片中叶绿素a、叶绿素b含量下降幅度较大,仅类胡萝卜素含量有所增加。根据以上结果可以确定光照度引起金幌叶色变化的主要原因是叶绿素含量的变化。
叶绿素合成是从5-ALA开始的连续的生化过程,该过程大约需要15种酶参与,产生相应的中间产物,阐明中间产物的合成变化对解释叶绿素b含量的变化具有重要意义。叶绿素合成的中间产物(前体物质)含量测定结果(表1)显示,高光照0 h时,金幌叶片中ALA、PBG、尿卟啉(UrogenIII)和粪卟啉(CoprogenIII)含量均高于粉晶,且CoprogenIII是累积量最高的前体物质,是粉晶叶片中CoprogenIII含量的2倍,而ProtoIX、Mg-ProtoIX及原叶绿素酯(Pchl)含量则相反。说明金幌叶绿素合成过程中在CoprogenIII到ProtoIX处合成受阻。
高光照处理48 h时,金幌叶片出现红色至棕色的类病斑,然后面积逐渐增大,至168 h时达到最大(图1)。粉晶在高光照处理48 h时出现类病斑,然后类病斑面积在高光照处理96 h时达到最大,之后开始逐渐降低。类病斑的形成说明叶片受到光伤害,而光伤害与植物体内的ROS(活性氧簇)途径异常有关。SOD、POD和CAT是ROS途径3个重要的酶。金幌叶片中,SOD活性随着光照时间的增加而逐渐降低;CAT活性先增加到最大值(高光照48 h),然后逐渐降低,并且其活性一直高于对照粉晶叶片CAT活性,但是低光(高光照0 h)时则相反;POD活性先降低至最低值(高光照48 h)然后逐渐恢复到初始水平(高光照0 h和168 h差异不显著),并且其活性一直高于对照粉晶叶片POD活性,但是高光照48 h时相反(图3)。高光照处理168 h后,金幌叶片中SOD、CAT及POD活性分别是对照粉晶叶片中酶活性的0.87倍、1.34倍和1.94倍。 这些结果表明金幌叶片中的光伤害高于对照粉晶。
2.2光照度对紫薇金幌光合能力的影响
图4显示,高光照0 h和48 h时,光合参数在金幌和粉晶2个品种间均无显著差异;高光照96 h和168 h时,光合速率、气孔导度及蒸腾速率在2个品种间表现出显著差异,而胞间二氧化碳浓度在2个品种之间仍无显著差异。随着高光照处理时间的增加,金幌的净光合速率先升高后降低,而粉晶的净光合速率逐渐升高;粉晶的气孔导度先升高后持续降低,而金幌的气孔导度呈上升趋势;金幌和粉晶的胞间二氧化碳浓度均随光照处理时间的增加而增大;金幌的蒸腾速率开始时增加,但48 h后迅速降低,而粉晶的蒸腾速率呈上升趋势,但96 h的蒸腾速率与168 h时差异不显著,说明粉晶的蒸腾作用趋于稳定。随着高光照处理时间的增加,金幌气孔导度持续下降,随之蒸腾速率下降,同时净光合速率持续降低,而胞间二氧化碳浓度随之升高,说明金幌的光合同化能力在减弱。
2.3高光照处理对紫薇金幌光合系统II的伤害
高光照0 h时,Fv/Fm值在金幌和粉晶间不存在显著差异(表2)。随着高光照时间的增加,金幌叶片的Fv/Fm值逐渐降低,而粉晶叶片的Fv/Fm值先降低后趋于稳定。这些结果表明金幌和粉晶的PSII在高光照下均受到损伤,但是金幌的损伤更为严重。
高光照0 h时,金幌的ETR值显著高于粉晶。随着高光照时间的增加,金幌叶片的ETR值逐渐降低,而粉晶叶片ETR值先降低后逐渐恢复。高光照处理4 h后,金幌的ETR值降低了34.58%。ETR呈下降趋势,说明电子传递效率下降,可能有2方面原因,一是单位时间内电子产率下降,二是电子转运系统受损。这些结果也表明金幌PS II受到的伤害程度较重。随着高光照处理时间的延长,PS II没有得到恢复。
高光照0 h时,NPQ值在金幌和粉晶间不存在显著差异。随着高光照时间的增加,相较于粉晶,金幌中NPQ值呈现先降低后增加的趋势,并在高光照7 h时达到最大。NPQ值升高,说明金幌捕光色素转化的光能以熱耗散的形式散失的部分增加,金幌受到的光胁迫较粉晶严重。
以上结果显示,在高光照处理中,金幌受到的光胁迫较粉晶严重,导致PS II受到的伤害程度较重。
3讨论
3.1紫薇金叶品种金幌叶色变化可能与CPO活性的降低有关
叶绿素代谢是一个高度协调的酶促反应过程,需要一系列的酶参与。高等植物体内叶绿素合成过程中任何一个环节异常,都可能致使叶绿素含量发生改变,进而引起叶片颜色的变异。在叶绿素合成通路中,Coprogen III 转化成为Proto IX需要粪卟啉原氧化酶(CPO)的催化。Coprogen III为光敏化合物,其水平的降低可导致烟草(Nicotiana tabacum)、拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)等植物出现光依赖型叶片黄化[12-15]。在拟南芥CPO缺失突变体lin2(A. thaliana lin2)中,幼叶淡绿色,中部有病斑,病斑起初较小或条纹状[13]。紫薇金幌叶片在高光照条件下,叶片出现白斑并逐渐扩张。在抑制CPO表达的转基因植物中发现随着叶片病斑的扩大,抗氧化物酶的活性逐渐增加[12]。本研究中,高光照处理金幌叶片时也出现了lin2类似的表型。高光照处理7 d后,金幌叶片中SOD、CAT和POD等3个酶的活性分别是对照粉晶叶片中酶活性的0.87倍、1.34倍和1.94倍,与Kruse等[12]研究结果类似。
在水稻的一个PROB缺失突变体fgl中,ALA、Proto IX和Mg-ProtoIX等3个叶绿素合成前体物质含量在高光照下急剧下降[16]。在玉米的一个UROD缺失突变体Les22中,在高光照下尿卟啉含量较野生型增加了2~3倍,而Coprogen III含量未受到影响[17]。烟草中RNA干扰CPO表达可导致Coprogen III的积累[12]。本研究中,高光照0 h时,金幌叶片中ALA、PBG、UrogenIII和CoprogenIII含量均高于粉晶,且CoprogenIII是累积量最高的前体物质,是粉晶叶片中含量的2倍,而ProtoIX、Mg-ProtoIX及Pchl含量则相反。这些结果表明金幌叶绿素合成过程中在CoprogenIII到ProtoIX处合成受阻。紫薇金叶品种金幌的总叶绿素含量显著低于粉晶,而叶绿素a和叶绿素b在高光照处理7 d后分别下降了47.89% 和70.26%,叶绿素b的减少更为显著。
總之,本研究中通过对金幌和粉晶在低光照和高光照条件下叶片中抗氧化物酶活性、叶绿素及前体物质含量的测定,发现紫薇金叶品种金幌叶色变异是由于Coprogen III积累导致的。
3.2高光照处理对紫薇金幌PS II功能的影响
叶绿素缺失突变体为高等植物光合作用的研究提供了一个有力的工具[17]。叶绿素荧光参数是光合效率的重要指标,能够反映光合系统内部的调节过程,其动态变化蕴藏丰富的光合及生理信息。初始荧光(Fo)参数的改变反映了植物叶绿体中类囊体膜结构的变化,最大荧光(Fm)参数的大小反映了电子传递的最大潜力,可变荧光(Fv)参数反映了质体醌A (QA)的还原情况[18]。在水稻突变体vyl中,由于叶绿素少于野生型,ETR和NPQ值显著低于野生型,而Fv/Fm在两者间不存在显著差异[19]。玉米的一个黄绿叶突变体中,在生育期的各个阶段,其ETR、NPQ和Fv/Fm均低于野生型[20]。在本研究中,金幌的Fv/Fm值在低光照状态下均大于粉晶,高光照处理后显著减小,而Fv/Fm在粉晶中则是先减小后增大。在高光照处理7 h后金幌的Fv/Fm与对照粉晶相比,减少了53.75%。这一结果说明粉晶光系统II在最初受到了伤害,但之后逐渐恢复正常,而金幌受到伤害后没有得到恢复。
金幌的NPQ较高,说明其叶片接收的光能主要通过热耗散的方式消耗。低光照条件下,金幌的ETR高于粉晶;高光照处理后,金幌的ETR迅速降低,之后基本保持稳定,而粉晶的ETR先是急剧降低,而后恢复至正常水平。高光照处理7 h后,金幌的ETR比粉晶减少34.58%。ETR的变化表明,金幌在高光照下电子传递系统受到破坏,且没有得到恢复,而粉晶在突然暴露于高光照下时电子传递系统受到破坏,但光适应后迅速得到恢复。
参考文献:
[1]王淑安,王鹏,杨如同,等.紫薇新品种‘金幌[J]. 园艺学报, 2016,43(3):609-610.
[2]张振宇,王鹏,王淑安,等. ‘金薇叶片呈色的结构和生理基础研究[J].北方园艺, 2014(4):68-72.
[3]YUAN M, XU M Y, YUAN S, et al. Light regulation to chlorophyll synthesis and plastid development of the chlorophyll-less golden-leaf privet[J]. Journal of Integrative Plant Biology, 2010, 52 (9): 809-816.
[4]潘一鸣. 遮阴对金叶女贞和红继木叶色影响的研究[D].雅安:四川农业大学, 2011.
[5]王明玉,姜卫兵,彭丽丽,等. 遮阴对金叶女贞不同叶位叶片色素及光合特性的影响[J]. 江苏农业科学, 2013, 41(7): 166-169.
[6]LJUBESIC N, WRISCHER M, PREPEG T, et al. Chloroplast structure and function in wild-type and aurea-type leaves of the Japanese spindle-tree over there life span[J]. Acta Bot Croat, 2003, 62(1):1-10.
[7]施爱萍,张金政,张启翔,等. 不同遮荫水平下4个玉簪品种的生长性状分析[J]. 植物研究, 2004, 24(4): 486-490.
[8]李淑顺,唐玲,李倩中,等. 红色鸡爪槭叶绿素荧光特征参数及其与日灼伤害的关系[J]. 江西农业学报, 2010, 22 (11): 15-17.
[9]李倩中,刘晓宏,李淑顺,等. 不同季节3个鸡爪槭品种的光合与生理特性[J].江苏农业科学,2012,40(6):149-152.
[10]BRIAN M, LECKIE B M, STEWART C N J. Agroinfiltration as a technique for rapid assays for evaluating candidate insect resistance transgenes in plants[J]. Plant Cell Rep, 2011, 30: 325-334.
[11]BOGORAD L. The Biosynthesis of Chlorophylls[M]. Amsterdam:Elsevier,1966:481-510.
[12]KRUSE E, MOCK H P, GRIMM B. Coproporphyrinogen III oxidase from barley and tobacco-sequence analysis and initial expression studies[J]. Planta, 1995, 196(4): 796-803.
[13]ISHIKAWA A, OKAMOTO H, IWASAKI Y, et al. A deficiency of coproporphyrinogen III oxidase causes lesion formation in Arabidopsis[J]. The Plant J, 2001, 27(2): 89-99.
[14]GUO C Y, WU G H, XING J, et al. A mutation in a coproporphyrinogen III oxidase gene confers growth inhibition enhanced powdery mildew resistance and powdery mildew-induced cell death in Arabidopsis[J]. Plant Cell Rep, 2013, 32(5): 687-702.
[15]WANG J, YE B, YIN J, et al. Characterization and fine mapping of a light-dependent leaf lesion mimic mutant 1 in rice[J]. Plant Physiol Bioch, 2015, 97: 44-51.
[16]SAKURABA Y, RAHMAN M, CHO S, et al. The rice faded green leaf locus encodes protochlorophyllide oxidoreductase B and is essential for chlorophyll synthesis under high light conditions[J]. The Plant J, 2013, 74(1): 122-133.
[17]HU G, YALPANI N, BRIGGS S P, et al. A porphyrin pathway impairment is responsible for the phenotype of a dominant disease lesion mimic mutant of maize[J]. Plant Cell, 1998, 10(7): 1095-1105.
[18]王俊忠,张超男,赵会杰,等. 不同施肥方式对超高产夏玉米叶绿素荧光特性及产量性状的影响[J]. 植物营养与肥料学报, 2008, 14(3):479-483.
[19]DONG H, FEI G L, WU C Y, et al. A rice virescent-yellow leaf mutant reveals new insights into the role and assembly of plastid caseinolytic protease in higher plants[J]. Plant Physiol, 2013, 162(4): 1867-1880.
[20]ZHONG X M, SUN S F, LI F H, et al. Photosynthesis of a yellow-green mutant line in maize[J]. Photosynthetica, 2015, 53(4): 499-505.
(責任编辑:张震林)
猜你喜欢
语文世界(小学版)(2021年5期)2021-06-04
课程教育研究·学法教法研究(2019年18期)2019-10-08
少儿科技(2019年9期)2019-09-10
东坡赤壁诗词(2019年1期)2019-04-30
山东农业科学(2018年3期)2018-03-29
山东农业科学(2018年11期)2018-01-17
作文成功之路·教育教学研究(2016年4期)2016-06-12
江苏农业科学(2016年2期)2016-04-11
民间文学(2015年9期)2015-10-21
时尚北京(2014年8期)2014-08-26
