
家蚕胰脂肪酶相关蛋白1基因(BmPLRP1)的克隆及表达分析
2020-04-23 08:23李玉霞钱荷英郝长富刘明珠赵国栋徐安英
河北农业大学学报 2020年1期
李玉霞,李 刚,2,钱荷英,2,郝长富,刘明珠,2,赵国栋,2,徐安英,2
(1.江苏科技大学 生物技术学院,江苏 镇江 212018;2.中国农业科学院 蚕业研究所,江苏 镇江 212018)
胰脂肪酶Pancrelipase(PL)主要是由胰脏胰腺细胞分泌,是参与肠道消化的主要酶,属于丝氨酸水解酶类,可分解摄食获得的脂类物质,在小肠中协助消化甘油三酯起着重要作用[1-2]。它利用胆汁盐可乳化食物中50%~70%膳食脂肪类[3]。PL是由两个域组成,一个含有活性位点的N末端域,另一个含有辅脂酶结合的C末端域。这种酶被验证具有两种不同的构象,一种是不活跃的封闭构象,在这种构象中有一种特殊的环状物即皮瓣[4],它覆盖了活性位点以及通过重新定位皮瓣来揭示活性位点而形成的活跃的开放构象[5]。
对不同物种的胰腺cDNA文库进行筛选,发现除了甘油三酯脂酶(Pancreatic triglyceride lipase,PTL)外,还存在胰脂肪酶相关蛋白1(Pancreatic lipase-related protein 1, PLRP1)与胰脂肪酶相关蛋 白 2(Pancreatic lipase-related protein 2, PLRP2)两种脂质相关蛋白[6-9],同属脂肪酶家族成员,与PTL相比这两种酶的分泌量极低,所以一直未被发现[8,10],其中PTL与PLRP1、PLRP1分别有68%和65%的氨基酸序列同源性。PLRP1 的分泌与生物的生长发育有着密切的联系, 研究表明,PLRP1的分泌量会随着发育时间的推移逐渐降低,而 PLRP2的表达与分泌却与发育状态无关[6]。PL、PLRP1和PLRP2在结构上很接近,但它们显示出非常不同的功能特性[11],其中PLRP1 在通常状态下无活性,但可以在一些特殊脂类物质的诱导下显示出极低的酶活特性[12-13]。研究表明PLRP1 在活性位点的氨基酸残基经过人为替换可以使其表现极强的活性[14],也有研究显示PLRP1可能通过与PTL和PLRP2竞争结合辅酶而间接影响脂肪的消化吸收[15],PLRP1与PTL进行竞争结合辅酶,会导致辅脂酶缺乏,当PLRP1缺乏时,则会增加脂肪的消化,最终导致胰岛素抵抗与肥胖症的发生[16]。尽管PLRP1的生理功能尚未明确,但一些研究报道该基因的改变可能涉及炎症、肥胖、高血脂、动脉硬化和前列腺癌等过程[17-18]。目前对PLRP1的研究大多集中在人、小鼠中,在昆虫中鲜有报道。
家蚕(Bombyx mori)是重要的经济昆虫,也是鳞翅目昆虫的模式生物。对家蚕PLRP1的研究不仅有助于了解BmPLRP1功能,也可以为其他昆虫的PLRP1活性研究提供一定的借鉴参考。本研究通过克隆PLRP1的编码基因,研究其在家蚕不同组织及受氟化物侵染后不同发育阶段的相对表达情况,为探究BmPLRP1的相关功能奠定基础。
1 材料与方法
1.1 供试蚕品种及处理
供试蚕品种‘733xin’由中国农业科学院蚕业研究所提供,按常规方法催青、饲养。前期研究表明,家蚕品种‘733xin’对氟化物污染敏感[19]。收蚁时,蛾区半分法将同一蛾区分成2区,一半喂食正常新鲜桑叶,另一半至3龄起蚕时添食经NaF处理的桑叶(将新鲜桑叶浸泡于100 mg/kg NaF溶液中5 min后自然晾干)。到5龄第1、3、5天取其中肠、脂肪体、血液、头、表皮、马氏管、丝腺 、生殖器、气管丛等组织材料置于-80℃备用。
1.2 家蚕胰脂肪酶相关蛋白1基因的克隆
1.2.1 总RNA提取及cDNA逆转录 取蚕品种‘733xin’ 5龄3天中肠,放入无RNase的研钵中加液氮充分研磨成粉末取约0.1 g样品至1.5 mL的无DNA/RNAse离心管中,用RNA 提取试剂盒RNAiso Plus (Takra,No.9109)提取中肠组织的总RNA,1%琼脂糖凝胶电泳验证总 RNA 的完整性,用微量核酸蛋白定量仪测定RNA 浓度。选用电泳条带清晰完整且OD260/OD280为 1.9~2.0的RNA用 PrimeScript ™ 1st Strand cDNA Synthesis Kit(Takara,No.6110A) 反转录试剂盒制备cDNA然后将制备的 cDNA 置于-20℃冰箱中保存备用。
1.2.2 引物设计 通过SilkDB数据库(http://silkworm.genomics.org.cn/) 和GeneBank数 据库(http://www.ncbi.nlm.nih.gov/ Genbank)的交叉比对搜索BmPLRP1的编码基因,根据同源基因序列,使用Prime 6.0软件设计基因扩增引物,上游引物P-F序列为5’-CGTTCTCGGTAACGCACGTA-3’,下游引物P-R序 列 为 5’-ACGACACATCTTTACAATAAAACGC-3’,引物由上海生工生物工程(上海)股份有限公司合成。
1.2.3BmPLRP1基因克隆及鉴定 以家蚕5龄3 d中肠的cDNA为模板,扩增BmPLRP1基因序列。25 μL PCR体系为: 10×ExTaqBuffer (含20 m mol/LMg2+)2.5 μL, 2.5 mM dNTP 2μL,10 μmol/L上下游引物各 1 uL ,5 U/μL ExTaq酶 0.3μL,1 000 ng/μL 的cDNA模板1μL,加双蒸水补足至25 μL。反应条件: 95℃预变性5 min,95℃变性50 s,57℃退火30 s,72℃延伸1 min,共34个循环,72℃终延伸10 min,4℃保存。PCR扩增产物经1%琼脂糖凝胶电泳检测并纯化后连接到pMD19-T载体(Takara,No.3271),转入E.coilDH10-B 感受态细胞,通过蓝白斑筛选挑选阳性克隆,挑取阳性单克隆菌落培养后进行菌液PCR鉴定,选取目的条带大小正确的菌液送至上海生工生物工程(上海)股份有限公司测序。
1.3 生物信息学分析
利用Singal P 4.1 server(http://www.cbs.dtu.dk/services/SinglP/)预测信号肽;使用TMHMM Server v.2.0(http://www.cbs.dtu.dk/services/TMHMM/)预测跨膜结构域;使用AntheProt 6.9软件预测蛋白质的二级结构;通过Blastp工具进行蛋白质保守结构域分析;利用网上蛋白质活性位点预测分析软件Scanprosite(https://prosite.expasy.org/)进行氨基酸序列活性位点分析;利用NetPhos 3.1 Server(http://www.cbs.dtu.dk/services/NetPhos/)预测磷酸化位点;利用NetNGlyc 1.0 DictyOGlyc 1.1 Server (http://www.cbs.dtu.dk/services/DictyOGlyc/)预测O-糖基化位点;利用Bioedit软件的Clustal-W进行氨基酸序列多重比对,应用MEGA 7.1软件以Neighbor-Joining(NJ)方法构建系统进化树,将Bootstrap值设置为1 000。
1.4 BmPLRP1基因的组织表达谱分析
提取家蚕5龄第3天幼虫中肠、血液、脂肪体、头、表皮、马氏管、丝腺 、生殖器、气管丛的总RNA,用反转录试剂盒(Takara,No.RR047A)将提取的各组织的mRNA分别反转录成cDNA。根据该基因的CDS序列设计引物 F(5'-GCGGTTTCTGATGTTTGG-3') 和 R(5'-CGTTCTTGTGATGTTCGTAT-3')。以各组织cDNA为模板,以BmActin 3基因为内参基因进行半定量PCR反应,对其扩增产物进行1%琼脂糖凝胶电泳检测。
1.5 家蚕‘733xin’经NaF处理后BmPLRP1的相对表达量检测
提取处理组与空白组收集的4个时间点的家蚕中肠、脂肪体、血液、丝腺等组织的总RNA以提取的RNA为模板用反转录试剂盒合成cDNA。设计qRT-PCR引物上游引物F(5'-GCGGTTTCTGATGTTTGG-3’)和下游引物R(5'-CGTTCTTGTGATGTTCGTAT-3’),引 物 由上海生物生工生物工程股份有限公司合成。使用SYBRPremix ExTaq™ II(Tli RNaseH Plus)试剂盒,以反转录合成的cDNA 进行实时荧光定量PCR反应。数据分析采用Excel和SPSS19.0软件进行。
2 结果与分析
2.1 家蚕PLRP1基因的克隆、鉴定与序列特征
以P-F/R为引物,家蚕‘733xin’中肠cDNA为模板扩增PLRP1基因,获得一条1 100 bp左右的条带(图1),将该片段进行胶回收并克隆到pMD19-T载体,重组质粒菌液PCR鉴定,经测序获得BmPLRP1基因的序列信息。将测序结果进行序列比对分析,结果表明克隆的BmPLRP1基因序列与NCBI数据库中登陆号为XP_004929472.1的序列编码以及SilkDB数据库中编号为 BGIBMGA010401-TA 的基因序列编码同一蛋白质。BmPLRP1的cDNA序列长1 462 bp,由90 bp的5’端、996 bp的编码区和373 bp的3’端序列构成,3’端序列包含1个多聚腺苷酸化信号(aataaa)和典型的poly(A)结构。
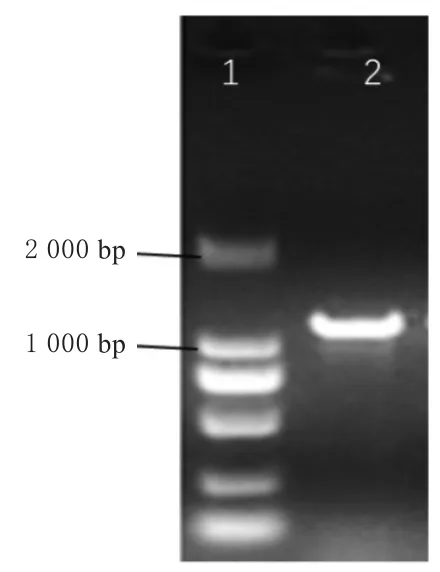
图1 家蚕PLRP1基因的RT-PCR扩增产物的凝胶电泳图谱Fig.1 The PCR gel electrophoresis of BmPLRP1 gene
2.2 BmPLRP1蛋白的基本特征分析
生物信息学分析结果显示,BmPLRP1基因位于家蚕第12号染色体的nscaf2993位点,核苷酸序列由7个外显子和6个内含子构成,外显子/内含子边界符合GT-AC规则,ORF长度为996 bp,编码331个氨基酸。利用各种软件分析BmPLRP1基因,预测其编码蛋白分子质量为37.1 kD,等电点为8.72。预测的蛋白二级结构如图2所示,包括α螺旋(α-helix)24%、β-折叠(β-sheet)25%、β-转角(β-turn)12%、无规则卷曲(Random coil)39%。该蛋白含有全部的基本氨基酸,其中丙氨酸(Ala)与甘氨酸(Gly)含量最高,同为8.13%;半胱氨酸(Cys)含量最低;无信号肽序列,为非分泌型蛋白;没有跨膜结构域,属于非跨膜蛋白;含有1个Pancreat_lipase_like结构域(图3),从序列分析得出,该结构普遍存在于许多脂肪酶中。对BmPLRP1蛋白的氨基酸序列进行了活性位点分析,结果在142~151 aa位点上发现有一个LIPASE_SER(Lipases, serine active site)活性位点,其氨基酸序列为VHLIGHSLGS。这个活性位点的存在,可能对BmPLRP1蛋白的活性激活具有重要意义,同时也证明了该蛋白是一种脂肪酶。预测该蛋白有35个潜在的磷酸化活性位点(图4),容易通过磷酸化或去磷酸化进行蛋白修饰;预测该蛋白不含N-糖基化位点,含有一个O-糖基化位点,易通过O-糖基化进行蛋白修饰。

图2 预测的BmPLRP蛋白的二级结构Fig.2 Predicted secondary structure of BmPLRP protein
2.3 系统进化分析
将BmPLRP1编码的氨基酸与GenBank数据库中的棉铃虫(Helicoverpa armigera)、野桑蚕(Bombyx mandarina)、斜纹夜蛾(Spodoptera litura)、丛林斜眼褐蝶(Bicyclus anynana)、脐橙螟(Amyelois transitella)、 小 菜 蛾(Plutella xylostella)、 埃及伊蚊(Aedes aegypti)、巴士果蝇(Drosophilabusckii)的PLRP1蛋白进行比对(图5),序列相 似 性 为 98.82%、80.30%、78.79%、78.12%、76.74%、72.73%、76.29%、46.05%、42.90%。 构建的系统进化显示BmPLRP1 与鳞翅目昆虫野桑蚕PLRP1(XP_028040615.1)的进化关系较近,与双翅目昆虫埃及伊蚊PLRP1(EJY57663.1)、巴氏果蝇PLRP1(XP_017854645.1)的进化关系较远(图6)。

图5 家蚕PLRP1蛋白的多重序列比对Fig.5 Multiple sequence alignment of PLRP1 protein form Bombyx mori and other species

图6 家蚕PLRP1蛋白和其他物种同源蛋白的N-J系统进化树Fig.6 N-J phylogenetic tree of Bombyx mori PLRP1 protein and homologous proteins of other species
2.4 BmPLRP1基因在家蚕各个组织中的表达特征分析
采用半定量RT-PCR对BmPLRP1基因在家蚕5龄3天幼虫不同组织的表达特征进行分析(图7)。

图7 半定量RT-PCR检测家蚕PLRP1基因在5龄3天幼虫各组织器官中的表达Fig.7 Expression of BmPLRP1 gene in various tissues of day 3 silkworm larvae of the 5th instar detected by RT-PCR
结果表明,BmPLRP1基因在头部、表皮、丝腺、生殖器和气管丛中有明显表达,其次是中肠和血液的表达量,而在血液和脂肪体中的表达最低。
2.5 BmPLRP1基因在不同时期的表达
利用qRT-PCR进行进行分析BmPLRP1基因在733和经氟化物处理过的733品种的不同时期不同组织的表达情况。经氟化物处理过后,4龄起蚕、5龄1、3、5天的中肠、血液、丝腺组织中,BmPLRP1基因的表达量相对升高,而在脂肪体组织中,BmPLRP1基因的表达在5龄后期上调,推测可能与氟化物的侵染有一定关系(图8)。
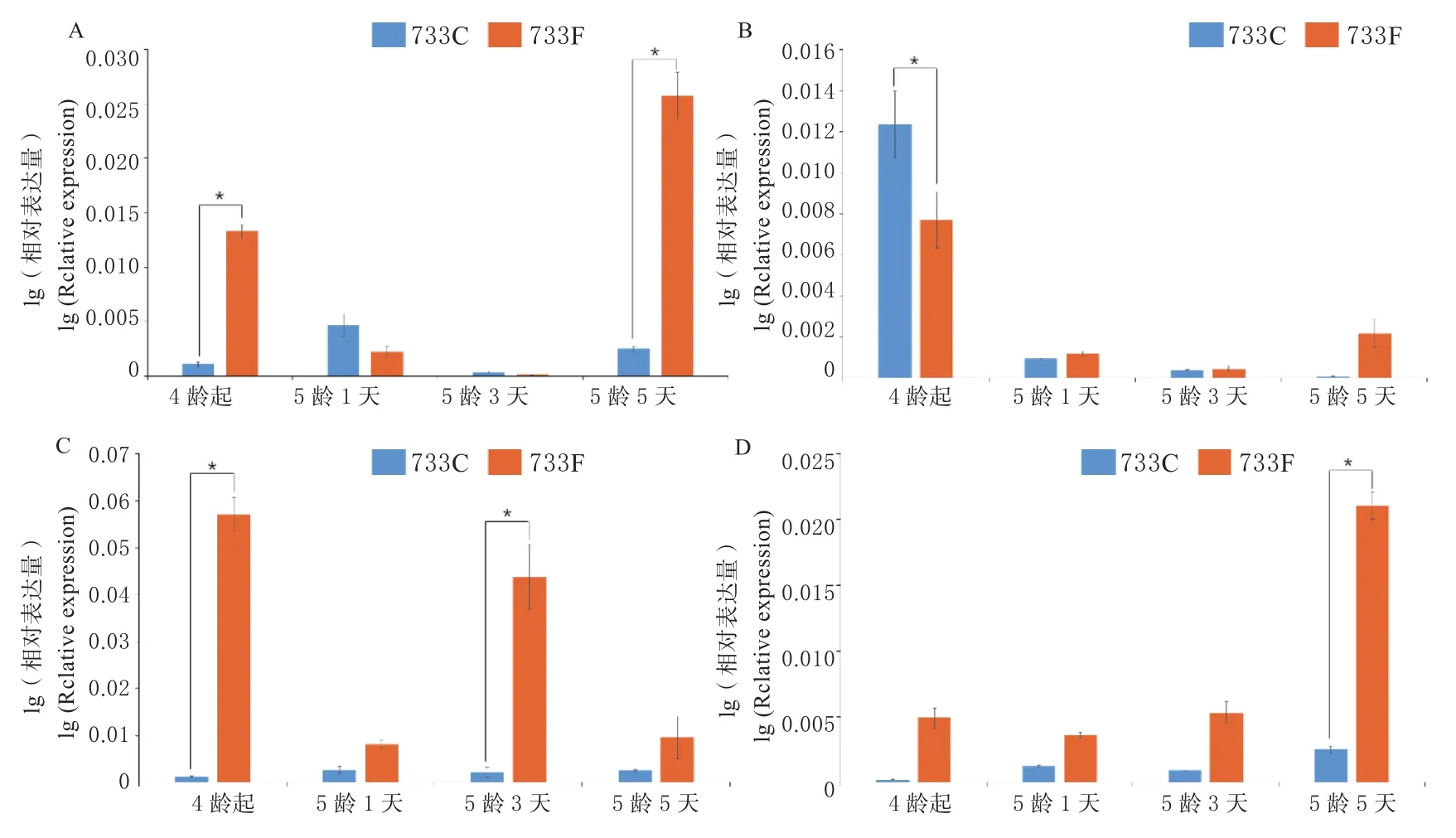
图8 BmPLRP1基因在733和733F中不同时期不同组织中的表达Fig.8 Expression of the BmPLRP1 in different tissues at different times in 733 and 733F
3 结论
本研究利用生物信息学及分子生物方法在家蚕中克隆得到BmPLRP1基因。BmPLRP1与其他物种的PLRP1在氨基酸序列上都具有一个相同的结构域,该结构普遍存在于许多脂肪酶,暗示BmPLRP1蛋白可能与其他的脂肪酶蛋白相互作用形成蛋白复合体。采用生物信息学分析得到该蛋白存在一个LIPASE_SER(Lipases, serine active site) 活 性 位点,暗示可能对BmPLRP1蛋白的活性激活具有重要意义;同时存在着潜在的磷酸化活性位点,表明容易通过磷酸化或去磷酸化进行蛋白修饰;还含有一个O-糖基化位点,易通过O-糖基化进行蛋白修饰,说明该蛋白是一糖蛋白。半定量RT-PCR检测BmPLRP1基因在头部、表皮、丝腺 、生殖器和气管丛中有明显表达,推测该基因在神经调节、生殖发生可能具有一定作用。家蚕胰脂肪酶相关蛋白1在通常状态下无活性,但可以在一些特殊脂类物质的诱导下显示出极低的酶活特性。本实验取材家蚕中肠组织而非肠道内容物对该基因的表达量进行检测,经3次重复显示目前结果,确认为在中肠内的相对表达量比在表皮、生殖腺、气管丛组织中的表达量较低。后期100 mg/Kg NaF处理后的对照实验表明,经过NaF诱导后在中肠组织、脂肪体、血液、丝腺等组织是有诱导性上调,表明该基因对NaF中毒有一定的应答反应。研究结果为进一步鉴定与氟侵染的关系,以及确定该蛋白质在家蚕生长发育中的具体功能作用奠定了一定的基础。
猜你喜欢
四川蚕业(2022年2期)2022-11-19
四川蚕业(2022年2期)2022-11-19
中华胰腺病杂志(2022年4期)2022-08-23
中国生物防治学报(2022年3期)2022-07-09
保健文汇(2022年1期)2022-06-06
四川蚕业(2021年2期)2021-03-09
四川蚕业(2021年1期)2021-02-12
放射学实践(2020年2期)2020-03-13
求知导刊(2019年15期)2019-08-30
广西科技大学学报(2018年2期)2018-09-10
