
遮光对糠椴幼苗光合特性的研究
2020-05-22 09:10刘兴赵丹杨晓金梦然刘煜光史宝胜
林业与生态科学 2020年1期
刘兴,赵丹,杨晓,金梦然,刘煜光,史宝胜
(河北农业大学 园林与旅游学院,河北 保定 071000)
光是植物光合作用和生长发育的重要影响因子[1]。植物在长期的进化过程中,形成了自身特有的需光特性[2]。植物可以在遮光环境下产生可塑性反应,通过增加叶绿素含量、减小栅栏组织和海绵组织厚度等来保持最大吸收光量子的能力[3-4];通过降低光饱和点和光补偿点,增加表观量子效率等来提高光能利用效率[5-6];通过降低暗呼吸速率等来减少碳的消耗,将更多的碳用于高生长,以期能最大程度地获得光照[7]。这些生理方面的调节对于植物在遮光环境下保持碳的平衡起到了至关重要的作用[8]。
糠椴(TiliamandshuricaRup et Maxim.)为椴树科椴树属落叶乔木。近年来,对糠椴的研究主要集中在椴树枝叶解剖结构、糠椴组培苗的诱导生根、蜜高产措施、糠椴检测大气氯污染等领域[9-12]。而在遮光对糠椴光合及生理影响方面未见报道。在椴树属的相关研究中,孙欣欣等研究表明紫椴通过调节叶片P、K含量来适应弱光环境[13]。陈婕等研究表明在光环境的转变下,蒙古栎和紫椴都通过增加叶绿素含量表现出对低光环境的适应[14]。本研究以糠椴幼苗为试材,利用盆栽遮光方法,分析遮光对糠椴叶片光合日变化、光响应参数、叶绿素荧光、色素含量的影响,揭示在不同光照条件下糠椴的光合生理反映特性,以明确紫椴幼苗适宜生长的光环境条件,为幼苗的栽培生产和园林应用提供科学依据。
1 材料与方法
1.1 试验地概况
试验地位于河北省保定市河北农业大学园林与旅游学院试验田。该地区位于经度 115.47 E、纬度38.87 N。属暖温带亚湿润气候区。
1.2 试验材料
选取生长良好、长势一致的2 a生糠椴幼苗为试验材料。植株载于盆中(花盆直径23.5 cm,高14 cm)。试验盆土为苗圃地园土、砂和营养土自然风干后按照2∶2∶1混合,常规管理。
于2018年7月22号用不同遮光率的黑色遮阳网进行遮光处理,分别为透光率70%(轻度遮光,LS)、透光率40%(中度遮光,MS)、透光率10%(重度遮光,SS),以自然光照处理(即透光率100%)为对照(CK),每处理8株,3次重复。在处理60 d时进行光合和生理指标测定。测定叶片均取自植株顶梢向下第6~8片叶。叶绿素测定的叶片于早8:00采集后,置于冰盒,于-80℃贮存备用。使用单因素随机区组设计试验,每小区8株,重复3次。
1.3 测定方法
1.3.1 叶绿素含量测定 参照李合生方法,测定叶绿素a含量和叶绿素b含量[15]。
1.3.2 光合参数的测定 采用Li-6400便携式光合测定仪测定。参照王建华等的方法[16]。于阳光充足的晴天,从08:00到18:00每2 h对不同处理的叶片进行净光合速率(Pn)、胞间CO2浓度(Ci)、气孔导度(Gs)、蒸腾速率(Tr)、水分利用率(WUE)参数测定,每处理测定7片叶。利用LED光源,测定不同处理叶片在不同光照强度下的净光合速率,绘制光合-光响应曲线。参照叶子飘的方法计算光补偿点(LCP)、光饱和点(LSP)、光饱和时最大净光合速率(Pmax)、暗呼吸速率(Rd)、表现量子效率(AQY)[17]。
1.3.3 叶绿素荧光参数的测定 使用PM100便携式荧光仪,于上午8:00到10:00对糠椴幼苗的叶片暗适应20 min后,测定植物叶片的主要荧光参数:最小荧光(Fo)、最大荧光(Fm)、PSII潜在活性(Fv/Fo)、PSII最大光化学效率(Fv/Fm)、吸收的光能被反应中心捕获的量子产量(φPo)、激子被反应中心捕获后,促进电子传递到电子传递链中超过QA的电子受体的激子与促进QA还原激子的比值(Ψo)、反应中心所吸收的光能应用于电子传递的量子产量(φEo)、用于热耗散的量子产量(φDo)、单位反映中心吸收的能量(ABS/RC)、单位反应中心捕获的用于还原QA的能量(TRo/RC)、单位反应中心捕获的用于电子传递的能量(ETo/RC)、单位反映中心耗散掉的能量(DIo/RC),每处理测定5片叶,3次重复。
1.3.4 数据处理及统计分析 所有数据转入Excel中进行计算,方差分析采用SPSS18.0参数选择ANOVA进行统计分析。
2 结果与分析
2.1 遮光对光合日进程的影响
遮光对净光合速率日变化的影响如图1所示。
遮光处理下对Ci、Gs、Tr、WUE参数的日进程如图2所示。
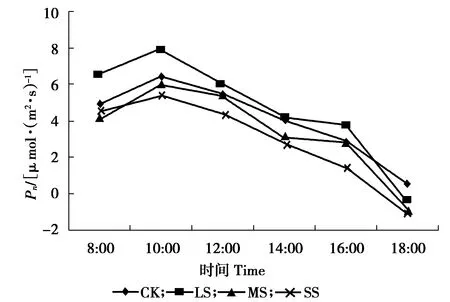
图1 遮光处理下对净光合速率日变化的影响
遮光处理下对Ci、Gs、Tr、WUE参数的日进程如图2所示。
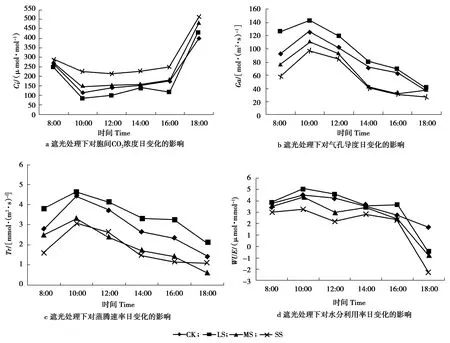
图2 遮光处理下相关参数的日进程
Figure 2 Daily process of relevant parameters under shading
由图1、图2可知,遮光后CK、LS处理的Pn呈双峰曲线型,MS、SS呈单峰型。单峰峰值出现在10:00,双峰峰值分别出现在10:00和16:00,且上午峰值高于下午。在LS处理的上午峰值比下午峰值高4.13 μmol /( m2· s)。
其中Gs、Tr、WUE变化趋势与Pn基本一致。Gs、Tr在LS处理的上午峰值分别比下午峰值高73.68 μmol /( m2· s)和1.38 μmol /( m2· s)。WUE在10:00和16:00达到峰值。如:在LS处理的上午峰值比下午峰值高1.34 μmol/mol。Ci在遮光下整体呈现升高趋势,在10:00达到最低,CK、MS、SS处理分别比LS提高了41.22%、83.27%和174.66%。
2.2 遮光对光响应及各参数的影响
遮光后糠椴光响应曲线的变化如图3所示。

图3 遮光对糠椴光响应曲线的变化
Figure 3 Light response curve under different shading conditions
在全日照和遮光环境下,幼苗净光合速率(Pn)随光合有效辐射(PAR)升高先逐渐增大,最后渐趋平缓。随光照的增强,遮光处理与对照间的差距逐渐增大,如在光合有效辐射为1 400 μmol /( m2· s)时,LS处理下的Pn比CK增加了19.12%,而MS、SS处理的比CK降低了25%、33.82%。
遮光下糠椴光响应曲线参数的变化如表1所示。
表1 遮光对糠椴光响应曲线参数
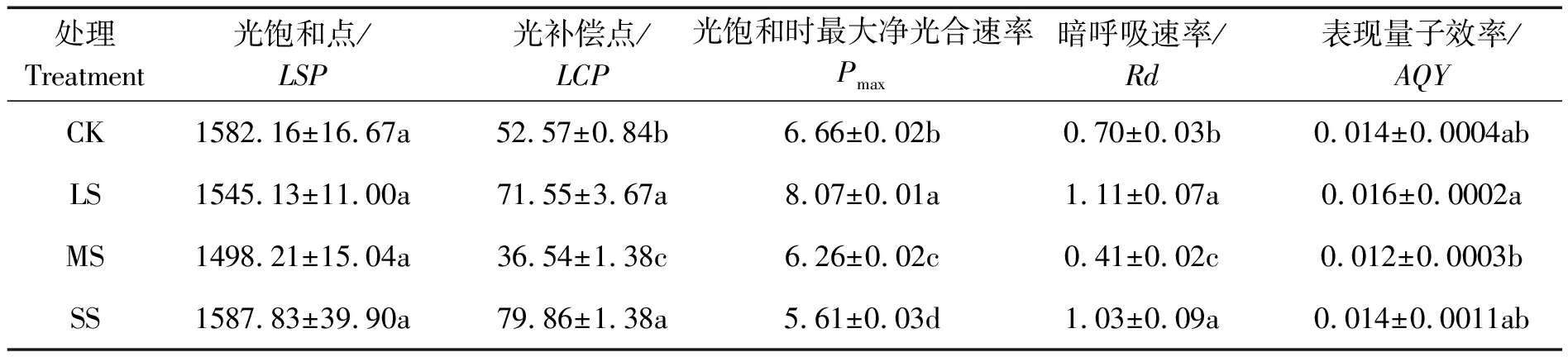
Table 1 Light response curve parameters under different shading conditions μmol/(m2·s)
注:小写字母表示不同遮光度之间差异达到显著水平(P<0.05),以下同。
遮光降低了幼苗的LSP、AQY和Pmax,并提高了LCP和Rd。LS时的LSP、AQY和Pmax与CK相比分别降低了2.34%、-14.29%、-21.17%。LCP和Rd与CK比提高了36.10%、58.57%。LS处理具有最高的Pmax,显著高于其余3个处理。在LS、MS、SS处理下,糠椴AQY分别为CK的14.29%、-14.29%、100%。
2.3 遮光对糠椴叶绿素荧光参数的影响
遮光后糠椴叶绿素荧光参数的变化如图4所示,在遮光处理下,叶绿素荧光参数Fo差异不显著,而Fm在MS处理下获得最大值。随着遮光度增加,Fv/Fo和Fv/Fm呈升高趋势,LS、MS、SS处理的Fv/Fo分别比CK增加14.09%、42.68%、42.00%;Fv/Fm比CK增加3.66%、9.34%、9.42%。
遮光后糠椴能量分配比率的变化和PSII比活性参数的变化如表2和表3所示。遮光处理后,Ψo、φEo的值逐渐增加,φPo的值变化不显著,而φDo的值则逐渐减少,在SS处理下分别比CK增加了9.50%、24.42%、35.07%,而φDo的值减少了25.18%。遮光处理后,ETo/RC、TRo/RC的值变化不显著。在SS处理下ABS/RC的值比CK降低了15.72%。MS、SS处理下DIo/RC的值,在SS处理下比CK降低了33.73%、37.35%。
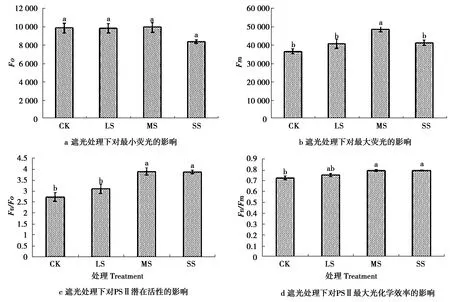
图4 遮光后糠椴叶绿素荧光参数的变化
Figure 4 Changes of chlorophyll fluorescence parameters in shading

表2 遮光后糠椴能量分配比率的变化

表3 遮光后糠椴PSII比活性参数的变化
2.4 遮光对糠椴叶绿素含量的影响
遮光后糠椴叶绿素含量的变化如表4所示。叶绿素b和叶绿素总量随遮光度的升高而增加。如在SS处理下叶绿素b和叶绿素总量分别比CK增加了114.29%和76.03%。且各遮光处理下叶绿素a、叶绿素b与CK间的差异达到显著水平。叶绿素a/b随遮光度的增加而降低,LS、MS、SS处理下分别比CK降低-4.47%、13.65%、23.57%。MS、SS处理下均与CK达到显著水平。
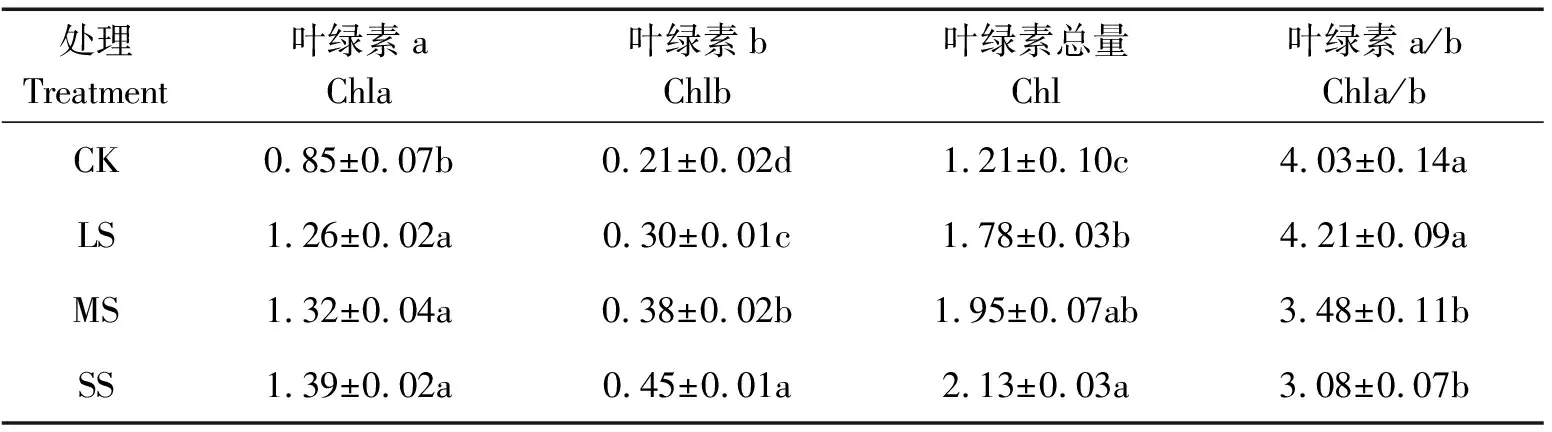
表4 遮光后糠椴叶绿素含量及比值的变化
3 讨论
影响植物光合作用的重要因素有光照、水分、温度和浓度等,其中光照是影响植物生长发育的最重要因素之一[18-19]。本试验中,糠椴在不同光照处理的日变化表现为双峰型曲线。其中净光合速率日变化整体由大到小依次为:LS、CK、MS、SS。说明LS更有利于糠椴幼苗的光合作用。
光响应曲线能有效反映出光合速率随光强改变的变化规律,能体现出不同环境对植物光合能力影响程度[20]。通常阳生植物的LCP在10 μmol/(m2·s)以上,LSP在1 000 μmol/(m2·s)以上[21]。本试验中糠椴幼苗在全光照下有较高的LCP(52.57 μmol/(m2·s))和LSP(1 582.16 μmol/(m2·s)),表现为阳生植物。通常在适宜的生长条件下,植物的AQY为0.02~0.05,其值越大,表明植物吸收与转换光能的色素蛋白复合体越多,植物利用弱光的能力越强[22-23]。不同遮光处理下糠椴幼苗的AQY均处于适宜范围内,而LS处理下AQY最高,表明轻度遮光处理有利于提高糠椴幼苗的光合利用率,且LS处理下利用弱光能力最强。AQY的值与对照相比在LS、SS处理下反而不显著,可能是因为在试验后期,植物本身物候期影响,植物由旺盛生长期向凋落期转变,代谢变慢,对外界环境变化的敏感性降低,从而出现光合指标回降现象。
叶绿素荧光技术广泛应用到植物生理的测定中[24]。有试验得出,Fv/Fm和Fv/Fo的升高有助于光合色素将其捕获的光能快速、高效的转化为化学能从而供给碳同化,提高光合速率,反映出植物在低光强环境中的适应性[25-27]。试验结果表明,随着遮光度增加,Fv/Fo和Fv/Fm逐渐升高。表明糠椴幼苗在遮光处理下有适应性,这与本试验中对表观量子效率分析得出的结论一致。
与全日照处理相比,遮光后糠椴φPo、Ψo均显著高于全光照,φEo在MS、SS处理下均显著高于全光照,φDo在MS、SS处理下则显著低于全光照,说明遮光后叶片吸收光能被反应中心捕获率、电子传递到QA-下游电子受体概率以及吸收光能将电子传递给超过QA的其他电子受体概率均显著提高,而用于热耗散的比率显著降低[28]。遮光对糠椴PSⅡ活性产生显著影响,主要表现为ETo/RC的升高和DIo/RC的降低,说明遮光明显提高了PSⅡ活性,更多光能用于电子传递,大大降低热耗散光能消耗。但是随遮光率的增加糠椴Fo的减少、φEo值的增加会使单线态叶绿素不能及时返回三线态叶绿素,单线态叶绿素的积累会将能量传递给O2,形成活性极强的单线态氧,这会引起光合机构的失活甚至破坏[29]。说明在SS处理下会引起光合机构的失活甚至破坏。
叶绿素是植物的光合色素,其含量和比例的变化不仅影响叶片呈色,也直接影响植物的光合效率[30]。叶绿素b主要吸收蓝紫光,其含量增加有利于植物对蓝紫光的利用,提高对弱光环境的适应性,因此通常情况下,叶绿素含量高,叶绿素a/b比值小的植物具有较强的耐阴性[31]。试验中在遮光处理下糠椴叶绿素b、叶绿素总量均随显著高于全光照,叶绿素a/b比值在MS、SS处理下均显著低于全光照,表明糠椴对遮光环境具有适应能力。
4 结论
综上所述,糠椴为阳生植物,幼苗对低光环境具有一定的适应性。轻度遮光环境下,糠椴幼苗叶绿素含量增加,且日平均净光合速率和最大净光合速率最高,光合能力强,说明轻度遮光下有利于提升幼苗的光合特性,促进幼苗生长。且幼苗光响应参数AQY、LCP、Pmax显著提升,LSP和Rd降低,说明轻度遮光提高了糠椴幼苗的光合利用率,更适宜糠椴幼苗生长。
猜你喜欢
今日农业(2021年10期)2021-11-27
阅读(科学探秘)(2020年8期)2020-11-06
中国果业信息(2019年1期)2019-01-05
生物学教学(2017年9期)2017-08-20
新课程·下旬(2017年7期)2017-08-14
中国火炬(2015年12期)2015-07-31
西北林学院学报(2015年5期)2015-01-03
食品工业科技(2014年6期)2014-05-10
中国火炬(2013年3期)2013-07-24
植物营养与肥料学报(2011年2期)2011-10-26
