
内蒙古锡林郭勒草原主要野生蘑菇分子进化及其系统发生学
2020-06-21 15:35李亚娇孙国琴郭九峰
江苏农业科学 2020年9期
李亚娇 孙国琴 郭九峰


关键词:草原蘑菇;ITS序列;分子系统发生学;系统进化树;分化时间
系统发生(phylogeny)也称系统发育、系统演化,是某一个类群的形成和发展过程,也就是进化或演化关系[1-2]。草原蘑菇主要生长在我国内蒙古中部草原地区,在草原上形成蘑菇圈,维持牧草与草原蘑菇共生的生态体系[3]。目前对草原蘑菇分类的研究一直处于形态学分类研究阶段,运用分子标记对其属内种类进行系统发生分析的研究还是比较滞后的。国内外有大量关于运用系统发生学研究食用菌分类的报道[4-6]。武冬梅运用分子系统发生学探究新疆野生羊肚菌在物种水平上表现出较高的遗传多样性[7]。刘晓亮等通过分子系统发育学对东北大小兴安岭地区的红菇属进行物种多样性分析,共鉴定出23个红菇属物种,其中有3个种此前从未在我国报道过,分别是果香红菇、暗红红菇和诺尔亚红菇[8]。目前,利用草原蘑菇内转录间隔区(ITS)序列进行系统发生学探究还未见报道,本研究通过18S和ITS 2组通用引物扩增草原蘑菇基因组DNA片段,产物直接测序,结合BLAST、MEGA 6.0等生物信息学软件进行序列比对、遗传参数计算,以及构建系统进化树、分子钟等[9-11],以期为草原蘑菇的鉴定、进化分类及遗传育种方面的深入研究提供数据参考。
核糖体DNA(rDNA)中既包含保守的编码区18S、5.8S和28S,适合种以下水平的研究[12-13],又包含进化速率较快的非编码区,内转录间隔区1内转录间隔区和2(ITS1和ITS2)以及非转录区,承受的进化选择压力较小,进化速度较快,适合属以下水平的研究[14-15]。因此,常选用18S和ITS通用引物作分子标记研究不同分类地位的物种间关系[16]。这在食用菌分子鉴定及系统发育中已有大量应用[17],钱雪婷等基于ITS序列分析秦巴山区黑木耳菌株的遗传多样性,为今后秦巴山区黑木耳菌株的遗传多样性及优良品种的遗传育种奠定了理论基础[18]。戴淑娟等对采自新加坡的多孔菌基于ITS序列的系统发育分析结果表明,该新种属于变色卧孔菌属分支的一个明确的支系[19]。刘丽娜等利用ITS序列分析一个子实体菌盖为酒粉红色的金针菇新变型,结果表明,该变型同一子实体具有5种不同的ITS序列,表明金针菇存在丰富的种内遗传多样性[20]。本研究采用18S和ITS 2组通用引物扩增产物直接测序,对测序结果进行比对,2组测序结果互相印证补充,可使结果更加可靠。
草原蘑菇是内蒙古锡林郭勒草原产出的上品山珍,具有较高的营养价值和药用价值,是珍稀的食药用菌[21-22]。草原蘑菇主要品种有白蘑、黑蘑、香杏蘑、鸡爪蘑、珍珠蘑等,被誉为“草原明珠”“口蘑之王”等[23]。草原蘑菇以草-牧-菌循环的独特生长方式维系着大草原的生态平衡。近年来,草原气候干旱、退化日益严重,载畜量增加,加之人们过度采食,使这一珍贵的野生草原蘑菇自然生存条件愈加恶化,甚至濒临灭绝,其资源保育、驯化及利用研究任务艰巨,而且目前市场上的草原蘑菇种类混乱,以次充好、胡乱命名等现象时有发生,令人真伪难辨[24]。本研究从本质上对内蒙古中部草原地区草原蘑菇品系进行鉴定,通过形态学与分子标记相结合,研究各菌株之间的亲缘关系和分化时间,以期为后续草原蘑菇的种质资源创新和利用、草原蘑菇品种选育和遗传改良提供理论依据。
1 材料与方法
1.1 供试材料
草原白蘑和草原黑蘑采自内蒙古锡林郭勒盟白音锡勒草原地区,草原白蘑编号为草原白蘑74~89号,包含蒙古口蘑、花脸香蘑、青腿子等;草原黑蘑编号为蒙白音1~73号,其中,1~37号为白色鳞片,38~73号为棕色鳞片。
1.2 DNA提取
1.2.1 子实体DNA提取 将供试子实体置于 65 ℃ 烘箱内8 h烘干,然后置于微量植物样品粉碎机内粉碎,分装于离心管内,做好标记以备DNA提取。称取适量子实体粉末用天根试剂盒提取DNA[25]。
1.2.2 菌丝体DNA提取 提取适量菌丝体,置于液氮中研磨后用天根试剂盒提取DNA。
1.3 PCR扩增
18S序列通用引物(18S上和18S下)碱基序列为18S上:GTAGTCATATGCTTGTCTC;18S下:TCCGCAGGTTCACCTACGGA。
ITS序列通用引物(ITS1和ITS4)碱基序列为ITS1:TCCGTAGGTGAACCTGCGG;ITS4:TCCTCCGCTTATTGATATGC。
PCR扩增采用20 μL反应体系,包括Mix 10 μL,引物各1 μL,DNA模板2 μL,ddH2O 6 μL。PCR反应程序:94 ℃预变性3 min;94 ℃变性40 s,45 ℃退火1 min,72 ℃延伸1 min,30个循环;72 ℃继续延伸10 min,4 ℃保存。电泳条件:1.2%琼脂糖凝胶,电压为100 V。
用琼脂糖凝胶电泳检测PCR产物,检测合格后送英潍捷基(上海)贸易有限公司测序。
1.4 序列比对、序列的数据分析及系统进化分析
将测序结果在NCBI中进行BLAST比对,根据比对序列结果及形态特征,确定食用菌名称和相关菌种信息。选择羊肚菌为外类群,进行系统发育分析。利用MEGA 6.0软件进行保守位点、变异位点、简约信息位点、单一位点、简并态密码子、CpG岛、覆盖率、种内种间遗传距离、分子钟、邻接(neighbor-joining method,简称NJ)法构建系统发育树,以自展法(Bootstrap 1 000次重复)检验各分支的置信度。
2 结果与分析
将采集到的草原蘑菇按照子實体颜色、大小、形状等特征进行形态学分类[26],然后通过ITS序列扩增产物,比较序列间的差异,来建立群体之间的亲缘、进化关系,从而从本质上进一步鉴定[27]。
2.1 形态描述
2.1.1 蒙白音系列 草原黑蘑别称白鳞蘑菇,属伞菌目蘑菇科蘑菇属。蒙白音系列子实体为白色和棕色2类,菌盖直径为6~15 cm,半球形至平展,中部鳞片厚而大,龟裂明显,有时留有菌幕残片。菌裙初期为白色,渐变为粉红色至黑褐色。菌褶初期为肉色,密,离生,不等长,随着子实体生长发育成熟变为褐色至黑褐色。菌柄实心白色近圆柱形,菌柄长5.0~7.5 cm,粗0.7~7.0 cm。孢子呈卵圆形、椭圆形至长椭圆形,孢子印为黑色、表面光滑,孢子大小为(5~10) μm×(4~7) μm。
2.1.2 白蘑系列 蒙古口蘑属伞菌目口蘑科口蘑属或白丽伞属,别称白蘑、口蘑、珍珠菇、查干蘑等。子实体通体纯白色,初期半球形至成熟期菌盖平展呈伞形,成熟子实体菌盖直径为5~17 cm,菌肉厚硬为白色,初期菌盖边缘内卷;菌褶白色,稠密,弯生,不等长;菌柄粗壮,白色内实,长3.5~7.0 cm,粗 1.5~4.6 cm,柄基部稍大。担孢子呈肾形或椭圆形,孢子印为白色,表面光滑,孢子大小为(4~6) μm×(6~9) μm。
花脸香蘑属伞菌目口蘑科香蘑属,别称白花脸。子实体菌盖为白色,初期扁半球形至成熟期菌盖平展边缘上翘呈斗笠状,边缘水渍状乌白色,成熟子实体菌盖直径为4.0~8.0 cm,菌肉较厚白色;菌褶白色,直生或弯生,稍稀,不等长;菌柄中粗,白色,长3.5~7.0 cm,粗0.5~2.0 cm,内实,靠近基部常弯生。孢子呈椭圆形至近卵圆形,孢子印为白色,具粗糙麻点,孢子大小为(3~5) μm×(6~10) μm。
青腿子属伞菌目口蘑科香蘑属。子实体菌盖纯白色,初期半球形至成熟期菌盖平展边缘上翘,成熟子实体菌盖直径为10~22 cm。菌肉较厚白色,初期边缘内卷;菌褶白色,稠密,弯生,不等长;菌柄中粗,白色,长3.5~7.0 cm,粗0.8~3.0 cm,内实,基部稍大,柄由下向上青色光泽逐渐变深。孢子呈椭圆形至近卵圆形,孢子印为白色,具粗糙麻点,孢子大小为(3~5) μm×(6~10) μm。
2.2 草原蘑菇序列分析
利用MEGA 6.0軟件统计89株草原蘑菇ITS基因序列,保守位点(conserve)数为275/544,变异位点(variable)数为268/544,简约信息位点(parsim-infor)数为180/544,单一位点(singleton)数为88/544,简并位点zore-fold为319/544、tow-fold为62/544、four-fold为40/544,CpG为30/544。89株草原蘑菇ITS序列G+C碱基含量分析结果见图1,黑蘑G+C碱基平均含量为42.73%,白蘑G+C碱基平均含量为4114%,G+C的含量明显低于A+T的含量,白蘑G+C的含量低于黑蘑G+C的含量。
2.3 遗传距离、亲缘关系及分化时间分析
ITS基因序列遗传距离计算结果见表1,属内遗传距离较小而属间遗传距离较大,黑蘑和白蘑种间遗传距离为0~0.45,其中黑蘑种内遗传距离为0~0.05,白蘑种内遗传距离为0~0.24。说明黑蘑之间进化距离较近,白蘑之间进化种类具有多样性(其中没有标出的编号遗传距离均为0)。
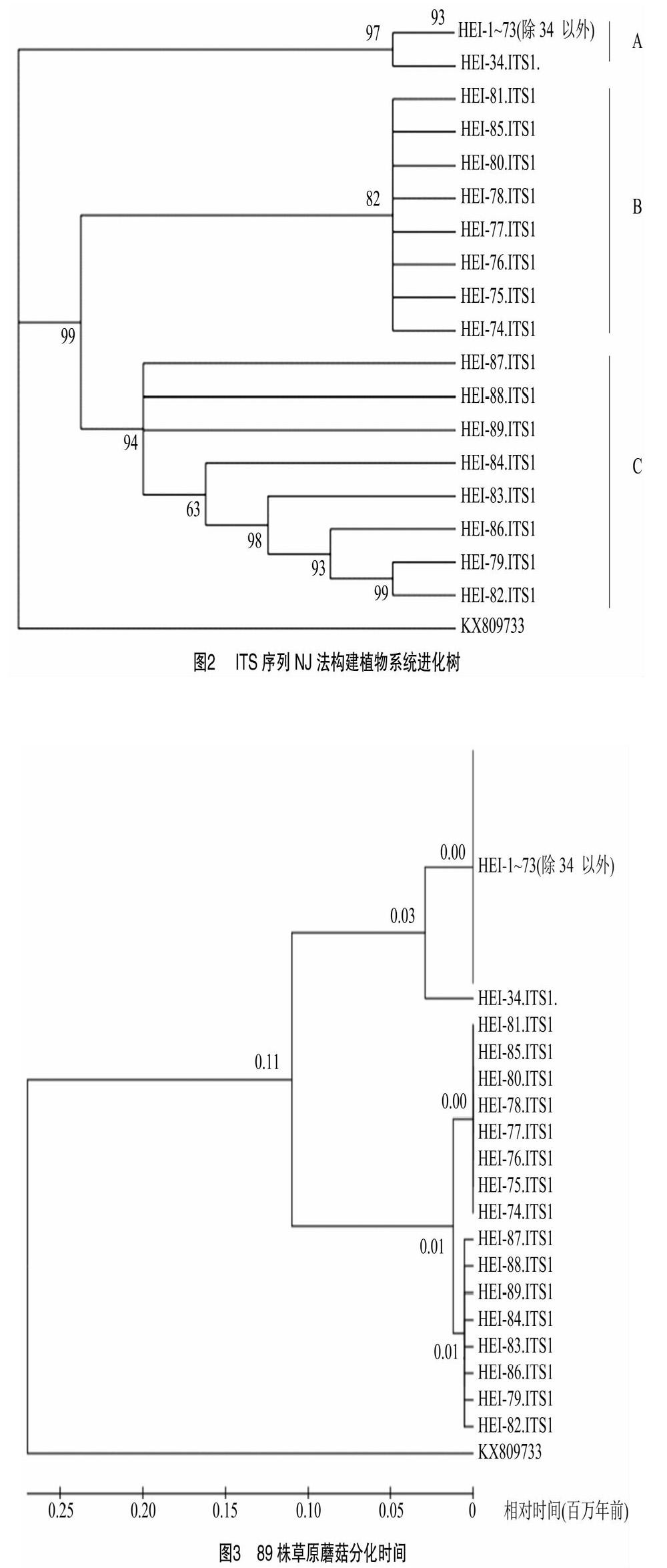
以测得ITS序列为依据,采用NJ法构建系统进化树,用羊肚菌(KX809733)作为外类群(图2),并用最大似然估计(maximum likelihood)法验证分类结果的准确性,结果表明,2种方法构建的进化树关系基本一致,89株草原蘑菇可分为3组,A组为黑蘑,B、C组为白蘑,白蘑遗传多样性较丰富,黑蘑则相对较少,34号与1~73号(除34号外)有相对差异外,其他基本相同,与遗传距离矩阵相符。
利用ITS基因序列,计算草原蘑菇之间的分化时间。结果(图3)表明,黑蘑和白蘑的分化时间为11万年前,其中黑蘑中34号与其他黑蘑分化时间为3万年前,白蘑中B类群和C类群分化时间为1万年前。不同属内草原蘑菇分化时间较长,同一属内的草原蘑菇之间分化时间较小。
3 结论
ITS序列己被广泛应用于食用菌的科、属级水平类群的亲缘关系系统发育分析[28-29]。本研究将形态学分类与ITS序列相结合,用于内蒙古中部地区的89株草原蘑菇亲缘关系研究,发现属内遗传距离较小亲缘关系较近,而属间遗传距离较大。通过计算分化时间,白蘑和黑蘑分化于11万年前,其中34号黑蘑与其他黑蘑分化于3万年前,白蘑中B组与C组于1万年前分化。采用NJ法构建系统进化树,用最大似然估计法验证分类结果,最终2种方法构建的进化树关系基本一致。从构建的进化树可以看出,89株草原蘑菇可分为3组,黑蘑为1组,黑蘑遗传较稳定,白蘑分为2组,白蘑的遗传多样性较丰富,很有可能是由于当地的气候条件引起的,如干旱、紫外线过强、昼夜温差过大导致变异。针对草原白蘑的遗传多样性状况,可以认为内蒙古中部地区草原白蘑具有较高的进化潜力和适应能力。内蒙古属干旱、半干旱地区,常年干旱少雨,近些年尤为严重,加之滥采滥挖造成野生草原蘑菇濒临灭绝,本研究在物种多样性的研究基础上结合ITS片段分析,对野生草原蘑菇的资源保护及如何利用分类结果进行人工驯化栽培具有指导意义。
草原蘑菇具有一定的遗传多样性,当地牧民多以形态学来定义其种类,即外观层面,则造成定义混乱,存在一定的偏差,因此,仅依据传统的形态特征来进行草原蘑菇的分类方法受到了越来越大的挑战,不能得到分子数据的支持。通过基于ITS分子层面测序,发现至少存在3个属,具有属间多态性,那么若要具体到种间分类将更加混乱,所以还需要新的分子标记方法进行进一步的分类、探索,充分地挖掘草原蘑菇资源。
参考文献:
[1]吴 芳. 木耳属的分类与系统发育研究[D]. 北京:北京林业大学,2016.
[2]Wu F,Yuan Y,Malysheva VF,et al.Species clarif canon of the most important and cultivated Auricularia mushroom wHeimuer:evidence from morphological and molecular data[J]. Phytotaxa,2014,186(5):241-253.
[3]渠志臻. 蒙古口蘑菌丝体营养生理特性及培养条件研究[D]. 呼和浩特:内蒙古农业大学,2010.
[4]Yang K,Tian Z X,Chen C H,et al.Genome sequencing of adzuki bean(Vigna angularis) provides insight into high starch and low fat accumula-tion and domestication[J]. Proc Natl Acad Sci USA,2015,112(43):13213-13218.
[5]Moncalvo J M,Lutzoni F M,Rehner S A,et al.Phylogenetic relationships ofagaric fungi based on nuclear large subunit ribosomal DNA sequences[J]. Systematic Biology,2000,49(2):278-305.
[6]邹 莉,孙婷婷,王旭彤,等. 松杉灵芝分离纯化及ITS分子鉴[J]. 江苏农业科学,2016,44(8):278-280.
[7]武冬梅. 新疆野生羊肚菌分子系统学和遗传多样性研究[D]. 乌鲁木齐:新疆大学,2015.
[8]刘晓亮,图力古尔,王向华. 东北大小兴安岭地区的红菇属物种多样性[J]. 菌物学报,2017,36(10):1355-1368.
[9]王 彤,刘 静,郭 月,等. 基于matK基因和ITS序列的江苏地方豆类植物的亲缘关系研究[J]. 南京农业大学学报,2017,40(5):795-803.
[10]谭河林,许欣颖,付立曼,等. 甘藍型油菜及其亲本物种甲基化酶Ⅰ基因的克隆及表达模式[J]. 作物学报,2015,41(3):405-413.
[11]李建伏. 基于DNA序列的进化树构建算法的研究[D]. 哈尔滨:哈尔滨工业大学,2008.
[12]姜芳燕,黄 海,杨 宁,等. 海南几种大型绿藻18S rRNA基因的克隆与序列分析[J]. 热带农业科学,2017,37(6):49-54.
[13]何 文,张 静,黄智林,等. 基于ITS序列对栽培中国樱桃遗传多样性及其群体遗传结构的分析[J]. 西北植物学报,2014,34(4):0463-0472.
[14]刘 悦,汤 欢,李西文,等. 基于ITS序列的伞菌类药用食用真菌的DNA条形码鉴定研究[J]. 世界中医药,2016,11(5):791-795.
[15]Schoch C L,Seifert K A,Huhndorf S,et al.Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for fungi[J]. Proceedings of the National Academy of Sciences of the United States of America,2012,109(16):4241-6246.
[16]杨 光,俞菊华,徐 跑,等. 利用18S和ITS序列揭示8种鲇形目鱼类的系统发育[J]. 动物学杂志,2010,45(4):110-117.
[17]沈 洪,陈明杰,赵永昌,等. 云南羊肚菌rDNA的ITS序列与亲缘关系分析[J]. 食用菌学报,2007,14(2):15-18.
[18]钱雪婷,陈文强,邓百万,等. 基于ITS序列分析秦巴山区黑木耳菌株的遗传多样性[J]. 北方园艺,2016(12):147-151.
[19]戴淑娟,戴玉成. 采自东南亚的变色卧孔菌一新种[J]. 菌物学报,2018,37(2):145-150.
[20]刘丽娜,张 平. 金针菇一个新变型的形态学鉴定及ITS序列分析[J]. 菌物研究,2016,14(1):38-42,51.
[21]吴晓彤. 蒙古口蘑分子鉴定及其菌丝体液体发酵技术与应用[D]. 呼和浩特:内蒙古大学,2014.
[22]孟建宇,宋馨宇. 蒙古口蘑子实体凝集素性质初步研究[J]. 生物技术通报,2012(2):188-192.
[23]刘晓婷,郭九峰,王淑妍,等. 蒙古口蘑担孢子萌发及初生菌丝生物学特性研究[J]. 北方园艺,2016(16):136-141.
[24]解亚杰,孙国琴,庞 杰,等. 草原野生食用菌蒙古口蘑的研究进展[J]. 食药用菌,2017,25(1):28-33.
[25]刘晓婷,郭九峰,王淑妍,等. 用rDNA-ITS方法鉴别内蒙古多种野生食用菌[J]. 食药用菌,2015,23(5):301-306.
[26]卯晓岚. 中国大型真菌[M]. 郑州:河南科技出版社,2000.
[27]郑怡鸿. 南瓜属植物的分子系统发育及分子钟研究[D]. 武汉:武汉大学,2011.
[28]张 明. 华南地区牛肝菌科分子系统学及中国金牛肝属分类学研究[D]. 广州:华南理工大学,2016.
[29]韩美玲. 中国拟层孔菌属及近缘属的分类与系统发育研究[D]. 北京:北京林业大学,2016.段燕燕,张换换,王 强,等. 不同施氮量对春小麦-青贮玉米土壤氨挥发的影响[J].
