
化石
2020-07-25 01:09
生物进化 2020年2期
叠层石最后的黄金时期
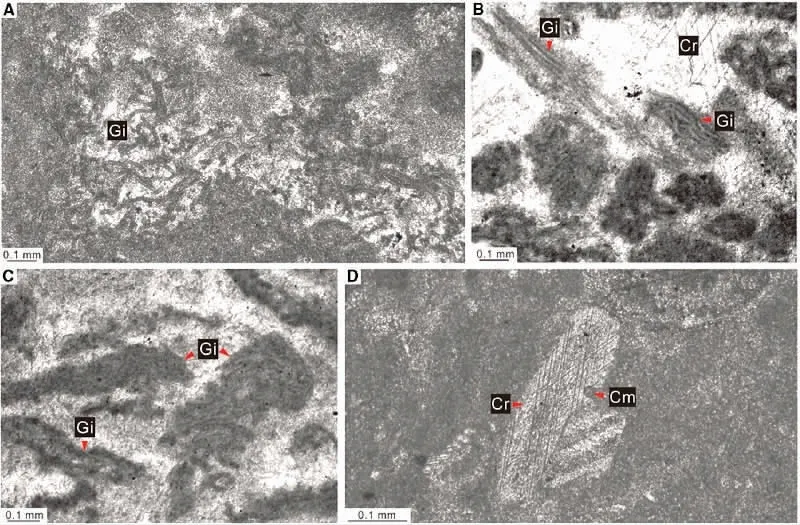
在距今4.85-4.43 亿年前的奥陶纪,浅海生态系统发生了一次地质历史上最重要的变革:后生动物主导的生物礁大规模地替代微生物礁。早先的研究认为,这一关键转变以古生代珊瑚-层孔海绵礁的兴起为标志,主要发生在中奥陶世。然而,近年来越来越多的证据显示,早奥陶世的苔藓虫、海绵类(例如石海绵、角质海绵和高钙化海绵等)和珊瑚型底栖动物便已在世界各地的陆表海环境参与造礁,甚至形成类似现代珊瑚礁的钙质礁格架结构。这些早期的后生动物礁大多出现在奥陶纪的第二阶(弗洛阶),而在早奥陶世特马豆克阶(第一阶)微生物礁仍然非常繁盛。近期,研究者通过对安徽南部下奥陶统地层进行多次野外考察,并通过显微薄片开展了深入研究。发现在较近岸的北贡剖面,叠层石礁呈平缓的穹隆状,微生物岩主体由纤维状钙质微生物(蓝细菌)构成,有柄棘皮类直接附着在微生物所形成的硬底上生长。而在相对远岸的马鞍山剖面,叠层石呈柱状或尖锥状,单个叠层石柱可高达1 米,同时缺乏棘皮动物等底栖固着生物。与这些碳酸岩台地边缘的叠层石形成对比,更远岸剖面斜坡相的大坞圲组因进入无光带而缺乏微生物礁。微生物岩的宏观和微相特征显示扬子台地东缘特马豆克阶仑山组叠层石的古地理分异主要受控于古水深。与扬子台地西部陆表海域相比,该研究聚焦的安徽南部案例显示出微生物礁主导的特点,底栖后生动物鲜见,不但说明后生动物礁替代微生物礁的过程具有强烈的地区差异,也印证了早奥陶之初确实是显生宙中微生物岩最丰富的时期之一,代表了叠层石最后的黄金时期。(Facies 2020,66:12)
奥陶纪末赫南特贝腕足动物群研究系列进展
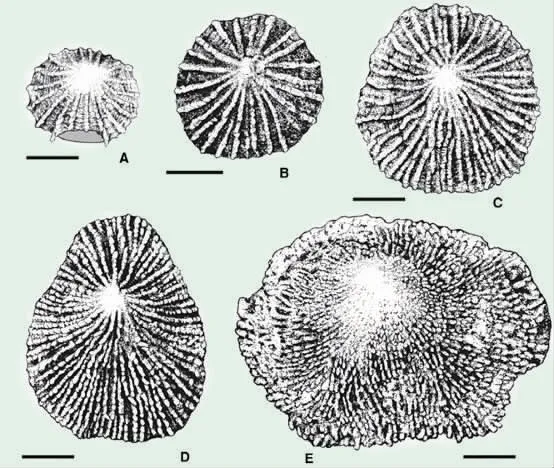
奥陶纪末大灭绝两幕事件之间的赫南特贝动物群广布全球,然而它在滇缅马地块的报道却异常稀少。这个地块包含现今的缅甸、马来西亚半岛、泰国西部、苏门答腊及中国云南西部的部分区域,在国际上处于重要的地理位置。近期,研究者对缅甸曼德勒地区、云南西部芒市地区,以及华南贵州湄潭等地区,同为赫南特贝动物群进行了大量的采集及研究,共识别出缅甸曼德勒地区腕足动物23 属23 种、滇西地区22属22 种、黔北地区13 属13 种,包括首次在缅甸发现赫南特贝,对一些关键分子的系统分类提出新的论点,建立新属奇异髑髅贝(Xenocra nia),厘定赫南特贝动物群中最常见属种的同物异名关系。通过对滇缅马和华南两个块体赫南特贝动物群重要成员、常见和共同分子的比较,显示在晚奥陶世晚期,其古地理位置与华南板块、拉萨地体相距并不遥远,都处于地球东半部热带浅水海域里。缅甸曼德勒地区和黔北湄潭地区的赫南特贝动物群分别属于典型的浅水及较深水动物群,它们在多样性、丰度及体型大小上均存在着明显差异。从浅水到深水的群落生态演替,反映了全球冰期气候和海洋环境的复杂性及其演变特征。(Palaeoworld 2020,29:1-30;Palaeoworld 2020, 29: 31-46;Journal of Pa leontology 2020, 94:239-254;Lethaia 2020, doi.org/10.1111/LET.12360;Papers in Palaeontology 20195,3:521-535.)
6 亿年前瓮安旋孔虫的身世之谜
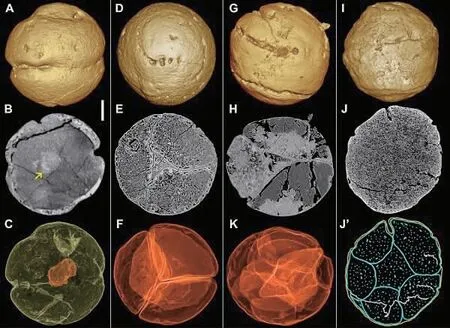
动物是如何起源的呢?这一直是困扰古生物学家和演化生物学家的前沿科学问题。探索这一问题的答案有两种途径:一种是演化发育生物学理论推测,即从分子的层面对比动物及其单细胞亲近的发育过程,在分子生物学构建的动物总界时间树的基础上,推测动物祖先的生物学特征,并重建这些特征的系统发生过程,估算其出现的时间;而另一途径则是古生物学实证研究,即通过追溯最早的动物及其近亲的化石记录,重建动物起源的过程并卡定动物起源时间的上限。6 亿年前的瓮安生物群中胚胎状化石的生物学属性一直充满争议,但近几年的研究进展对这些化石的认识逐渐趋向了一致:它们当中一些类群可能代表了最早的动物(包括干群动物),而另一些可能代表了动物的单细胞亲近。瓮安生物群中有一类胚胎状化石——瓮安旋孔虫,表面上长有奇怪的螺旋状线圈,这到底是一种什么样的生物?长期以来学界有各种假说,但均没有足够的证据证实。近期,研究者采用高分辨率显微CT 和同步辐射显微断层成像技术,扫描了数百个瓮安旋孔虫化石标本,发现,瓮安旋孔虫具有从单细胞期通过不增长的细胞分裂一直发育到成千上万细胞阶段的发育过程。这一发育过程表明瓮安旋孔虫不可能是单细胞的有孔虫,而可能属于动物总界的一员,为理解动物起源提供了重要线索。(Sci.Adv.2020,6,eabb0083)
埃迪卡拉纪宏体化石埋藏学和古生态学
全世界范围内广泛分布的埃迪卡拉生物群是"寒武纪生命大爆发"前夕最为独特的宏体化石生物群。其时代为埃迪卡拉纪中-晚期(5.79 亿-5.41 亿年前),代表了一些最早的具复杂形态的宏体多细胞真核生物组合,其中大多化石是以铸模或铸型的方式保存在粗碎屑岩中。近来的研究表明,部分埃迪卡拉型化石也可以以铸模或铸型的方式保存在碳酸盐岩中,或者以碳质压膜的形式保存在黑色页岩中,并且可以延伸到埃迪卡拉纪早期(6.35 亿-5.79亿年前),但仅有极少数类型化石可以同时出现在不同的埋藏窗口中。近期,研究者在华南的蓝田生物群和石板滩生物群,以及澳大利亚的埃迪卡拉生物群中,发现了一类共同的化石分子——扇形藻(Flabellophyton)。扇形藻是蓝田生物群(6.35 亿-5.79 亿年前)中的代表分子,以碳质压膜的形式保存于埃迪卡拉纪蓝田组二段的黑色页岩中;随后又被发现于澳大利亚南部埃迪卡拉纪中-晚期的埃迪卡拉生物群(5.60 亿-5.50 亿年前)中,并以铸模或铸型的方式保存在埃迪卡拉砂岩段;华南埃迪卡拉纪晚期的石板滩生物群(5.51 亿-5.39亿年前)中也含有这类化石,以铸模或铸型的方式保存在灯影组石板滩灰岩段中。基于蓝田生物群和石板滩生物群中的材料,研究者识别出了三个不同的形态种:蓝田扇形藻、标准扇形藻和钝锥扇形藻。依据形态学特征和解剖学结构,将其恢复为一种具有基部固着器、下部柄状结构、上部锥状主体和顶端开口的,营固着底栖、直立生长的疑难生物类型。这一发现不仅扩展了扇形藻的地层和时代分布,同时也使其成为目前唯一一种可以保存在三种不同埋藏窗口中的埃迪卡拉型化石。蓝田生物群扇形藻化石埋藏学研究表明有机质碳化、黄铁矿化和粘土矿物参与了化石的保存过程;石板滩生物群化石表明微生物席的参与和自生的方解石的充填有助于化石的保存;而埃迪卡拉生物群化石埋藏学研究认为自生的二氧化硅或者黄铁矿有助于微生物席形成"死亡假面",促进了化石的保存。扇形藻具有较强的环境适应性和生态迁徙/扩张的能力,从埃迪卡拉纪早期蓝田生物群代表的间歇性氧化的较深水安静泥质基底环境,扩展到埃迪卡拉纪中晚期的石板滩生物群代表的动态氧化的较浅水碳酸盐岩基底环境和埃迪卡拉生物群代表的氧化浅水砂质基底环境。蓝田生物群中的扇形藻形成了埃迪卡拉纪早期的较深水环境中的固着底栖、直立生长的生态群落,这一简单的垂向生态系统取代了之前的较为原始的微生物席状生态系统,并在埃迪卡拉纪中-晚期同其它埃迪卡拉型生物一起逐步迁徙/扩展到了整个海洋,为之后复杂生态系统的形成奠定了基础。(Gondwana Research 2020,84: 296-314)

我国第一具翼龙化石骨架——魏氏准噶尔翼龙头骨腭区研究进展
魏氏准噶尔翼龙是我国发现的第一具较完整的翼龙化石骨架,1964 年杨钟健研究命名并建立了准噶尔翼龙科,当时认为这是第一次在我国发现的翼龙类化石。实际上,早在1951 年,杨钟健和刘东生等在山东莱阳就发现少量破碎的可能属于翼龙的骨骼化石,后经研究证明这的确是我国最早发现的翼龙类化石。魏氏准噶尔翼龙产自我国新疆准噶尔盆地西缘的克拉玛依乌尔禾地区下白垩统吐谷鲁群的湖相沉积中。在乌尔禾地区进行的野外考察与发掘,发现了该地区富含白垩纪陆生脊椎动物化石,并将这一脊椎动物化石组合命名为"乌尔禾翼龙动物群",包含数量众多的翼龙及其他门类化石,其中有3 件较为完整的准噶尔翼龙头骨,以及一类比准噶尔翼龙体型更小的复齿湖翼龙。近十年来,研究者在乌尔禾和周边地区进行了新一轮的野外科考,发现了大量翼龙、恐龙等骨骼及足迹化石等。

准噶尔翼龙具有非常独特的头部形态特征,如上下颌前部锥状上翘无齿的吻端,在鼻眶前孔前缘开始发育并且一直向后延伸至枕区上部的头饰,小小的眼眶,锥状短圆的牙齿及齿列后部的附着牙齿的上颌骨膨大等特征,在翼龙家族中独树一帜,很容易与其他翼龙区分。由于翼龙飞行需要减轻重量,它们的骨骼细长中空,很难保存化石,即使保存也大多不完整。由于魏氏准噶尔翼龙头骨较完整,都呈三维立体保存,为翼龙的腭区的形态学研究提供了非常理想的材料。近期,研究者通过对多件魏氏准噶尔翼龙腭区每一块骨骼的详细形态观察和对比,并通过对全世界已知保存颚区的翼手龙类的17 个属种化石材料对比,从内鼻孔之后的腭面骨骼相互关系和结构分析,魏氏准噶尔翼龙的腭区特征与无齿的神龙翼龙类的成员相近。新研究发现,准噶尔翼龙上下颌在吻端外侧面,从第一颗牙齿开始,发育从咬合面镜像对称且分别向上下延伸的沟,这很可能是准噶尔翼龙吻端附着角质外套边缘的印痕。此外,其吻端上翘无齿的部分表面还发育不规则分布的大量细长小孔,可能为滋养孔,这些新的特征显示准噶尔翼龙吻端可能发育类似鸟类的角质喙。准噶尔翼龙的牙齿短粗圆钝,与其他大部分翼龙具有的细长尖锐的牙齿明显不同,成年个体的牙齿数量一般在上颌一侧发育14-15 颗,下颌11 枚左右,一些牙齿可以观察到明显的平行于腭面的磨蚀面。准噶尔翼龙所具有的上翘无齿具角质喙的吻部、短粗圆钝具磨蚀面的牙齿等形态,进一步佐证了其挖掘捕食泥沙中具有坚硬外壳的贝壳类等食物的生活习性。(Peer J 2020,8:e8741)
藏在牙齿里大灭绝之谜
六千五百万年前的大灭绝事件,重创了地球生态系统,但恐龙的后代——鸟类,是如何躲过的,一直是备受关注。近期,研究者通过同步辐射的高解析穿透式X-光显微镜,对小型非鸟类恐龙-包含伤齿龙、近鸟龙、小盗龙,以及古鸟类——包含今鸟类、反鸟类、会鸟和热河鸟等牙齿的微结构进行对比观察和研究,发现尽管简单的釉质层在早期古鸟类中都有保留,但牙釉质与牙本质之间的多孔罩牙本质层皆已消失,而多孔罩牙本质层被认为是肉食性恐龙牙齿中发育的、避免其在掠食过程中牙齿断裂的特殊避震保护结构。在古鸟类中,此次研究的一种小盗龙标本的罩牙本质层也已经消失。这意味着鸟类与部分亲缘关系相近的恐龙,其牙齿不再需要特殊的力学保护结构;间接证实了其在饮食习惯,如咬合力与掠食性方面,与肉食性恐龙产生了极大差异,通过食性转换避开了与肉食性恐龙对食物生态位的竞争,适应能力极大提高,度过最艰难的时刻。相较于古鸟类的普遍性植食或杂食化的演进趋势,虽然非鸟类恐龙的少数类群,如小盗龙,也发生了趋同演化,但仍难以避免灭绝的危机。(BMC Evolutionary Biology 2020,20:46)

中国已知最晚的离龙类——侏儒黑山龙
离龙类是一类已灭绝的水生/半水生爬行动物,最早出现于中侏罗世,至中新世消失。虽然全世界已报道的离龙种类并不多,仅有12 属25 种,但这个类群呈现出多样的形态特征,如新离龙类体型大、吻长颈短且下颞孔不闭合(如西莫多龙),而"非新离龙类"的体型小、吻短、下颞孔闭合,有长颈(如潜龙)和短颈(如满洲鳄)两种类型。亚洲,尤其是东亚,被认为是离类在早白垩世时辐射演化的关键地区。中国的离龙类绝大多数为辽西早白垩世热河生物群的成员,如满洲鳄和潜龙等。而在热河生物群之外离龙类的化石记录很少,只有来自内蒙下白垩统罗汉洞组的孙氏伊克召龙、新疆上侏罗统齐古组一些不确定的破碎颌骨材料,以及河北中上侏罗统髫髻山组的侏罗青龙。近期,研究者报道了一来自辽宁黑山县下白垩统沙海组煤层(1.2亿-1.0 亿年前)的离龙类新属种——侏儒黑山龙(Heishanosaurus pygmaeus),属名取自化石产地黑山县,种名缘于该属种与其他白垩纪离龙类相比体型较小的特点。沙海组地层产出了数量较多的脊椎动物化石,其中哺乳动物丰富,有多瘤齿兽类、三尖齿兽类、真兽类等。黑山龙标本表面只暴露了一个齿骨和一个颧骨,通过高精度CT 扫描在岩石内部发现了更多骨片,包括小半个头骨和一些零散骨片。研究者通过对扫描数据进行建模,再放大、镜像后3D 打印,最终恢复出侏儒黑山龙的大部分头骨结构。与侏罗纪的栉颌鳄和青龙相似,黑山龙吻短、下颞孔未闭合。根据下颌长度推测,侏儒黑山龙的体长约30 厘米,与栉颌鳄和潜龙的大小相当。在离龙类的系统发育树上," 非新离龙类" 满洲鳄、潜龙、戏水龙、复活龙等构成了异离龙类这一支;栉颌鳄、青龙和黑山龙位于新离龙类和异离龙类构成的支系之外,个体都比较小、形态特征原始,其中侏儒黑山龙在时代分布上最晚。(Journal of Systematic Palaeontology 2020,DOI:10.1080/14772019.2020.1749147)

华南地区志留纪中华盔甲鱼类研究新进展
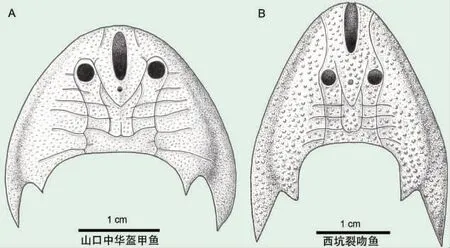
盔甲鱼类是无颌鱼类"甲胄鱼纲"下的一个类群,该类群最早出现于中国华南地区的志留系,在志留纪早期迎来了第一次适应辐射,出现了形形色色的盔甲鱼类群,称之为"张家界动物群"。中华盔甲鱼科属于该动物群中的"中华盔甲鱼-修水鱼组合",是真盔甲鱼目中最原始的类群,能够为解决脊椎动物从无颌向有颌转变过程中有颌类关键特征的演化提供化石证据。然而,由于该科中山口中华盔甲鱼的许多特征存在争议,且西坑"中华盔甲鱼"的大量形态学特征尤其是感觉管特征缺失,使得这一类群的单系性一直以来存在争议。近期,江西志留系发现的中华盔甲鱼科新材料填补了这些缺失特征的空白,基于深入的形态学研究,为西坑"中华盔甲鱼"建立了一新属——裂吻鱼属(Rumporostralis),并建立了新属下的一新种石盘裂吻鱼(R.shipanensis)。新属以中背孔后端位于眶孔之上,前端到达头甲吻缘并使吻缘裂开为主要鉴定特征。系统发育分析结果表明,裂吻鱼属与中华盔甲鱼属、安吉鱼属亲缘关系更近,它们构成一个单系类群——中华盔甲鱼科,从而确认了中华盔甲鱼科的单系性;而原来归到中华盔甲鱼科的曙鱼、煤山鱼属,则构成一个新的单系类群——曙鱼科,代表了真盔甲鱼目的最原始类群。中华盔甲鱼类的侧线系统在盔甲鱼类中非常特殊,展示出基干盔甲鱼类、真盔甲鱼目、多鳃鱼目和华南鱼目等多个类群的镶嵌特征。侧线系统是鱼类和两栖类独有的一类感觉器官,作为水流感受器,能够感知水的流向、压力、以及周围环境移动物体的情况,有助于动物调整姿态和运动方向。江西志留系的中华盔甲鱼类新材料保存了完好的侧线系统,深入的形态学研究表明中华盔甲鱼类的侧线系统由4条纵干管和3-8 条横向联络管相互交叉在头甲背面形成格栅状分布,可能反映了脊椎动物侧线系统的祖先状态。盔甲鱼类的生活环境长期以来一直被认为是近岸的滨海环境,而江西修水西坑组则指示了一种低盐度半咸水的近海环境,可能是由于大量淡水从附近河流注入而导致盐度降低,而含鱼层位的灰绿色泥质粉砂岩中含有大量的泥砾则指示了鱼化石经历了短距离的河流搬运。因此,该时期的盔甲鱼可能生活在其埋藏区域上游的淡水河流里。研究表明盔甲鱼的生态空间可能在志留纪早期就已经向 大 陆 盆 地 扩 展。(Peer J 2020,8:e9008)
南亚发现美洲豹的远古祖先
美洲豹亦称美洲虎,是新大陆现生最大的猫科动物。美洲豹现在分布在中南美洲,但分子系统发育学的研究表明美洲豹属于非洲大猫,与豹和狮组成一个支系,并且是这个支系最早分异的现生成员。这显示美洲豹应该起源于非洲,经过迁徙扩散到达北美和南美。事实上早在分子生物学的证据出现之前,欧洲学者就已经识别出欧洲更新世常见的中-大型豹属物种冈巴佐格豹在形态上与美洲豹十分接近,可能代表美洲豹在旧大陆的祖先。这一观点近年来得到多数学者的认同。然而在这种假说下,新大陆的美洲豹应该是旧大陆的冈巴佐格豹从欧洲迁徙到北美演化而成的,那唯一的迁徙路径是通过亚洲白令陆桥,最终到达北美洲。然而亚洲地区冈巴佐格豹的化石记录非常稀少,已知的两个有效记录来自于中亚和西亚。近期,研究者发表了关于南亚地区哈罗河流域早更新世化石点(古地磁测年142±0.35 万年)猫科材料的系统描述和对比。其中主要的化石材料可以归入冈巴佐格豹。该记录代表美洲豹演化支系在南亚地区的首次发现。化石材料包括一个不完整头骨和一些零散的下颌残断和牙齿。通过和冈巴佐格豹的其他材料对比,研究者确定南亚的冈巴佐格豹和西亚的格鲁吉亚冈巴佐格豹十分接近,很可能代表该种群进一步向南亚迁徙扩散的一个记录。通过对已知化石记录的梳理,研究者整理出了美洲豹的迁徙大致时间和路径:美洲豹最早的化石记录为南非的(190 万年前)。大概这一时期,美洲豹走出非洲,迁徙到了欧洲和西亚。美洲豹在欧洲长期存在,一直延续到中更新世晚期(大约30 万年前),而进入西亚的一支继续扩散,在100-14 0 万年前出现在中亚和南亚地区。北美的最早的化石记录来源于早更新世晚期约85 万年前西弗吉尼亚州的汉密尔顿洞穴。这条路线上目前最大的不确定处来自于东亚地区美洲豹支系的分布,有待将来的更多的研究来揭示。(Journal of Quaternary Science 2020,1-14)

世界上最大的亚洲足迹
亚洲足迹属是2005 年在日本下白垩统地层中发现并命名的一类恐龙足迹,模式种为跟垫亚洲足迹,其主要特点是具有明显清晰的蹠趾垫印迹,代表了兽脚类恐龙的足迹类型。近期,研究者在准噶尔盆地西北缘的乌尔禾及相邻地区,发现了多个层位保存的大量的翼龙、恐龙、鸟类和龟鳖类足迹组成的足迹化石群。这些兽脚类恐龙足迹保存在长约12 米宽约7 米,面积约80 平方米的灰绿色细粒砂岩层面上,地层近于水平。足迹化石层有上下两层,上部层面保存了6 个足迹,下部层面仅出露了7 个足迹(大部分被上层所覆盖),共组成3 条行迹和4 个孤立足迹。上部层面同时还保存了对称波痕和2条虫迹。通过对足迹、行迹特征的详细观察研究,识别出大小两种三趾型足迹类型。大型足迹在上部层面组成一条行迹和2 个孤立足迹,在下部层面也有两个孤立足迹,足迹呈趾行式、无拇趾印迹和尾迹、足迹为轴对称图形、V 字形、具有非常明显的蹠趾垫(脚跟)印迹,蹠趾垫呈近似椭圆形,足迹全长47-56 厘米,宽31-42 厘米。中型足迹在下部层面组成两条行迹,一些特征与大型三趾型足迹一致,但化石相对较小,比大型足迹小一半左右,足迹全长21-27 厘米,宽18-20 厘米。从足迹的大小和形态特征分析,所有足迹与实雷龙足迹科特征相似。通过对实雷龙足迹科已知的6 个属的正型标本和1 个属种未定成员的对比,两种足迹类型均归于亚洲足迹属,大型足迹属于亚洲足迹属一新种——牛氏亚洲足迹(Asianopodus niui),种名赠予足迹的主要发现者牛春旺先生。这一亚洲足迹新种比已知最大的亚洲足迹大约1.5 倍,是亚洲足迹属中最大的类型。中型足迹归入已知的跟垫亚洲足迹。根据经验公式,通过后足足长可以对造迹恐龙的臀高和体长进行估算。一般来说兽脚类恐龙的臀高约为后足足长的4 倍,体长约为臀高的2.63 倍。据此推算牛氏亚洲足迹造迹恐龙的臀高约为2.3 米,体长近6 米,属于大型兽脚类恐龙。跟垫亚洲足迹造迹恐龙的臀高约为1米,体长约为2.5 米。根据臀高和复步长等数值,也可根据经验公式进一步对恐龙的行走速度进行估算,其中牛氏亚洲足迹造迹恐龙的行走速度约为8 千米/小时,跟垫亚洲足迹造迹者的速度约为6.5 千米/小时。一般认为,复步长和臀高比值得出的相对复步长如果大于2,属于快速行走和奔跑状态,反之则属于缓慢行走状态,由于两类造迹恐龙的相对复步长都小于2,因此都处于漫步的状态。根据足迹层面保存的对称波痕(常见于双向水流的滨湖或滨海环境)、虫迹以及足迹化石的完整保存等特征,推断足迹化石是两类造迹恐龙漫步行走于滨湖环境形成的。(科学通报2020, 65)

5.14 亿年前新型多毛类环节动物
作为在寒武纪生命大爆发中出现的类群,环节动物(身体分节的蠕虫)例如蚯蚓、水蛭、沙蚕等在当今的陆地和海洋生态系统中起着举足轻重的作用。其中,多毛类动物是环节动物中物种最丰富、进化历史最悠久的一个类群。但环节动物在寒武纪早期地层中的化石记录非常罕见,这导致科学家在研究其起源和早期演化时缺乏有力的化石证据。近期,研究者在云南省东部富含软躯体化石的寒武纪地层中发现了一个新的、距今约5.14 亿年的多毛类动物,这是已知的现生环节动物支系的最早化石记录。研究者将这个寒武纪的新属种命名为"丹尼多毛虫(Dannychaeta tucolus),以纪念丹麦著名的环节动物专家Danny Eibye-Jacobsen。该动物具有一个典型的铁楸状头部和一对细长的触须,属于现生多毛类环节动物中的长手沙虫科。这是现生环节动物支系的最早化石记录,表明丹尼多毛虫和现生环节动物由同一个最近共同祖先演化而来,统称为环节动物冠群。现生环节动物在现代海洋中有各种生活模式,包括固着的滤食者或伏击捕食者。此前所知的寒武纪环节动物可能都是在海底爬行,而丹尼多毛虫则完全不同,是管居固着生活。现生的多毛类动物中有许多物种为管居或固定穴居,以此来躲避捕食者或在捕食过程中隐藏自己,通常被称为固着类环节动物。此前,现代生物学家通过系统发育基因组学研究,认为固着类环节动物应该代表环节动物进化树中一个比较古老的支系,但直到现在,固着生活方式一直未在早期的环节化石中被发现。寒武纪是早期生物多样性大爆发的一个关键时期,丹尼多毛虫的发现显示了"寒武纪生命大爆发"不仅仅是生物多样性,也是生态多样性剧增的一个重要进化事件。(Nature 2020,doi.org/10.1038/s41586-020-2384-8)

5.12 亿年前最古老的寄生关系
腕足动物是小型有壳的海洋动物,形似双壳类,现存约450 种,但是化石记录显示存在逾12000 种。腕足动物的化石记录可以追溯到寒武纪生命大爆发的初期。动物在寒武纪大爆发(约5.41 亿年前)后不久便需要与寄生虫作斗争。近期,研究者分析了在云南发现的乌龙箐新圆货贝的化石种群,测定年份约为5.12 亿年前。他们发现许多乌龙箐新圆货贝的壳外覆有一种管居生物,而覆有管居生物的乌龙箐新圆货贝体型明显较小,而且这些管状构造与腕足动物自身的摄食水流方向一致。因此,研究者认为这种管居生物通过偷窃宿主的食物,削弱宿主的健康(偷窃寄生现象)。这项发现标志着迄今已知在化石记录中发现的最古老的寄生关系。(Nature Communications 2020,11:2625)

猜你喜欢
娃娃乐园·综合智能(2022年10期)2022-10-18
奥秘(2022年7期)2022-08-09
广东第二课堂·小学(2022年5期)2022-06-02
奥秘(2022年5期)2022-05-31
奥秘(2022年4期)2022-04-26
红蜻蜓·低年级(2021年11期)2022-01-19
今日农业(2021年5期)2021-11-27
小哥白尼(野生动物)(2020年4期)2020-07-27
科学(2019年3期)2019-09-10
故事会(2009年5期)2009-05-21
