
利用基因组编辑技术构建拟南芥双突变体iqm5-1 flc
2020-07-30 14:01李惠梅范甜吕天晓周玉萍田长恩
科技视界 2020年14期
李惠梅 范甜 吕天晓 周玉萍 田长恩
摘 要
本研究组前期发现,拟南芥IQM5基因突变引起开花推迟,可能是提升了开花抑制因子FLC的转录本水平所致。然而 ,缺乏遗传学证据。本实验利用基因组编辑技术构建FLC的编辑载体,转化IQM5基因的T-DNA插入突变体iqm5-1,通过抗生素抗性筛选和测序得到双突变体iqm5-1 flc,为确定IQM5通过FLC调控开花提供了遗传材料。
关键词
IQM5;FLC;基因组编辑;双突变体;拟南芥
中图分类号: Q943.2 文献标识码: A
DOI:10.19694/j.cnki.issn2095-2457 . 2020 . 14 . 67
拟南芥的IQM家族共有6个成员, N-端有一段序列与豌豆重金属诱导蛋白同源, C-端则有一段与核糖体失活蛋白天花粉素同源的序列。IQM家族各成员可能在不同环境条件和不同发育阶段所起作用各不相同[1-2]。本研究组发现在幼苗、茎、莲座叶、花序叶和花蕾中,均存在 IQM5.1 和 IQM5.2 的两种转录本,不过,不同组织器官中二者的比例不尽相同[3]。在拟南芥中,已探明春化、光周期、赤霉素、年龄途经、自主以及温度等6条调控开花时间的遗传途径。开花抑制因子FLC能直接抑制包括FT、SOC1和FD在内的开花整合基因的表达[4]。本研究组发现,IQM5的T-DNA插入突变体iqm5-1和iqm5-2在长日照或短日照下均呈现晚花表型;FLC在野生型植株的茎、叶中均微弱表达,但在突变体iqm5-1和iqm5-2的茎、叶中表达量均显著增高[5]。因此,推断IQM5基因突变引起开花推迟,可能是提升了开花抑制因子FLC的转录本水平所致[5]。本研究旨在利用CRISPR/Cas9基因组编辑技术构建双突变体iqm5-1 flc,为确定IQM5通过FLC调控开花提供遗传材料。
1 实验材料与方法
1.1 实验材料
植物材料:iqm5-1(CSHL_GT18219)是Landsberg erecta(Ler)生态型的IQM5 T-DNA 插入突变体,来源于Cold Spring Harbor Laboratory, Cold Spring Harbor, USA。载体:pYLsgRNA-AtU3d、pYLsgRNA-AtU3b、pYLCRISPR/ Cas9-Pubi-H菌种由华南农业大学刘耀光院士友好赠送。菌种:大肠杆菌E.coli DH5α及农杆菌GV3101菌株均由本实验室保藏。试剂:Bsa I核酸内切酶购于NEB公司,其他试剂盒与工具酶均购自TAKARA生物工程有限公司;MS盐(Murashige&Skoog )购自Sigma公司;潮霉素、Yeast Extract、Typtone和Agar M等为生工生物工程(上海)有限公司产品 。测序及引物合成:由生工生物工程(上海)有限公司及完成。引物序列 (均为5′→3′):SP-L1:GCGGTGTCATCTATGTTACTAG,SP-R:CGACATAGATGCAATAACTTCG;3D-FLC-F:GTCACCTTCTCCAAACGTCGCAA,3D-FLC-R:AAACTTGCGACGTTTGGAGAAGG;3B-FLC-F:GTCAGGAGAAGCTGTAGAGCTTGC,3B-FLC-R:AAACGCAAGCTCTACAGCTTCTCC;Cas9-FLC-F:GCGGTACACGTGGCAATCTT,Cas9-FLC-R:ACAACATCGAGCACGCATCA。
1.2 实验方法
质粒提取、PCR、酶切、DNA片段回收与纯化方法参照对应试剂盒说明书进行。载体构建参考CRISPR/Cas9基因编辑技术方法[6]进行:在FLC基因中设计2对靶点接头3D-FLC-F/R和3B-FLC-F/R,分别退火连成小片段,重组插入经BsaI线性化的载体pYLsgRNA-AtU3d和pYLsgRNA-AtU3b中;经gRNA表达盒扩增后重组入经BsaI线性化的双元载体pYLCRISPR/Cas9-Pubi-H;热激转化大肠杆菌,经菌落PCR鉴定及测序验证后提取质粒转化农杆菌,再利用农杆菌蘸花法转化突变体iqm5-1;收获成熟种子,经潮霉素抗性筛选得到T0代幼苗,移至土壤培養;在靶点上下游设计引物Cas9-FLC-F/R进行PCR扩增和测序,筛选基因编辑成功株系,在后代中继续筛选纯合转化植株。
2 结果与分析
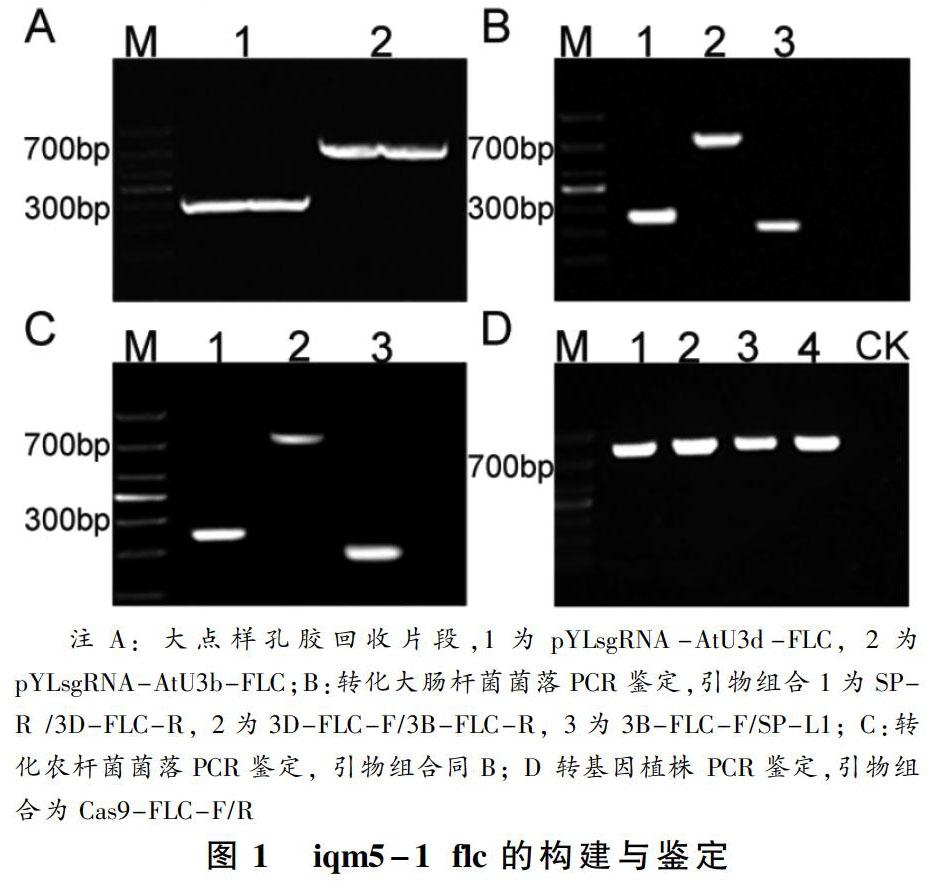
本研究利用CRISPR/Cas9对拟南芥iqm5-1的FLC基因进行编辑。首先,分别将靶点接头3D-FLC-F/R和3B-FLC-F/R退火连成小片段,插入pYLsgRNA-AtU3d-FLC和pYLsgRNA-AtU3b-FLC,经gRNA表达盒扩增,得到预期产物(图1-A)。其次,将扩增产物插入线性化的pYLCRISPR/Cas9-Pubi-H后转化大肠杆菌,挑选抗性菌落进行PCR鉴定,获得阳性菌落(图1-B);进一步,从阳性菌落提取质粒,转化农杆菌,菌落PCR鉴定结果符合预期(图1-C);最后,利用农杆菌转化拟南芥突变体iqm5-1,收获种子经潮霉素抗性筛选获得候选植株,对其进行PCR鉴定获得预期产物(图1-D),外送测序获得预期的基因组编辑植株。
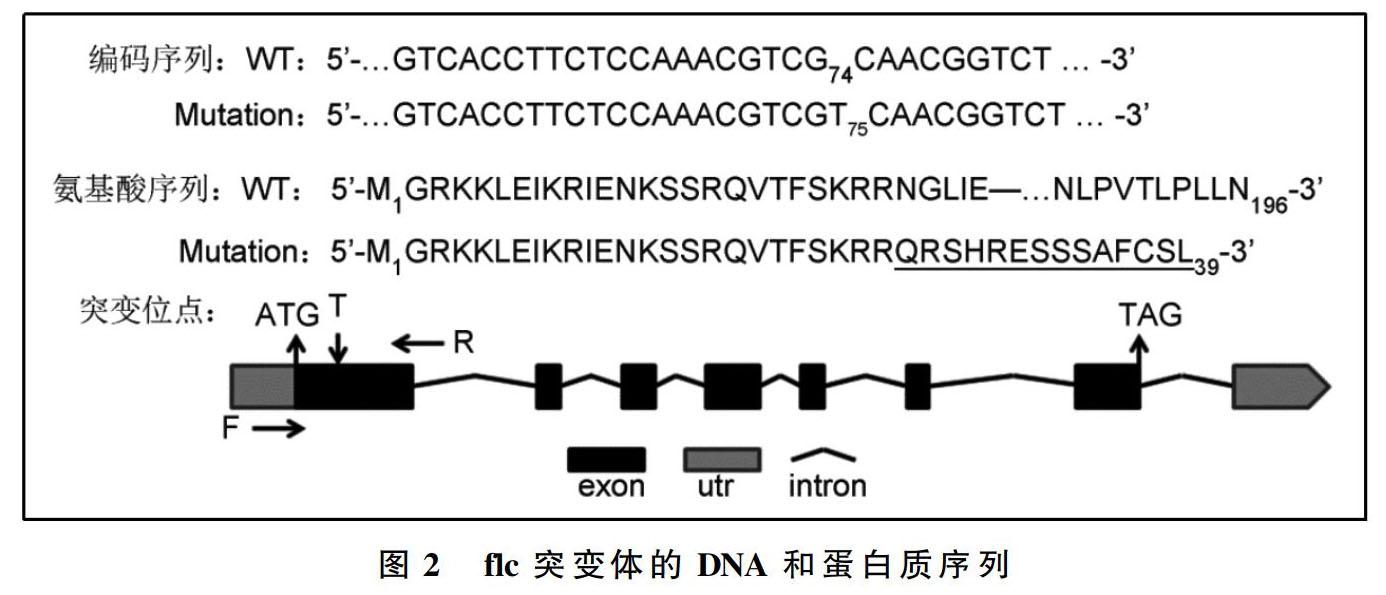
将上述候选编辑植株后代继续进行抗性筛选和序列分析,成功获得1株纯合双突变体iqm5-1 flc。在该双突变体中,iqm5-1 已经报道[5];flc的突变如图2所示:从FLC起始密码子ATG的A开始,在第74与第75位碱基间插入1个碱基(T),该碱基的插入导致其后序列发生移码突变,并提前终止,产生仅有39个氨基酸残基的突变蛋白,而野生型FLC有196个氨基酸残基。
3 讨论
本研究组前期发现,IQM5 的2个T-DNA插入突变体iqm5-1和iqm5-2在长日照或短日照条件下均呈现晚花表型,且其茎、叶中FLC的表达量均显著增高。因此,IQM5基因突变可能通过提升开花抑制因子FLC的转录本水平而实现[5]。不过,缺乏遗传材料。本研究利用CRISPR/Cas9基因编辑技术方法,成功编辑了突变体iqm5-1中的FLC基因,筛选得到双突变体iqm5-1 flc,为确定IQM5通过FLC调控开花提供了遗传材料。
参考文献
[1] 田长恩, 周玉萍. 植物具IQ基序的钙调素结合蛋白的研究进展[J]. 植物学报, 2013,48 (4): 447-460.
[2] ZhouYP, Chen YZ, Yamamoto KT, Duan J, Tian CE. Sequence and expression analysis of the Arabidopsis IQM family[J]. Acta Physiol Plant, 2010,32: 191–198.
[3] 弓路平, 萧文慧, 周玉萍,黄小玲,田长恩. 拟南芥 IQM5.2 的克隆、表达及生物信息学分析[J].生物技术通报,2016,32(5):69-74.
[4]張艺能, 周玉萍, 陈琼华,黄小玲,田长恩. 拟南芥开花时间调控的分子基础[J]. 植物学报,2014, 49 (4): 469–482.
[5] Gong LP, Cheng JZ, Zhou YP, Huang XL, Tian CE. Disruption of IQM5 delays flowering possibly through modulating the juvenile-to-adult transition[J].Acta Physiol Plant,2017,39:21.1-10.
[6] Ma XL, Liu YG. CRISPR/Cas9-based multiplex genome editing in monocot and dicot plants[J]. Curr Protoc Mol Biol , 2016, 115:31.6.1-31.6.21.
猜你喜欢
空间科学学报(2021年1期)2021-05-22
生命科学研究(2018年1期)2018-05-29
环境保护与循环经济(2017年5期)2018-01-22
上海农业学报(2017年3期)2017-04-10
安徽医科大学学报(2016年12期)2017-01-15
山东农业工程学院学报(2016年6期)2016-12-01
中国果菜(2016年9期)2016-03-01
天津医科大学学报(2015年2期)2015-12-22
中国蔬菜(2015年9期)2015-12-21
山东医药(2015年40期)2015-02-28
