
腐乳源酵母分离鉴定及其极性脂质分析
2020-08-07 03:51陈思锴林盛杰
食品与机械 2020年7期
陈思锴 林盛杰 李 理
(1. 华南理工大学,广东 广州 510642;2. 广州市白云联佳精细化工厂,广东 广州 510445)
脂质是细胞膜的主要成分,构成了细胞边界,可以储存能量并参与细胞的信号传导过程[1-2]。对于哺乳动物而言,酵母菌的脂质组成相对简单,脂质代谢相关基因所占比例相对较高,分子遗传分析手段较为成熟,使得酵母在脂质研究中备受关注[1]。酿酒酵母是研究真核生物脂质调控机制的主要模型生物[3],耶氏解脂酵母能代谢多种底物并转化成生物脂质[4],斯达氏油脂酵母[5]等产油酵母侧重于对甘油酯的代谢分析,酵母脂质组学研究缺乏对其他酯类物质和酰胺类化合物的关注。据报道[6-7],马克斯克鲁维酵母和库德里阿兹威毕赤酵母可应用于生产生物脂质,但试验所分离的大部分菌种目前未见其脂质方面的研究。
生物样品中脂质种类繁多、基质复杂,故脂质组学分析需要借助先进的分离技术和检测手段[8]。近年来,基于质谱的脂质组学研究提高了对细胞脂质生物学的认识,但脂质的定性定量问题仍是制约脂质组学发展的主要障碍。超高效液相串联质谱法(UHPLC-MS/MS)能将液相的强分离效果和质谱的高灵敏度、高分辨率以及快速检测的优势良好地结合起来[9],能对生物体内脂质种类和含量进行监测,是目前脂质组学研究的一种重要手段。研究拟从腐乳厂的酸浆水和毛胚中分离鉴定出多个酵母菌株,着重于酵母的产酶特性分析,并利用UHPLC-MS/MS对其中7个菌株的脂质组成进行非靶向定性分析,研究酵母菌的脂质组成,旨在为酵母的生产应用以及脂质组学的发展提供理论依据。
1 材料与方法
1.1 材料与仪器
1.1.1 材料与试剂
腐乳毛胚、酸浆水:广东某腐乳厂;
阿米塞毕赤酵母(PichiaamenthioninaY):实验室前期从豆腐酸浆水中分离纯化得到;
马克斯克鲁维酵母(KluyveromycesmarxianusK):科汉森公司;
酵母DNA快速提取试剂盒:广州东盛生物科技有限公司;
氯仿、甲醇:色谱纯,美国Sigma公司。
1.1.2 主要仪器设备
超高效液相串联质谱:UHPLC-/Q-Exactive Plus型,美国Thermo scientific公司;
高速冷冻离心机:CR22G型,日本日立公司;
紫外分光光度计:UV-5500PC型,上海元析仪器有限公司;
医用离心机:JW-HR型,安徽嘉文仪器装备有限公司;
电泳仪:DYY-6C型,北京六一生物科技有限公司;
紫外凝胶成像系统:WD-9413B型,广州誉维生物科技仪器有限公司;
基因扩增仪:TR-96/G/H(b)C型,杭州博日科技有限公司。
1.2 方法
1.2.1 酵母分离纯化和鉴定
(1) 酵母菌株的分离纯化:分别取1 mL豆腐酸浆水和1 g腐乳毛胚于9 mL无菌生理盐水中进行梯度稀释,选择适宜梯度的稀释液1 mL涂布于YPD平板中,再于28 ℃培养48 h。挑取疑似酵母菌落进行液体培养并划线分离,如此反复3~4次,直至获得纯的单菌落,将划线分离得到的菌株保藏于50%的甘油溶液中,并于-20 ℃保存。
(2) 分子生物学鉴定:对分离获得的菌株进行分子生物学鉴定。采用酵母DNA快速提取试剂盒提取酵母菌的基因组DNA,然后设计ITS序列通用引物ITS1:5’-TCCGTAGGTGAACCTGCGG-3’和ITS4:5’-TCCTCCGCTTATTGATATGC-3’进行 PCR 扩增,扩增条件为94 ℃预变性3 min;94 ℃ 30 s,46 ℃ 30 s,72 ℃ 1 min,共 30个循环;72 ℃延伸10 min。将扩增得到的 DNA进行测序,并在 NCBI数据库中进行 BLAST 比对。
1.2.2 脂肪酶活测定
(1) 粗酶液的制备:将过夜活化的酵母菌液按体积分数1%接种于YPD液体培养基中,30 ℃培养24 h,离心获得酵母菌体,用无菌水洗涤2次,获得酵母种子液,调节菌液浓度为108CFU/mL,按体积分数1%接种于20 mL YPD培养基中,30 ℃摇床培养24 h,离心,测定上清液脂肪酶活。菌体用无菌水洗涤后,重悬于无菌水中,调节菌液OD600 nm值为2.5,作为粗酶液用于测定细胞结合型的脂肪酶活。
(2) 酶活测定:参照GB/T 23535—2009并修改。试验组锥形瓶中加入 4 mL 4%聚乙烯醇溶液和 5 mL pH 7.5 的磷酸缓冲液,37 ℃保温5 min,再加入 1 mLOD600 nm为2.5的酵母菌液或培养基上清液,混匀,37 ℃保温30 min,随后加入15 mL乙醇(95%)终止反应。用NaOH溶液(0.026 mol/L)滴定至中性,记录消耗NaOH溶液体积。对照组则先加入乙醇溶液终止反应,再加入酶液。
1.2.3 脂质组成分析
(1) 酵母培养及脂质的提取:将过夜活化的菌液按体积分数1%接种到YPD培养基,28 ℃摇床培养48 h,离心收集菌体,用无菌水洗涤2次,再次离心收集菌体,冻干保存。取0.12 g冻干的酵母菌体于带螺旋盖的玻璃试管中,再加入2 mL氯仿—甲醇(1∶1)溶液,涡旋提取1 min。加入2 mL 0.8 mol/L的KOH—甲醇溶液,42 ℃、160 r/min下孵育30 min。加入5 mL氯仿和2.25 mL 水涡旋直至皂化,3 000 r/min离心10 min,取下层有机相过有机膜进样检测[10]。
(2) UHPLC-ESI-MS/MS:色谱柱为ACQUITY UPLC®BEH C18(2.1 mm×100 mm,1.7 μm),质谱系统为配备ESI离子源和Orbitrap的API14000质谱仪。色谱分离采用二元的梯度洗脱,溶剂A为添加0.1%甲酸的水溶液;溶剂B为甲醇;流速300 μL/min,进样量2 μL。洗脱梯度为0 min,85% B;2 min,85% B;20 min,100% B;22 min,100% B;22.1 min,85% B。正离子模式下进行质谱分析,扫描范围150~10 000。所有谱图数据处理采用X caliber 2.0软件分析,并与MZcloud数据库进行比对确定物质的相关信息。
2 结果与分析
2.1 酵母菌的分离
由表1可知,酸浆水体系中共分离得到10个酵母菌株,除了1株膜醭毕赤酵母(Pichiamembranifaciens)ATW-6外,其他的9个菌株均为嗜酒假丝酵母(Candidaethanolica),说明嗜酒假丝酵母的耐酸性较好,酸浆水过酸的环境不适宜其他酵母的生长。嗜酒假丝酵母也存在于红茶菌[11]、新疆传统发酵酸驼乳[12]、龙舌兰酒[13]等食品中。膜醭毕赤酵母主要应用于柑橘青绿霉病的控制,通过与病原菌进行营养和空间竞争,诱导果树产生抗病性[14]。

表1 腐乳酸浆水中酵母分离菌株鉴定结果
由表2可知,毛胚中共分离得到13个菌株,包括3株马克斯克鲁维酵母(Kluyveromycesmarxianus)、3株Diutina(Candida)mesorugosa、2株库德里阿兹威毕赤酵母(Pichiakudriavzevii)、3株挪威毕赤(Pichianorvegensis)和2株阿萨希毛孢子菌(Trichosporonasahii)。马克斯克鲁维酵母已通过美国GRAS和欧洲QPS安全认证[15],可在高达45~52 ℃的温度下生长,其碳源代谢谱十分广泛[16]。该菌种在面包生产中具有一定研究和应用价值,能够改善发酵面包的风味和质构[17-18]。库德里阿兹威氏毕赤酵母在西非一种自然发酵的面团Mawe中是优势菌种[19],该菌种也存在于猪肉肠、芝士、葡萄酒渣、可可豆和米糠中[20]。Hong等[21]发现,库德里阿兹威氏毕赤酵母SJP-SNU含有致病性相关基因,但未见其致病性报道,同时含有植酸酶和纤维素酶基因,该菌株的益生特性和安全性需进一步研究。P.kudriavzeviiMF-121[21]和P.kudriavzeviiDMKU 3-ET15[22]在高温条件下也能高产乙醇,其MF-121菌株对酸、乙醇、温度、盐均具有较高的耐受性[23-24]。Hellström等[25]从一种坦桑尼亚的发酵食品中分离得到挪威毕赤酵母;且发酵初期的挪威毕赤酵母占其中非酿酒酵母的14%[26]。Diutina(Candida)mesorugosa与某些感染性疾病相关[27]。阿萨希毛孢子菌是一种罕见的条件性真菌病原体,可导致免疫缺陷患者发生系统性感染,甚至对嗜中性粒细胞减少的患者具有生命威胁[28-29]

表2 腐乳毛胚中酵母分离菌株鉴定结果
考虑到菌种的安全性和差异性,从分离的23株菌株中各选择1株嗜酒假丝酵母ATW-1、膜醭毕赤酵母ATW-6、马克斯克鲁维酵母SP-1、库德里阿兹威毕赤酵母SP-4和挪威毕赤酵母SP-5共5株酵母菌株进行后续试验,并选取实验室前期分离的阿米塞毕赤酵母(Pichiaamenthionina)Y和商业菌株马克斯克鲁维酵母(Kluyveromycesmarxianus)K进行对比。前期研究[30]表明,阿米塞毕赤酵母Y(Pichiaamenthionina)对温度和NaCl具有较高的耐受性,能降低大豆豆腥味成分,产生酯类香气成分。
2.2 脂肪酶酶活
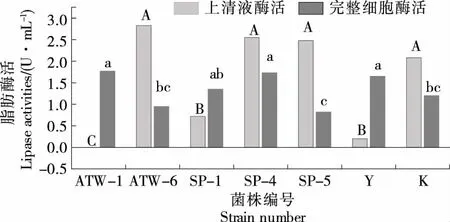
由图1可知,菌株ATW-6、SP-4、SP-5与K的上清液脂肪酶酶活显著高于其他3个菌株,为2.08~2.83 U/mL,菌株ATW-1与菌株Y的上清液基本无脂肪酶酶活。胡珺等[31]发现,未优化产酶条件前,芽孢杆菌酶活仅为2.17 U/mL。枯草芽孢杆菌U5脂肪酶酶活为3.30 U/mL,优化后可达4.30 U/mL[32]。菌株ATW-6的上清液酶活最高,为2.83 U/mL,与上述提到的芽孢杆菌进行对比,其优势比较突出。就细胞结合型脂肪酶活而言,菌株ATW-1、SP-4和Y的酶活高于其他菌株,基本处于1.70 U/mL左右,说明上清液脂肪酶酶活与细胞结合型脂肪酶活之间不存在必然联系。
2.3 脂质非靶向分析
由图2可知,菌株ATW-1、ATW-6、SP-4、Y和K的峰位置和强度较为接近,0.98,6.36,8.09 min处的相对响应较强。而菌株SP1、SP5的中间离子峰信号较弱,说明这两株酵母与其他酵母的脂质组成存在差异。
字母不同表示差异显著(P<0.05)
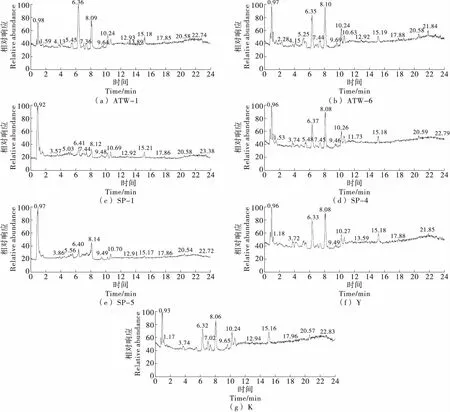
图2 酵母提取物的UHPLC-MS/MS总离子流图
由图3可知,酵母脂质化合物包括2种鞘氨醇、16种酰胺、22种酯类、3种脂肪酸4大类物质。酰胺是以长链脂肪酸酰胺为主;酯类主要包括长链脂肪酸甲酯、芳香酯类和单甘油酯3大类;脂肪酸包括二十碳五烯酸、二十二碳六烯酸和α-亚麻酸3种。其中,酯类是酵母脂质的主要组成成分,同时菌株ATW-1、SP-4与Y检测出的酯类物质相对多于其他菌株。这3个菌株的上清液脂肪酶活较低,但其细胞结合型脂肪酶活却较高,与其代谢产物中存在较为丰富的酯类物质似乎有一定的相关性。
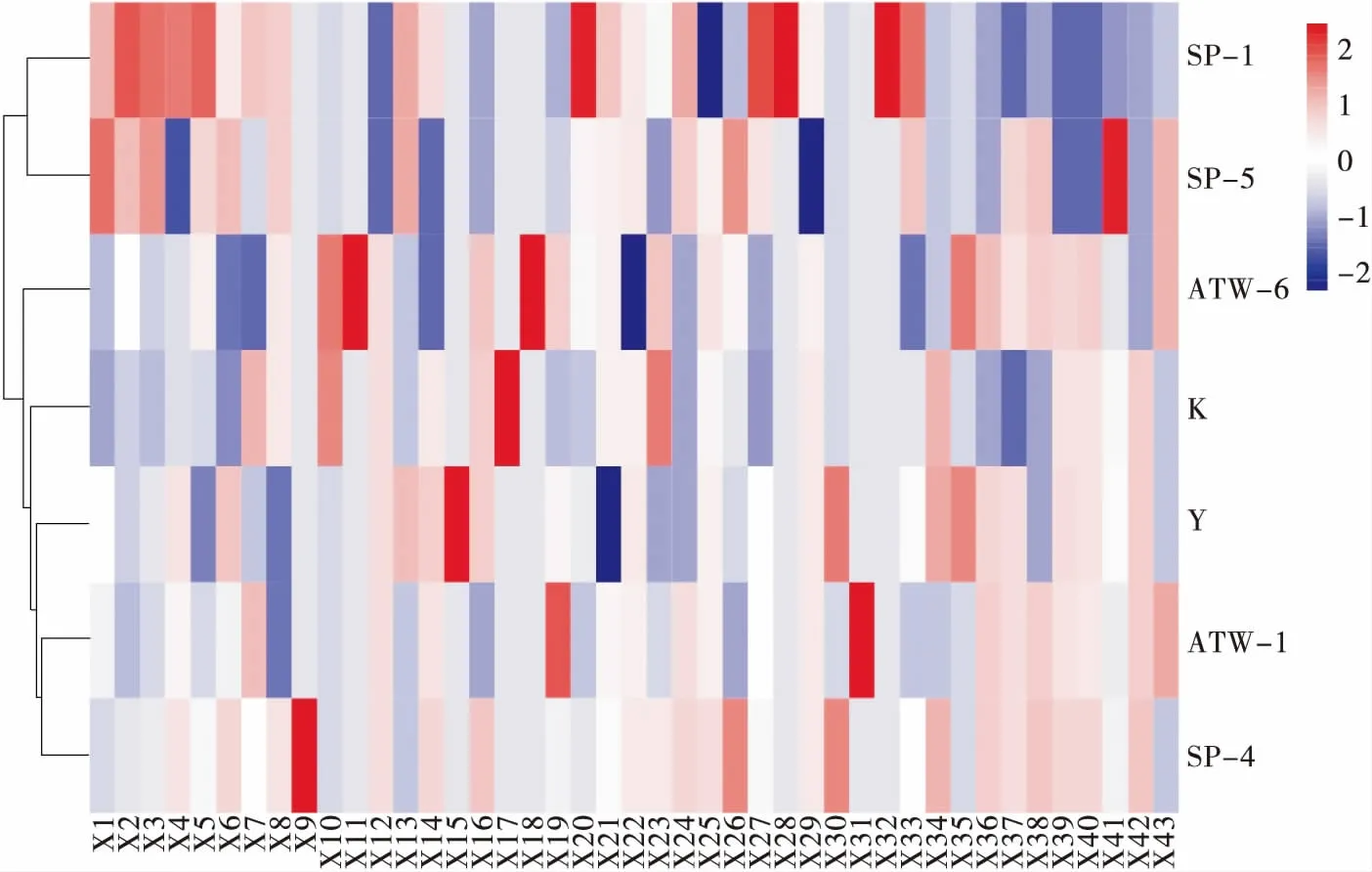
X1. N,N-二甲基鞘氨醇 X2. 鞘氨醇 X3. 十六酰胺 X4. 硬脂酰胺 X5. 油酸酰胺 X6. 烟酰胺 X7. 芥酸酰胺 X8. 二十二酰胺 X9. α-亚麻酸乙醇酰胺 X10. 脯氨酸酰胺 X11. 油酰乙醇酰胺 X12. N,N-二乙基-3-甲基苯甲酰胺 X13. 十八碳四烯酰甘氨酸 X14. N-乙酰腐胺 X15. N1-乙酰精胺 X16. 戊基芬太尼 X17. 油酰苯胺 X18. N8-乙酰亚精胺 X19. 9(Z),11(E),13(E)-十八碳三烯酸甲酯 X20. 三(2,4-二叔丁基)亚磷酸苯酯 X21. 十六烯酸甲酯 X22. 邻苯二甲酸二甲酯 X23. 棕榈酸甲酯 X24. 顺式-12-十八酸甲酯 X25. 蓖麻油酸甲酯 X26. 3-(3,5-二叔丁基-4-羟基苯基)丙酸正十八烷醇酯 X27. 新蛇床内酯 X28. 邻苯二甲酸二异丁酯 X29. 二十二碳六烯酸甲酯 X30. 异烟酸甲酯 X31. 滨蒿内酯 X32. 鲁望素 X33. 4-羟基香豆素 X34. 2-花生酰甘油酯 X35. 1-硬脂酰甘油酯 X36. 2-亚油酸甘油酯 X37. 纽甜 X38. 鳄梨酸1-乙酸酯 X39. 2-(14,15-环氧二十碳三烯酰基) 甘油酯 X40. PPG Acrylate n10 X41. 二十碳五烯酸 X42. 二十二碳六烯酸 X43. α-亚麻酸
由图4可知,前3个主成分的积累方差贡献达到了71.5%。基于前两个主成分可知,菌株SP-4、ATW-1、Y和K的位置较为接近,位于第1组分的负端和第2组分的正端,说明样品组成较为相似,与TIC图一致;而菌株SP1、SP5、ATW-6则分别分布于第1、第4和第3象限,说明与其他菌株脂质组成差异较大。
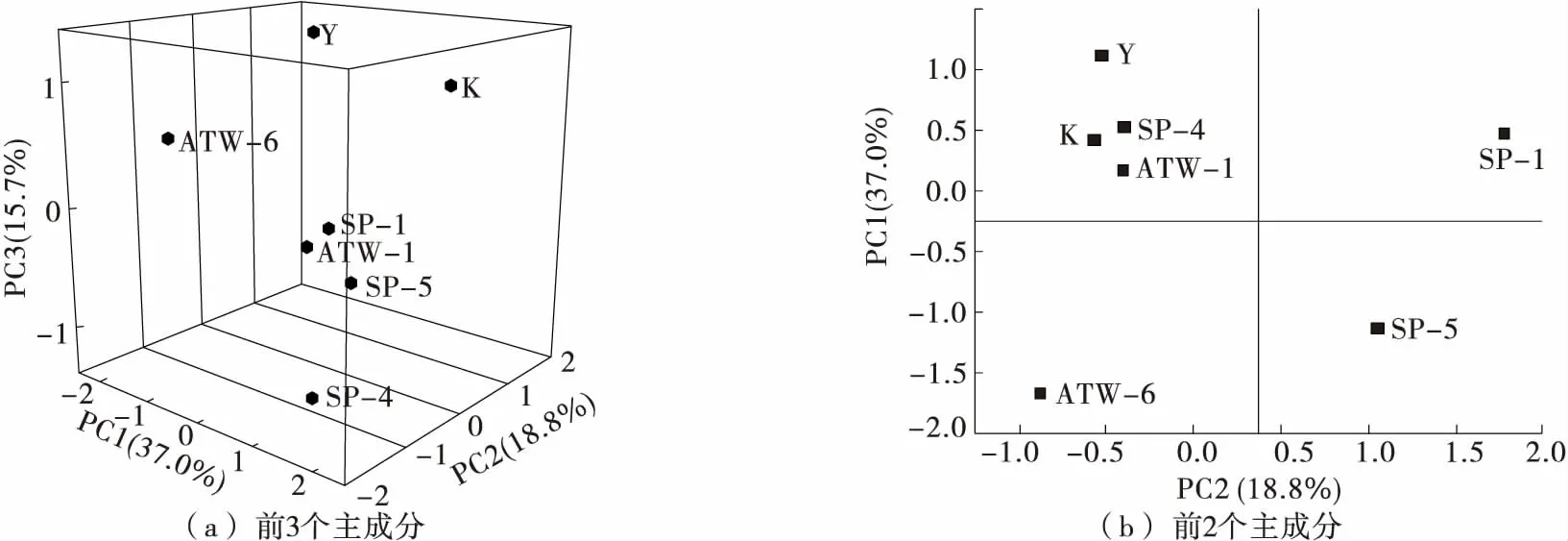
图4 主成分分布图
2.4 鞘氨醇代谢分析
7株酵母菌株中均存在N,N-二甲基鞘氨醇和植物鞘氨醇。鞘氨醇属于鞘脂化合物,是合成神经酰胺的前体物质[33],而鞘脂在哺乳动物角质层的保水性和表皮通透性屏障功能中发挥着重要作用[34]。
由图5可知,马克斯克鲁维酵母SP-1中植物鞘氨醇相对含量最高,其次为挪威毕赤酵母SP-5。植物鞘氨醇是较为常见的一种鞘氨醇,解脂假丝酵母(Candidalipolitica)33M菌株所产神经酰胺主要是以18碳和20碳的植物鞘氨醇与脂肪酸酰化形成[35],酵母神经酰胺的磷酸肌醇衍生物均以含植物鞘氨醇基的鞘磷脂化合物为主[36]。挪威毕赤酵母SP-5中N,N-二甲基鞘氨醇相对含量最高,其次为SP-1。N,N-二甲基鞘氨醇是氨基上的氢被两个甲基取代而形成,二甲基鞘氨醇不但是一种蛋白激酶C的抑制剂,也是特异性的鞘氨醇激酶抑制剂,其抗癌作用已成为化疗发展的基础。由于鞘氨醇及其衍生物种类和结构的多样性,不同培养条件、酵母菌中鞘脂的种类也存在差异[37-38],酵母菌株中极有可能存在其他未知结构的鞘脂类化合物。
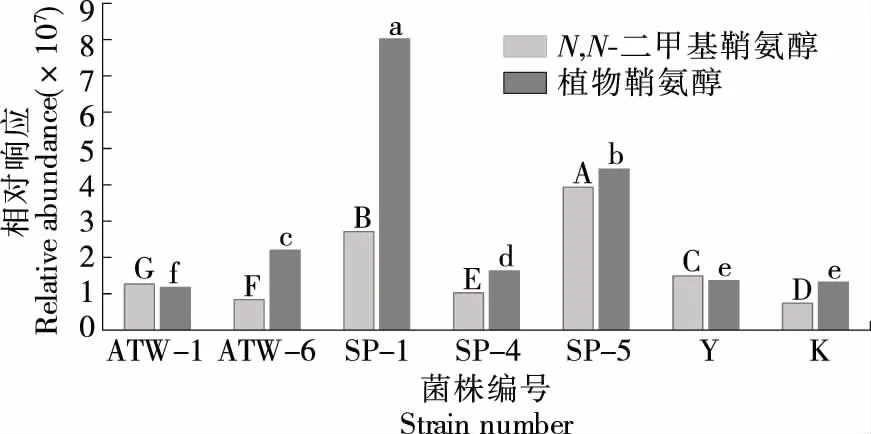
图5 酵母中植物鞘氨醇和N,N-二甲基鞘氨醇的相对响应
3 结论
从豆腐酸浆水和腐乳毛胚中共分离得到23个菌株,利用ITS测序技术进行菌种鉴定。结果表明,酸浆水中分离得到1株膜醭毕赤酵母(Pichiamembranifaciens)和9株嗜酒假丝酵母(Candidaethanolica);毛胚中分离得到3株马克斯克鲁维酵母(Kluyveromycesmarxianus)、3株Diutina(Candida)mesorugosa、2株库德里阿兹威毕赤酵母(Pichiakudriavzevii)、3株挪威毕赤酵母(Pichianorvegensis)和2株阿萨希毛孢子菌(Trichosporonasahii)。通过对酵母菌脂肪酶活和脂质组成进行分析发现,菌株ATW-1、SP-4和Y的细胞结合型脂肪酶活高于其他菌株,同时菌株ATW-1、SP-4和Y中的酯类物质相对比较丰富,PCA分析进一步表明这3个菌株脂质组成具有一定相似性。酵母菌中丰富的代谢酯类物质可能与细胞结合型脂肪酶之间存在密切联系,与上清液的脂肪酶酶活力无关。马克斯克鲁维酵母SP-1和挪威毕赤酵母SP-5中鞘氨醇相对响应较高,推测这两个菌株在鞘脂合成方面具有一定研究潜力,后续可进行鞘脂的定量分析,充分挖掘酵母中丰富的脂质资源。
猜你喜欢
分子催化(2022年1期)2022-11-02
中华胰腺病杂志(2022年4期)2022-08-23
医学概论(2022年4期)2022-04-24
食品安全导刊(2021年20期)2021-08-30
磷肥与复肥(2021年5期)2021-06-19
当代水产(2020年3期)2020-06-15
求知导刊(2019年15期)2019-08-30
广西科技大学学报(2018年2期)2018-09-10
家庭医学(2017年3期)2017-05-27
现代农业科技(2016年20期)2016-12-20
