
DNA甲基转移酶3A调控少突胶质细胞发育
2020-08-07 12:57方敏溪杨爱芬徐晓锋
杭州师范大学学报(自然科学版) 2020年4期
方敏溪,俞 倩,易 敏,杨爱芬,徐晓锋
(杭州师范大学生命科学研究院,浙江省器官发育与再生技术研究重点实验室,浙江 杭州 311121)
0 引言
少突胶质细胞(Oligodendrocytes, OLs)由少突胶质前体细胞(Oligodendrocyte precursor cells, OPCs)经过增殖、迁移、分化形成,是脊椎动物中枢神经系统(central nervous system, CNS)唯一的髓鞘形成细胞[1].髓鞘包裹在神经元周围能够确保神经冲动在神经元快速准确地传导[2].近年来的研究表明,由于OLs谱系分化成熟障碍、再生受阻或者OLs功能低下,导致髓鞘结构或功能异常与许多神经系统疾病的发生相关,如多发性硬化症(multiple sclerosis, MS)、横贯性脊髓炎(transverse myelitis)以及阿尔茨海默症(Alzheimer’s disease)等[3-5].刺激CNS内保留的OPCs向OLs的再分化和重新髓鞘化将成为治疗脱髓鞘相关疾病的有效途径之一.
随着表观遗传学概念的不断深化和研究方法的发展,包括DNA甲基化和组蛋白修饰在内的表观遗传调控在少突胶质细胞发育和脱髓鞘疾病治疗中的作用逐渐引起人们的关注[6].高等动物的DNA甲基化一般发生在鸟嘌呤二核苷酸(CpG)上,CpG序列中胞嘧啶的甲基化会关闭基因启动子的活性,从而抑制其表达[7].研究显示,许多脱髓鞘神经退行性疾病存在DNA甲基化的改变,MS患者脑组织中DNA去甲基化的活性比正常人脑组织高出了两倍[8],因此,DNA甲基化可能作为MS疾病活性标记物.其中,DNA甲基转移酶3A (DNA methyltransferases 3A,DNMT3A) 负责催化DNA的从头甲基化,参与调控多种器官的发育.分离Dnmt3a基因敲除小鼠的神经干细胞进行体外诱导分化,发现分化为神经元的细胞数量减少,分化为胶质细胞的数量增多[9],表明DNMT3A介导的DNA甲基化参与调控神经元和胶质细胞的命运特化.目前为止,DNMT3A对体内OPCs分化和OLs发育的调控作用还未见报道.本研究以Dnmt3a全局性敲除小鼠为对象,分析敲除Dnmt3a对少突胶质细胞发育的影响,结果发现DNMT3A功能缺失后造成OLs分化的延迟,处于未分化状态的OPCs数量增加.因此,DNMT3A对于小鼠少突胶质细胞的正常发育是必需的.
1 材料和方法
1.1 动物

表1 Dnmt3a小鼠基因型鉴定引物[10]Tab.1 The genotyping primer of Dnmt3a mice
Dnmt3a+/-品系(Stock No: 018838)小鼠从美国Jackson Lab平台购置,特异性缺失Dnmt3a基因组第13至第17个外显子序列,已被广泛用于研究Dnmt3a基因敲除后对不同组织或器官发育的影响.本实验用到的全局性Dnmt3a敲除小鼠(Dnmt3a-/-),是由Dnmt3a+/-杂合小鼠相互交配获得.得到的后代采取碱裂解法鉴定基因型,鉴定引物如表1所示(由Jackson Lab平台提供).
1.2 脊髓组织取材
收集出生前后不同时期(E14.5,E16.5,E18.5,P0,P3,P7和P10)小鼠脊髓组织.胚胎期小鼠直接剥离新鲜组织,用预冷的PBS润洗后置于4%多聚甲醛(PFA)中固定过夜.出生后小鼠先依次用PBS和4%PFA进行灌注,然后分离出脊髓组织,置于4%PFA中固定过夜.次日将组织转入30%蔗糖溶液中进行脱水处理,4 ℃放置过夜.随后,将组织放入OCT包埋剂中,-80 ℃保存.
1.3 RNA原位杂交
RNA原位杂交用小鼠脊髓组织切片厚度为18 μm.RNA原位杂交第1天,提前将组织切片从-80 ℃取出,恢复至室温.50 ℃烘烤30 min,4%PFA室温固定10 min,然后依次经过蛋白酶K消化,4%PFA再固定以及乙酸酐处理,用不含探针的杂交液室温孵育至少1 h.最后,用含RNA探针(终浓度为0.1~0.2 ηg/μL)的杂交液于65 ℃孵育过夜.RNA原位杂交第2天,将多余杂交液洗脱,用含10%绵羊血清的封闭液封闭至少1 h,用含偶联碱性磷酸酶(AP)的抗地高辛抗体于4 ℃孵育过夜.RNA原位杂交第3天,将多余的抗体洗脱,使用NBT/BCIP显色液室温避光显色,至由明显蓝紫色信号出现终止显色反应.甲醇洗脱背景色,封片,在光学显微镜下观察拍照.
1.4 统计学方法
本文中的数据均使用Graphpad Prism 6.0统计分析作图,重复样本数n≥3,通过t检验对统计结果进行差异显著性分析,当P<0.05时差异具有统计学意义.
2 结果与分析
2.1 Dnmt3a在不同发育时期小鼠脊髓组织中的表达
为了确定Dnmt3a的空间表达模式,我们通过RNA原位杂交检测了不同发育时期小鼠脊髓组织中Dnmt3a的表达.结果发现,早期Dnmt3a在灰质区域新分化或成熟的神经元中广泛表达.从E14.5开始,白质区域开始出现零星分布的Dnmt3a+细胞(图1 A).随着发育的进行,白质中Dnmt3a+细胞逐渐增多(图1 B),一直持续到P7(图1 C),随后Dnmt3a在该区域的表达逐渐消失.Dnmt3a在白质中的表达时期正是脊髓组织中少突胶质细胞开始大量分化的时期,暗示其可能参与调控该过程.

A-C:Dnmt3a在E14.5, E18.5和P7野生型小鼠脊髓中的RNA原位杂交结果. E14.5期开始在白质区域表达,持续至P7. 箭头表示Dnmt3a在白质中的表达.
2.2 Dnmt3a基因敲除对少突胶质细胞分化的影响
与已有报道一致,Dnmt3a纯合突变小鼠(Dnmt3a-/-)呈现发育不良、个体矮小、小脑畸形等表型,出生后三周左右死亡[10].为了判断Dnmt3a敲除是否会造成OLs发育异常,我们收集出生前后不同时期Dnmt3a-/-和野生型小鼠脊髓组织,通过RNA原位杂交检测髓鞘相关基因Mbp和Plp1的表达.如图2和图3,野生型小鼠脊髓组织分别于E14.5天和E16.5天开始检测到Mbp(图2 A)和Plp1(图3 A)在腹侧中线区域的表达.随着发育的进行,Mbp+和Plp1+的少突胶质细胞逐渐增多,主要分布在脊髓白质区域.而在Dnmt3a-/-小鼠中,Mbp和Plp1的信号产生出现明显滞后.E16.5期开始在腹侧中线附近明确检测到Mbp的表达(图2 B’),P0期左右在腹侧白质区域检测到Plp1的表达(图3 B’).随后突变体中少突胶质细胞的分化没有被抑制,细胞数目逐渐增多,至P7天与野生型基本一致.进一步对Plp1+细胞进行定量分析,发现Dnmt3a-/-小鼠中,Plp1+细胞数目的差异在OLs分化初期最明显,随着发育的进行,这种差异逐渐缩小,至P10左右无明显差异(图3 E).通过对髓鞘相关基因的表达进行分析,我们发现Dnmt3a基因敲除会延迟少突胶质细胞的分化.
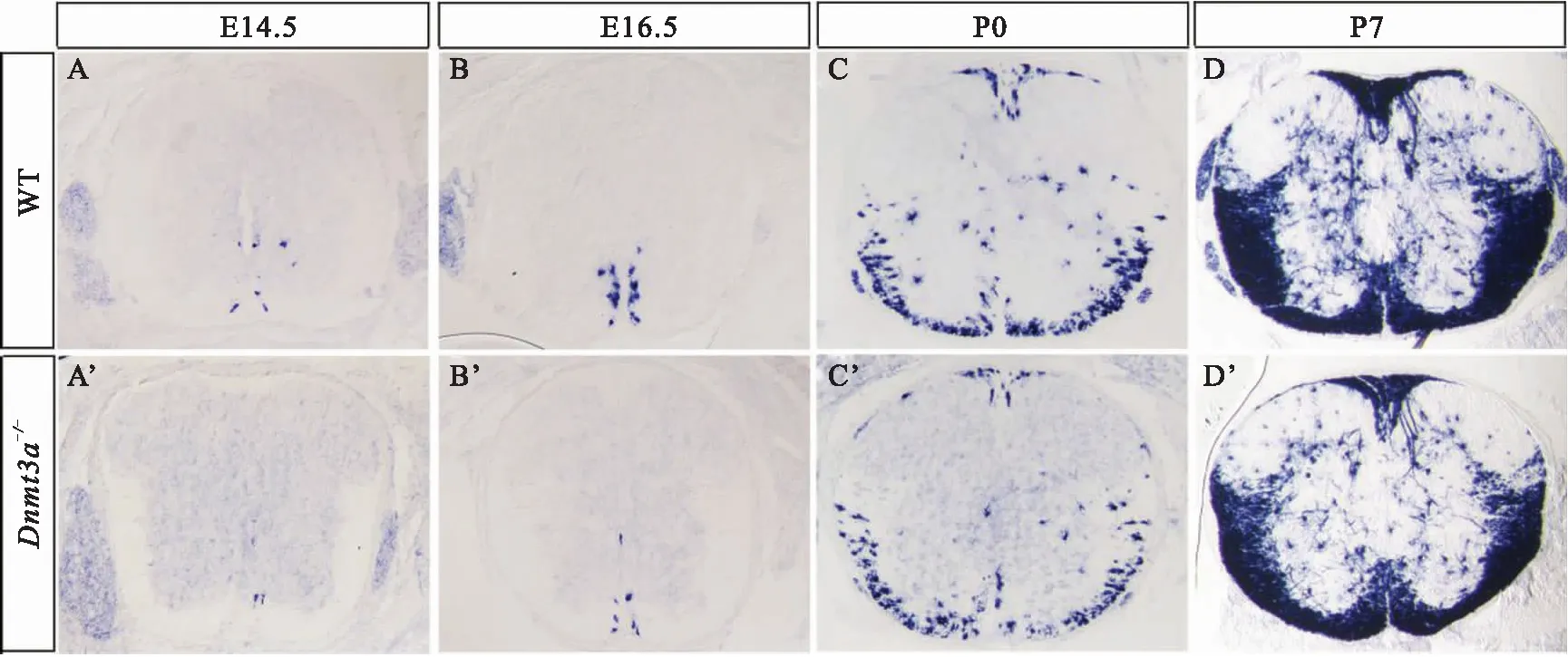
A-D’:Mbp在野生型和Dnmt3a-/-小鼠中的表达分析.与野生型小鼠相比,突变小鼠中Mbp的表达在E14.5时被明显抑制,E16.5开始有少量Mbp+细胞在腹侧出现,P0至P7差异逐渐减小.
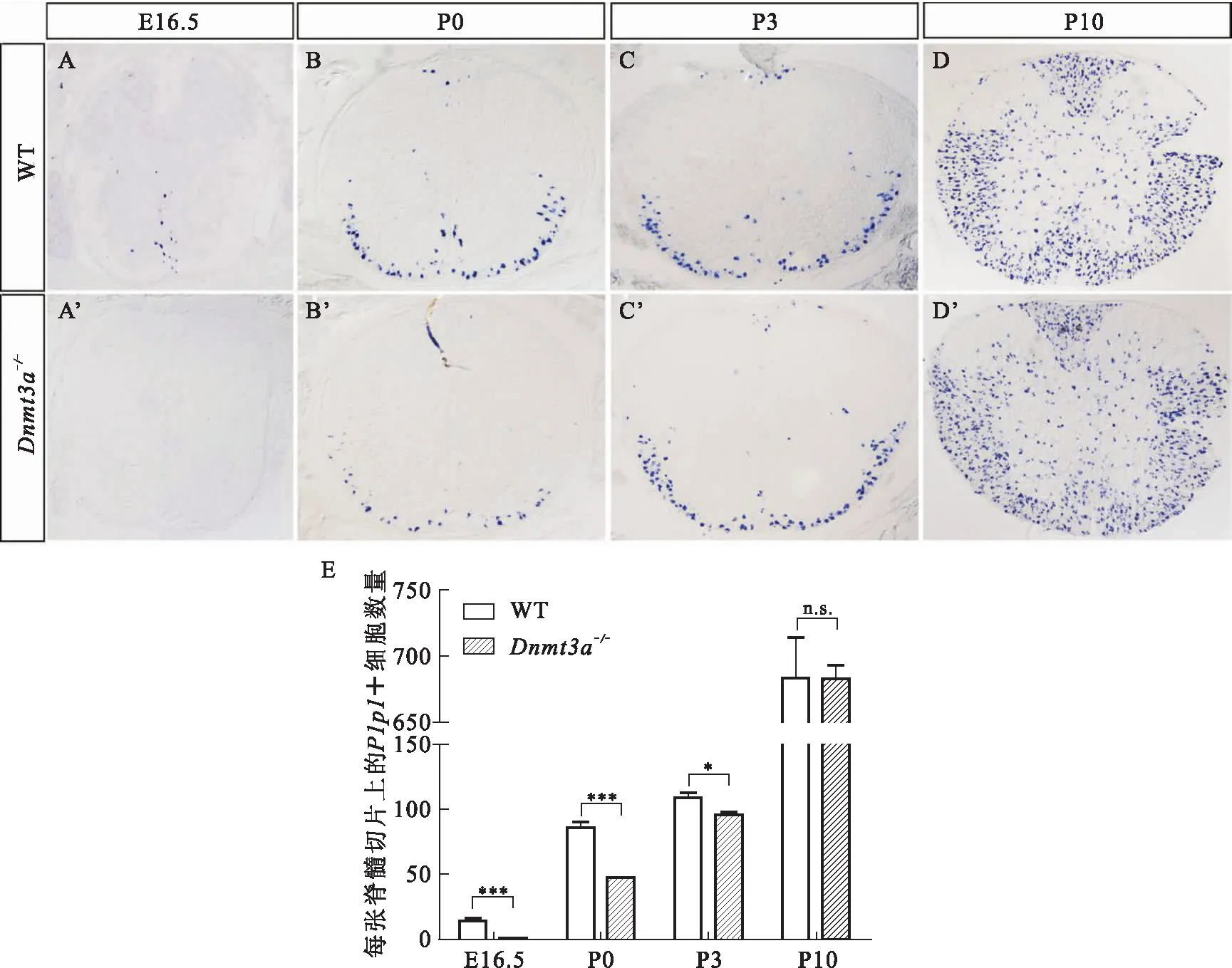
A-D’:Plp1在野生型和Dnmt3a-/-小鼠中的表达分析.与野生型小鼠相比,突变小鼠中Plp1的表达在E16.5时被明显抑制,P0开始有少量Plp1+细胞在腹侧出现,P3至P10差异逐渐减小. E:Plp1+细胞的数量统计结果(n≥3,*P<0.05, ***P<0.005,n.s.:无显著性差异).
2.3 Dnmt3a基因敲除对OPCs产生和增殖影响
考虑到Dnmt3a突变小鼠体内OLs分化的延迟可能与OPCs的异常相关,我们进一步检测了不同时期OPCs 标记基因Pdgfrα的表达情况.如图4所示,在出生前E16.5时期,小鼠体内已经特化的OPCs发生迅速增殖(图4 A).与野生型对照组相比,该时期Dnmt3a-/-小鼠体内Pdgfrɑ+细胞的分布和数目无明显差异(图4 A’),说明Dnmt3a基因敲除不影响早期OPCs的特化.P0时期,OPCs开始大量向OLs分化,Dnmt3a-/-小鼠体内Pdgfrɑ+细胞数目显著增加(图4 B、B’),至P7时期仍明显多于野生型对照组(图4 C、C’、D).因此,Dnmt3a基因敲除会造成分化阶段OPCs数量增加,暗示可能与该阶段OLs分化被延迟相关.
3 讨论
包括DNA甲基化在内的表观遗传调控在少突胶质细胞发育过程中的功能正在被逐渐揭示.其中,DNA甲基转移酶3A(DNMT3A)在体内少突胶质细胞分化和发育过程中的功能尚不清楚.本研究发现,Dnmt3a早期在灰质区域神经元中广泛表达,从E14.5天开始在白质内表达,随后表达量逐渐上升,当OLs形成后表达消失,暗示DNMT3A可能参与调控OLs的起始分化.对Dnmt3a全局性敲除小鼠进行分析,发现Dnmt3a基因敲除后,造成OLs分化的延迟,处于未分化状态的OPCs数量增加.因此,我们的研究表明,DNMT3A的功能对于小鼠体内少突胶质细胞的正常发育是必需的.
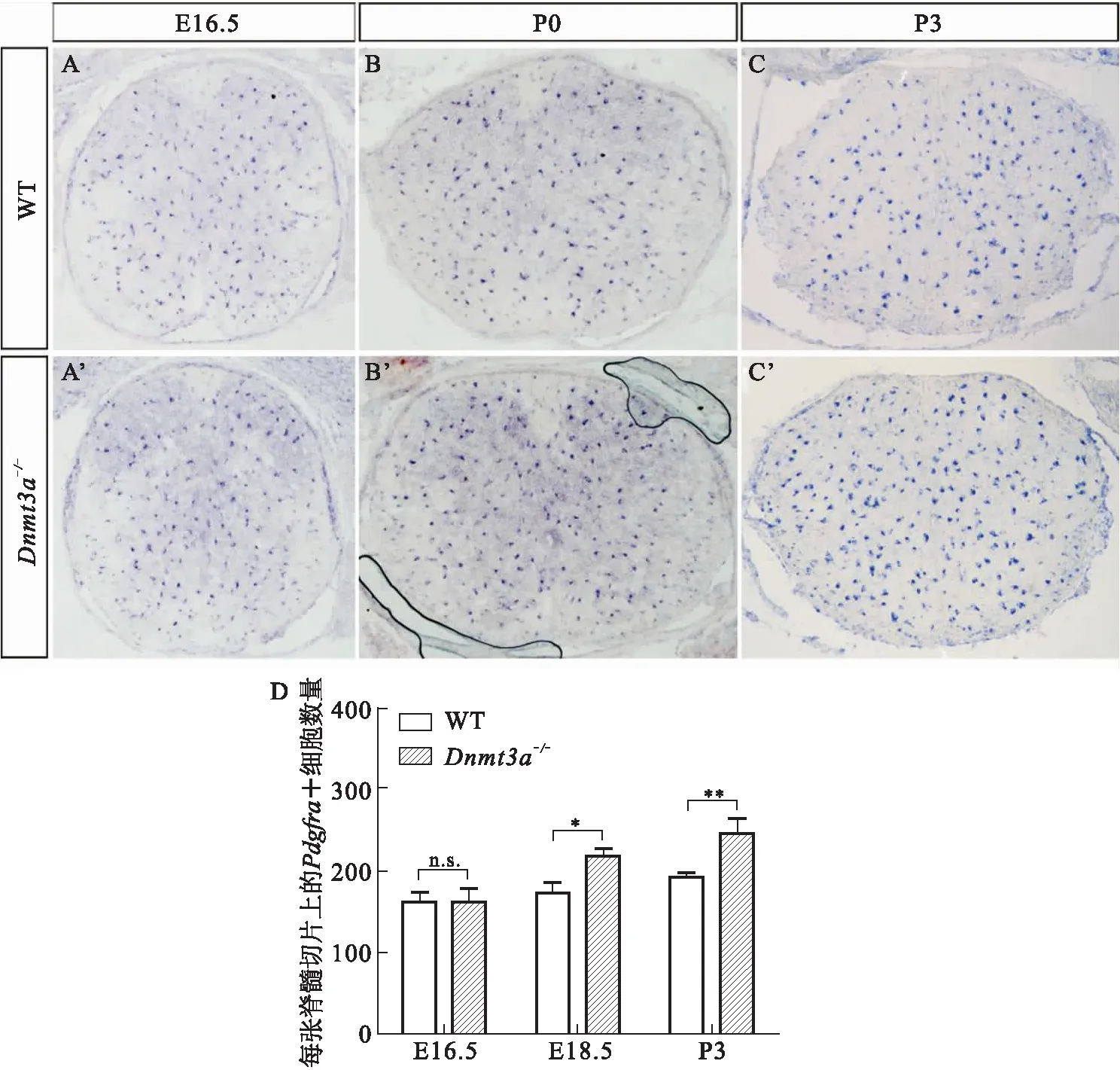
A-C’: Pdgfra在野生型和Dnmt3a-/-小鼠中的表达分析.与野生型小鼠相比,突变小鼠中Pdgfra的表达在E16.5期无明显差异,P0-P3期Pdgfra+细胞数目增多.D: Pdgfra+少突胶质前体细胞的数量统计结果(n≥3, *P<0.05, **P<0.01, n.s.:无显著性差异).
然而,DNMT3A在体内介导上述过程的分子机制尚不清楚.已有研究表明,通过对正常和Dnmt3a敲除的神经前体细胞进行表达谱分析,筛选到与少突胶质细胞增殖和分化相关的3个关键基因(Pdgfrα,Nkx2.2和Mbp)表达水平显著上调,其近端启动子区域高度去甲基化,暗示DNMT3A可能通过特异性诱导上述因子的甲基化,从而阶段性调控OLs的分化[9].与Dnmt3a的表达类似,Nkx2.2在OLs谱系发育过程中具有阶段性表达的特定,且两者动态表达模式相吻合[11-12].我们的早期研究也已经证实,敲除Nkx2.2也会造成OLs分化的延迟[13].综上所述,我们初步推测,DNMT3A介导的DNA甲基化与NKX2.2之间可能存在紧密联系,相互作用协调OPCs的增殖和分化.
猜你喜欢
四川生理科学杂志(2022年4期)2022-12-06
北京大学学报(自然科学版)(2022年2期)2022-04-08
昆明医科大学学报(2021年3期)2021-07-22
昆明医科大学学报(2021年3期)2021-07-22
神经损伤与功能重建(2020年10期)2020-12-23
神经损伤与功能重建(2020年11期)2020-12-01
医学新知(2019年4期)2020-01-02
保健与生活(2019年15期)2019-09-12
中国当代医药(2017年4期)2017-04-06
飞碟探索(2015年11期)2015-09-10
