
马尾松林向香樟林改造林下植物功能多样性研究
2020-08-19 08:26吴科君何欣芮綦远才齐代华
生态学报 2020年13期
黄 超,魏 虹,吴科君,何欣芮,汪 鹏,綦远才,齐代华
三峡库区生态环境教育部重点实验室,重庆市三峡库区植物生态与资源重点实验室,西南大学生命科学学院, 重庆 400715
植物功能多样性是近年来生态学研究的重要内容。所谓功能多样性,是指特定生态系统中物种功能特征的数值和范围,强调群落中物种功能性状的差异[1- 3]。生态系统功能不仅依赖于物种的数目,更依赖于物种所具有的功能性状[4- 5]。相比物种多样性,功能多样性对于生态系统功能的贡献更大,两个具有相同物种数目的群落,因其物种性状的不同,很可能会在功能多样性上表现出较大差异[5- 6]。因此,在面对和解决与群落相关的生态学问题时,越来越多学者提倡用功能性状的多样性来代替物种多样进行研究[7- 9]。
马尾松(Pinusmassoniana)是我国特有的乡土树种,用途广泛,经济价值高,马尾松林也是我国分布最为广泛的针叶林。目前,我国近70%马尾松林为次生纯林,普遍存在多代更新、林分密度大和水平分布不匀等问题,造成局部立木间竞争、分化强烈,加之立地条件较差、容易招致病虫害等缺点,使得林分生产力水平和生态效益均不高,迫切需要对马尾松林进行改造[10]。在我国亚热带地区,马尾松林属于常绿阔叶林的一个演替过渡类型。有学者认为在对马尾松林进行改造时应优先选择适生的接近演替顶级的优势乔木树种,以加快该森林生态系统的演替进程[11- 12]。香樟(Cinnamomumcamphora)为樟科的常绿大乔木,主要分布于我国南方及西南各省区,是我国着力培育、并广泛用于医药、油料工业、用材和绿化造林等领域的一种珍贵树种。同时,香樟也属于当地一种演替顶级的乔木树种。因而,在亚热带地区,香樟常被考虑为马尾松林改造的目标树种之一。
目前,在森林改造研究中,已有许多有关马尾松和香樟林群落的报导。然而由于条件的限制,其中大多数研究要么只涉及到了其中一种群落,要么研究不够深入,缺乏比较深入的对马尾松林改造成香樟林的完整研究。林下植物是森林生态系统中丰富度最大的群体,作为森林生态系统中不可缺少的部分,在森林群落多样性、生态功能稳定性和持续立地生产力等方面都具有特殊的功能和作用[13- 14]。因而,在森林改造过程中,有必要对林下植物进行深入的研究和探讨。
光是光合作用的主导因子,对于林下植物而言更是生长和发育的主要限制因素[13]。而植物的繁殖性状则比其他性状更能深刻反映植物群落空间分布格局及生态过程[15- 16]。因此,本文以马尾松林向香樟林改造过程中4种不同阶段群落为研究对象,选取与林下植物对光照响应和繁殖相关的功能性状特征,进行功能多样性研究,以期探讨马尾松林改造对林下植被部分功能性状和功能多样性的影响;并尝试从植物功能多样性角度,分析不同改造阶段中林下植物群落的动态构建过程,以期为马尾松林改造、林下植物多样性保护、森林经营和管理等各方面提供理论参考。
1 研究区域与方法
1.1 研究区域概况
研究区域位于重庆市北碚区观音峡林场孔雀谷景区,地理位置29°45′28″N,106°26′33″E。该区雨量充沛,年均降雨量1050 mm,年均气温18℃,温暖湿润,海拔500 m左右,属中亚热带东部偏湿性季风气候。微酸性黄壤。
区域内森林植被类型以马尾松天然次生纯林为主,生态效益低下,自20世纪80年代起陆续对其进行了改造。对乔木层马尾松进行50%强度的间伐后,替代性地植入2年龄香樟幼苗,种植密度为2500株/ hm2,并对其周边林下干扰树种进行清理;间隔8年后对乔木层的马尾松再进行一次50%强度的间伐,同时补植了相同密度的香樟幼苗,之后停止干预,由其自然生长。从而,在研究区域内分布有成片的马尾松天然次生林以及不同造林阶段的香樟经济林,为研究马尾松天然次生林改造过程中的林下植物功能多样性提供了理想场所。
1.2 研究方法
1.2.1样地设置
本研究采用空间代替时间的方法,在研究区域内,选取马尾松天然次生林(CK)和改造过程中不同阶段:改造前期(S1)、改造中期(S2)和改造后期(S3)的群落为研究对象,以尽量遵循立地条件相近为原则,于每种群落类型中各设置了1个样地,各样地林分分布面积为2.2—3.0 hm2。其中样地S1是在间伐后的马尾松林中补植了香樟幼苗并生长至5年龄后的改造初期马尾松林-香樟样地,样地S2和S3为第一批植入香樟幼苗生长至15年和35年左右的改造中期和后期的香樟林群落样地。表1为4个样地基本情况。
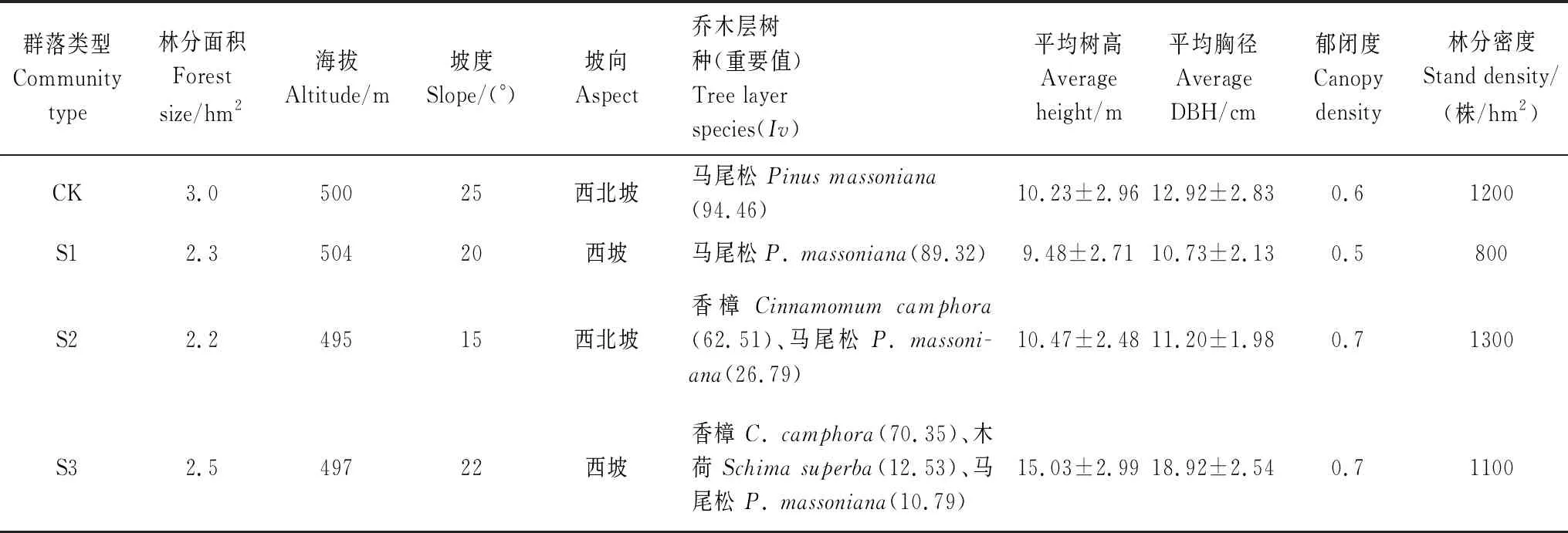
表1 不同改造阶段样地基本情况
1.2.2数据获取
(1) 样方数据获取
2019年6月,于每种样地中随机设置6个25 m2(5 m×5 m)的灌木样方,共24个灌木样方,于每个灌木样方的对角设置2个1 m2(1 m×1 m)的草丛样方,共48个草本样方,样地调查总面积为600 m2,并进行群落学野外调查。群落样方调查记录各样地的海拔高度、地理位置、坡形、坡度、坡向、群落的名称和群落的外貌特征。群落的灌木层由高度小于5 m的木本植物构成,草本层则由草本植物组成。灌木层记录植株的序号、种名、高度、基径/胸径、冠幅;草本层记录植物的种名、数量、高度、盖度。
(2) 功能特征类型的选取
功能性状的选取是功能多样性研究的基础。在对功能性状进行选择时应结合具体的研究目的与背景,重点考虑与生态系统过程和功能相关的生物特征,以提升功能多样性研究的准确性[17- 18]。由于目前对植物功能性状与生态系统过程与功能关系的认知有限,因而,如何选择合适的功能性状成了功能多样性研究中非常棘手却又很重要的问题[18- 19]。有学者根据与生态系统的关系,将功能性状分为响应性状和影响性状。响应性状反映植物对环境因子的响应,影响性状决定植物对生态系统功能的影响,这两类性状在一定程度上相互重叠[20- 21]。本文在对功能性状进行选取时主要参考了该分类方法,同时考虑到环境因子中光照对林下植物的影响[5,7,22]以及繁殖在维持生态系统功能中的重要作用[21- 23],最终选取了13种功能性状类型。根据各功能性状的特征及其相关信息[19,22-26],将“光照习性”、“生活型”、“叶型”、“叶质”和“叶物候”等特征归类于“光响应性状”,“生活史”、“种子扩散方式”、“传粉方式”、“开花期”与“花期时长”划分为“繁殖性状”,而“株高”、“多度”和“盖度”作为通用性状(表2)。

表2 林下植物功能特征类型
植物的“株高”、“多度”和“盖度”这3个功能特征数据通过野外数据调查获得,其余10个功能特征主要通过查阅《中国植物志》获得[27]。
1.2.3数据处理与分析
功能多样性由功能丰富度、功能均匀度和功能离散度3个部分组成[28]。对林下植物各功能性状数据赋值后(表2),分别计算功能树状图指数(Functional diversity based on dendrograms,FD)、二维功能均匀度指数(Function evenness,FEve)和二次熵指数(Quadratic entropy,Rao),分别作为功能丰富度、功能均匀度和功能离散度的代表[29]。采用R3.5.1软件中的FD软件包计算功能多样性指数,使用SPSS.22.0软件单因素方差分析和Tukey多重比较对4个群落功能多样性差异进行统计分析,用Origin软件作图。
FD通过获得各功能性状矩阵,将性状矩阵转化为距离矩阵后,聚类生成树状分类图,计算树状分类图的分枝总长度等步骤完成[4,30]。
FEve和Rao计算公式如下:
式中,EWb为加权平均的均匀度,dij为物种i与物种j之间的欧氏距离,PEWb为局部加权平均均匀度,wi为物种i的相对多度,wj为物种j的相对多度,S为物种数。
2 结果与分析
2.1 林下植物物种和功能性状组成动态
2.1.1物种组成变化
根据野外调查数据,在马尾松林不同的改造阶段中,所有样方中林下植物种类共计47科73属87种。其中,木本植物27科36属45种,优势种主要有:菝葜(SmilaxchinaL.)、香樟(C.camphora)、矩叶鼠刺(Iteaoblonga)、白栎(Quercusfabri)、山莓(Rubuscorchorifolius)、银毛叶山黄麻(Tremanitida)、木荷(Schimasuperba)等;草本植物25科37属42种,优势种有:芒萁(Dicranopterisdichotoma)、淡竹叶(Lophatherumgracile)、蕨(Pteridiumaquilinum)、狗脊(Woodwardiajaponica)、芒(Miscanthussinensis)、石香薷(Moslachinensis)。
不同阶段林下物种数量分布情况见图1。马尾松林改造后,3个阶段林下灌木层和草本层的物种数目均高于对照组。其中,改造中期林下物种数目最高,其次为改造前期。林下物种数目的增加主要体现在草本层,灌木层的物种数目相对稳定。
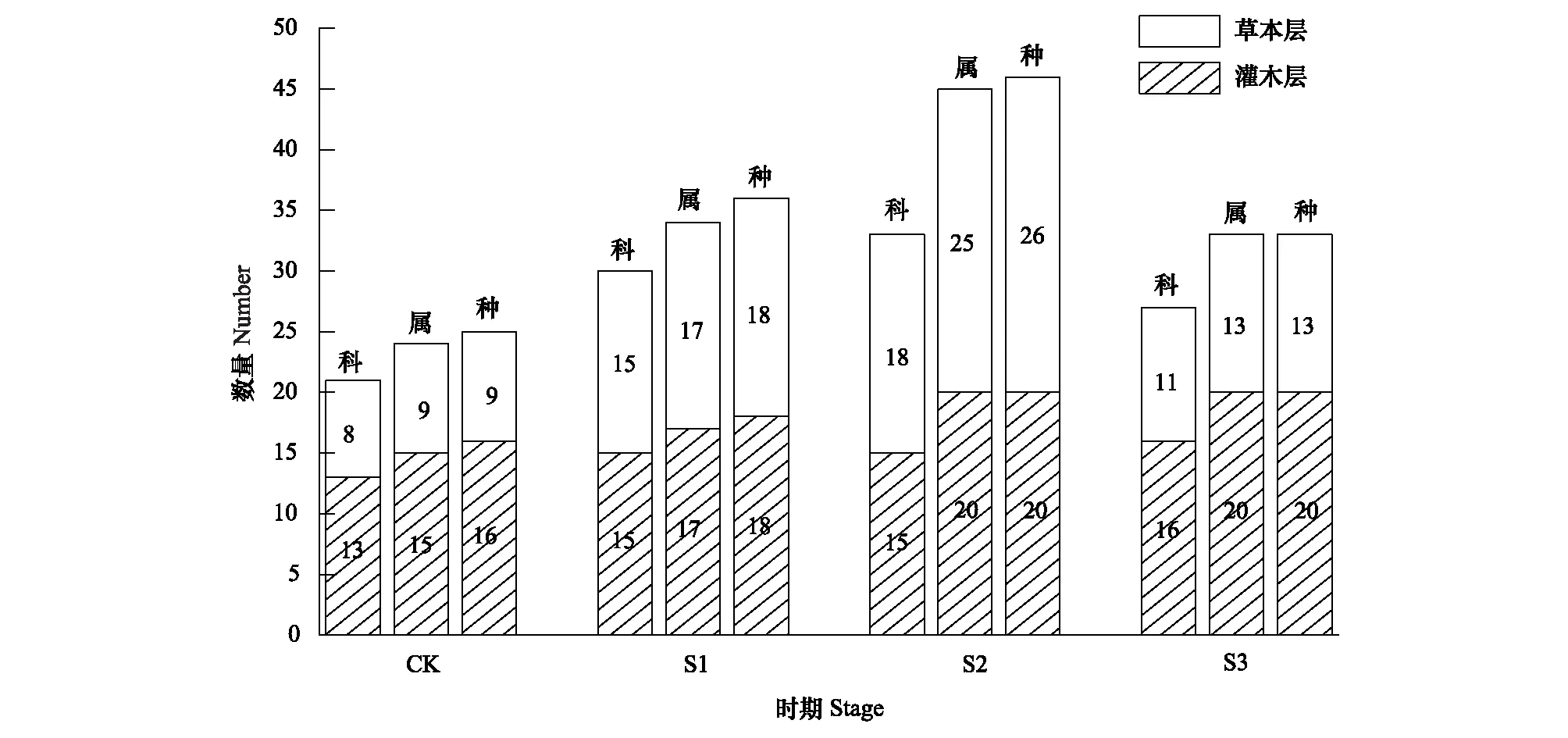
图1 马尾松林改造不同阶段林下物种组成动态Fig.1 Dynamics of undergrowth species at different stages of P. massoniana forest transformationCK: 马尾松林(对照),S1:改造前期群落,S2:改造中期群落,S3:改造后期群落
2.1.2功能性状组成变化
马尾松林改造成香樟林过程中,林下植物不同功能性状的组成变化存在差异,体现了植物功能性状特征的多样性(表3)。
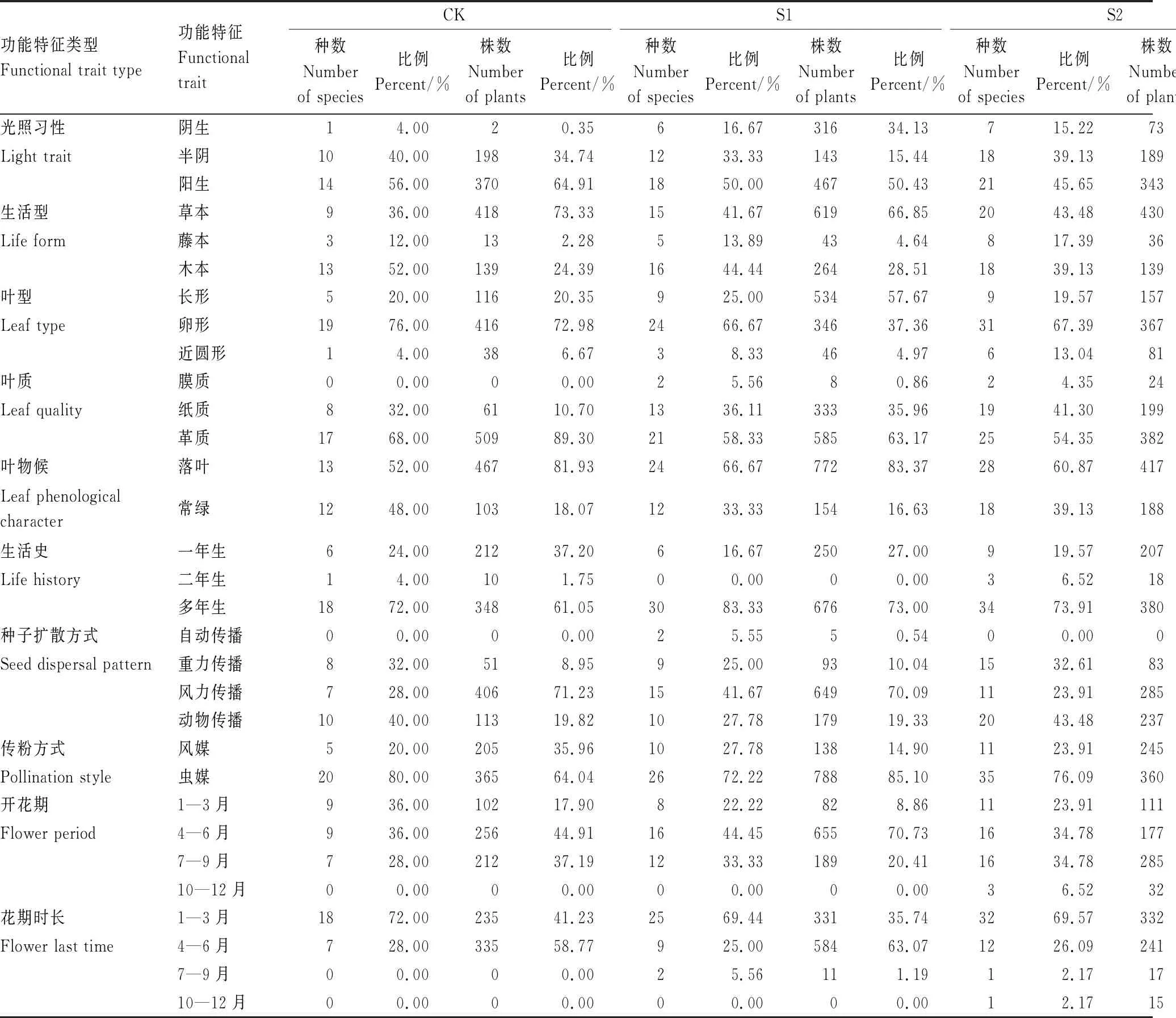
表3 改造过程中林下植物功能性状的动态组成
从林下植物组成看,“光响应性状”的改变较为显著。阳生植物在改造过程中占据的物种比例一直较高,但随着改造的推进,阳生植物的多度比例表现出下降的趋势,阴生植物比例逐渐增加。叶候特征中,落叶和常绿的物种比例组成变化较小,而常绿物种的多度比例在改造后期大幅增加。林下植物叶片形状的变化主要体现在改造前期,叶片长条形特征的植物多度比例增加并超过了叶片卵形植物。随着森林改造进程的推进,林下植物生活型与叶质的性状组成变化相对较小,未形成明显比例变化的趋势。
“繁殖性状”功能特征组成中,种子扩散方式为重力传播的物种比例在改造中期增加。通过风媒传粉的植物多度比例在改造后期超过了虫媒植物。开花期为4—6月的植物的多度比例在改造前期增加,花期时长在1—3月内的植物的多度比例则从改造中期起便超过了花期时长在4—6月的植物。
2.2 林下植物功能多样性的动态变化
2.2.1综合功能多样性
在马尾松林向香樟林的改造过程中,林下植物综合功能多样性指数的动态变化见图2。功能丰富度指数FD和功能均匀度指数FEve的变化趋势相近,在未改造时期(CK),林下植物整体功能丰富度指数最低,在改造前期(S1)突增,在改造中期(S2)和后期(S3)则缓慢下降。林下植物的功能离散度指数Rao在改造后的表现为持续上升,于改造中期(S2)达到了峰值,但在后期(S3)出现了明显下降。
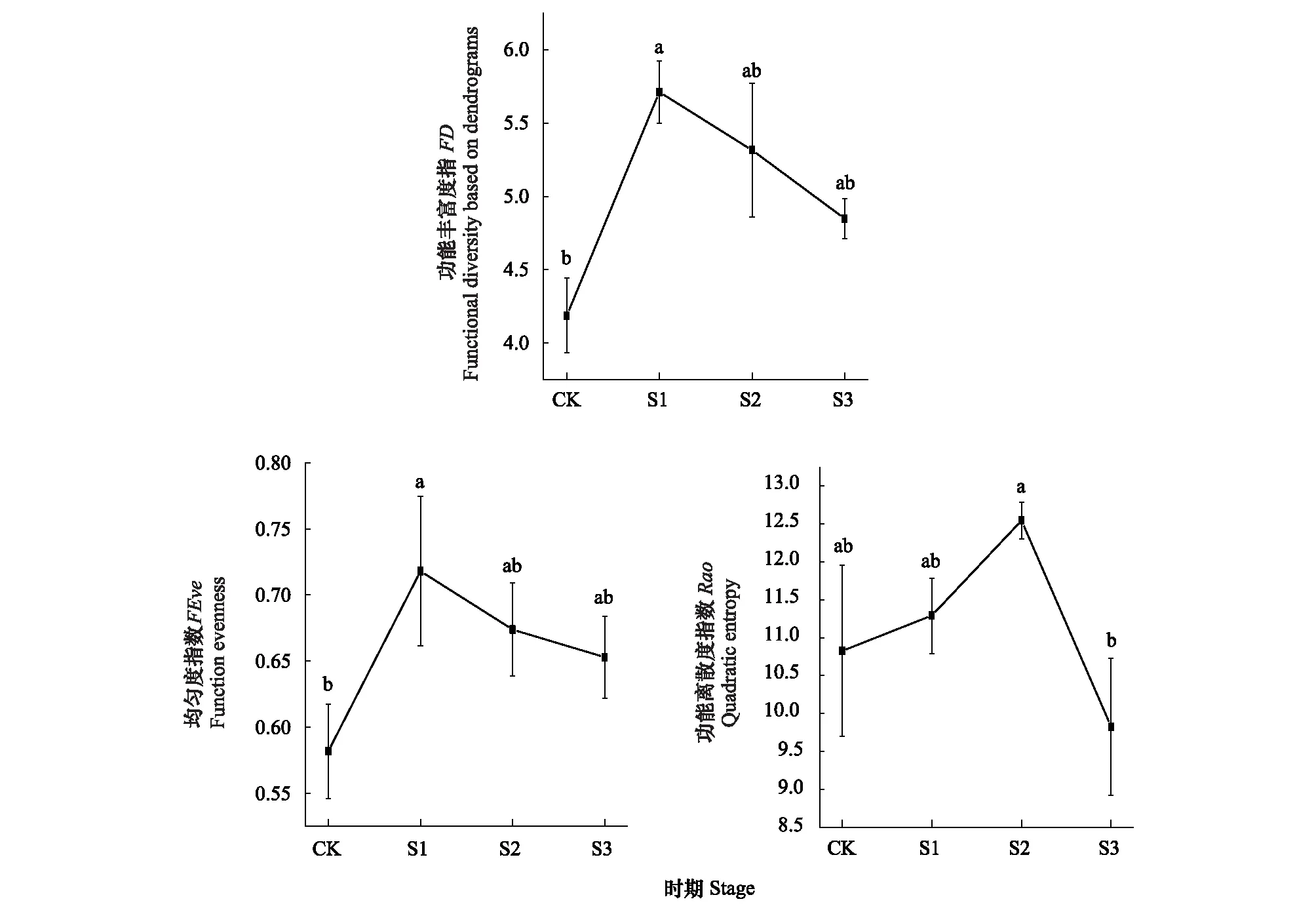
图2 综合功能多样性指数动态Fig.2 The dynamics of the comprehensive functional diversity index图中不同小写字母表示不同时期该功能多样性指数有显著差异(P<0.05)
2.2.2“光响应性状”的分类功能多样性
图3为林下植物“光响应性状”的功能多样性指数变化动态。随着森林改造的进行,功能丰富度指数FD的变化特征与林下植物综合功能多样性指数大体相同,均表现为在改造前期突增,在改造中期和后期则逐渐下降。随着森林改造的推进,“光响应性状”的功能均匀度指数FEve在改造初期不显著地高于其他3个阶段。“光响应性状”功能离散度指数Rao在改造前期出现较大的增势,改造中期增势减缓,改造后期则显著降低。
2.2.3“繁殖性状”的分类功能多样性
林下植物群落“繁殖性状”的功能丰富度指数FD的变化特征与林下植物 “光响应性状”功能多样性指数表现出相似的变化规律,在改造前期和中期维持在较高的水平,改造后期出现下降趋势。林下植物“繁殖性状”的功能均匀度指数FEve较为平稳,在4个改造阶段没有表现出显著的差异。“繁殖性状”的功能离散度指数Rao在改造中期达到峰值,显著增加,但在改造后期急剧降低。
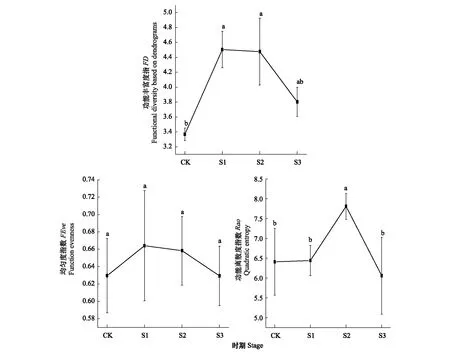
图4 “繁殖性状”功能多样性指数动态Fig.4 Dynamics of functional diversity index of “reproductive related traits”图中不同小写字母表示不同时期该功能多样性指数有显著差异(P<0.05)
3 讨论
3.1 马尾松林改造对林下物种与功能性状组成的影响
森林群落中,乔木层因其处于垂直结构的优势地位,是影响林下植物光照资源及其分布的最明显因素,这种影响主要通过乔木层的优势树种及其丰富度来实现[31- 33]。在对原马尾松林的改造过程中,随着乔木层植物格局的演变,林下植物物种和功能性状组成的相应变化也反映了乔木层的这种影响。
在改造前期,由于之前对乔木层马尾松持续的疏伐以及林下灌木层香樟幼苗的替代性抚育,导致了乔木层树木的不断减少,而短期内多数香樟幼苗来不及长大成林,乔木层对林下植物的影响较小,林下植物群落获得了较多的光照资源。这种优越的光照条件一方面促进了喜光的先锋植物和随机种的侵入,另一方面促进了灌木层植物的生长,但同时给草本层植物创造了阴生的生长环境,导致草本层阴生植物的增加,所以改造前期林下植物的种数和多度均大幅增加[34- 35]。在改造前期,林下植物“光响应性状”中,除草本层阴生植物物种和多度比例增加外,叶片形状为长条型的植物多度比例也大幅增加,这是林下植物群落对光照环境变化的一种响应[36]。在改造的影响下,“繁殖性状”也表现了相应的变化特征,其中林下植物开花期在4—6月的植物多度比例增加,而7—9月的开花植物多度比例下降,可能是因为林下较好的光照质量和强度促进了林下植物整体开花时间的提前[37]。此外,传粉方式中,风媒植物的多度比例减小,虫媒植物的多度比例增加,一般在进化上虫媒是比风媒更先进的一种传粉方式,相比风媒,虫媒更有利于促进种群分布的扩散[38]。在改造前期,林下植物性状组成的变化更多地体现在各功能性状的多度比例上,而各功能性状的物种组成比例变化不明显。
改造中期,森林群落所受干扰较小,各层次的植物处于自然生长状态,之前植入的香樟幼苗逐渐长大成林,森林群落类型由以前的马尾松针叶林转变为香樟占优势的针阔混交林,乔木层的这种转变可能导致林冠层的开度减小和林下光照环境的复杂,林下光照资源逐渐减小[39]。光照资源减小又会导致环境容纳的植物数量少,如即便是习惯阴生的植物其多度比例也大幅下降。同时林下部分复杂的光照环境,给半阴植物提供了合适的生存条件,林下半阴植物的多度比例也比改造前期增加了一倍,半阴植物的增加和随机种的入侵促进了改造中期物种数量的增加[35]。长条形叶片植物的多度比例相比前期显著降低,卵形叶片植物数量占据了多数,以及常绿植物多度比例的增加,可能都是对林下光照环境的一种响应,以便于更好的利用光照资源[36]。“繁殖性状”中,种子扩散方式和传粉方式各性状的植物多度比例也发生了较大变化,这可能是林下植物群落扩散策略的一种调整,以提高对林下资源利用效率[38]。林下植物的开花时期表现出了滞后,花期时长增加,这种生殖发育时间的延长可能与林下光照质量下降有关[40]。
改造后期,群落已基本更替为成熟的香樟林,乔木层树木相对更为高大和密集,冠层结构变复杂,透光性进一步降低,经过一段时间的物种竞争后,部分物种被淘汰,此阶段林下植物种类和数量下降,但也相对提升了林下植物群落对环境的适应能力[34,41]。“光响应性状”中,阴生植物多度比例增多,阳生植物多度比例减小,常绿植物多度比例增加,落叶植物多度比例减小,这些变化都体现了对光照资源减少的响应,因常绿植物的叶片大多为革质,所以叶片为革质的植物多度比例也随着增加。“繁殖性状”中,种子扩散方式和传粉方式等功能性状组成的变化,体现为林下植物群落扩散范围减小的特征,可能是林下资源分布不均引起[23,38]。开花期和花期时长的变化则显示了生殖发育时间延长的趋势,也是对光照资源减小的一种响应表现。
因而,马尾松林改造对林下植物的影响机制可能是,乔木层的演变格局导致林下光照资源分布变化,进而影响林下植物的生长、发育和扩散。在经过林下植物对环境的响应变化和生物竞争等共同作用下,林下植物群落物种种类、多度和空间分布格局发生改变,并最终影响生态系统功能。
3.2 马尾松林改造对林下植物功能多样性的影响
功能多样性被认为可能是生物多样性影响生态系统功能的主要机制,其可以指示生态系统或群落的生产力大小、资源动态和稳定性等变化特征[3,18-19]。基于目前还没有一个完善的测定植物功能多样性的指标,现有的每一种指数反应的生态系统过程其侧重点也不一样,因此在进行功能多样性研究中,选用多种指数,从不同角度进行研究可以起到比较好的互补作用[17,42]。
功能丰富度反映生态位空间被现有物种占据情况,丰富度越高,生态位空间越被充分占据,群落生产力越强,生态系统功能也越稳定[32]。功能丰富度一般与物种丰富程度成正比,当性状随机分布时,物种越多,它们所占据的性状空间也越大[2,33]。林下植物综合功能丰富度指数和“光响应性状”与“繁殖性状”两种分类功能丰富度指数FD的大小在改造不同时期的排序均为:S1>S2>S3>CK,这与林下光照资源优势情况大致吻合。两种分类功能丰富度变化的差异主要体现在改造中期,此时“光响应性状”的功能丰富度指数FD减小的比例较大,可能是此时林下光照资源下降引起,而“繁殖性状”的功能丰富度指数变化较小,可能与此时物种数目的增加有关,也可能反映光照的变化对“光响应性状”功能丰富度的影响先于“繁殖性状”。同时,马尾松林经改造后的各个时期林下植物功能丰富度指数均高于对照组,整体上反映了改造后林下植物占据了更多的生态位空间,群落生产力增加,生态系统功能更稳定[10,13]。
功能均匀度衡量物种性状在已占据的性状空间内的分布情况,指示群落对资源的利用程度。功能均匀度高时,群落性状的分布较规律和均匀,群落对资源的利用也较充分;低则说明群落中物种及其多度在性状空间内呈分散集群状态,同时说明某些资源利用过度,而其他资源利用较少或没被利用[4,30]。改造过程中,林下植物综合功能均匀度指数的变化趋势与综合功能丰富度相同,一定程度上反映了功能均匀度的与功能丰富度相同的响应机制,主要受到了林下光照环境的影响。而从“光响应性状”的功能均匀度FEve的表现中,可以推断其受到了光照资源分布的一定影响。改造前期,林下光照资源相对最丰富,因而群落物种扩散较为均匀,功能均匀度最大,资源的利用效率高;改造中、后期,乔木层群落开始由针叶林转变为阔叶林,林下光照资源大幅减小,光照环境发生变化,林下植物倾向于依不同生境集群生长,如群落边缘地带物种较丰富,而群落内部分阴生环境物种较少等,造成功能性状分布不均,功能均匀度降低。“繁殖性状”的功能均匀度变化幅度较小,可能说明其对林下光照资源变化的响应相对滞后[38]。
功能离散度描述植物性状在群落性状空间的离散程度,功能离散度高时物种簇分布在性状空间的边缘。功能离散度在一定程度上反映了群落植物形态和生存策略[4,30]。林下植物综合功能离散指数和“光响应性状”与“繁殖性状”两种分类功能丰富度指数Rao的大小在改造不同时期的排序均为:S2>S1>CK>S3。林下植物整体功能离散度较高的S2和S1时期,物种的种类和数量相对较多,而功能离散度小的CK和S3阶段物种的种类和数量较少,这反映了功能离散度和物种的种类以及数量具有一定的关联性。改造中期林下植物群落功能离散度最大,可能是由林下光照资源分布不均导致的生境分化和物种数量的增加而引起,使物种簇在性状空间的离散程度增加;而在改造前期“光响应性状”和“繁殖性状”的功能离散指数变化趋势差异最大。结合两者之后的变化趋势,说明了林下植物“光响应性状”对林下光照资源变化的响应比“繁殖性状”更为敏感,在森林改造的更早阶段即发生响应。而改造后期的功能离散度最小,除了林下物种的种类和数量较小的原因外,可能还受种间竞争的影响,最后剩下来的物种相似程度高,所以功能离散度较小,这样的变化可能更有利于生态系统功能的稳定[42]。
从以上研究中可以发现,改造过程中林下植物“光响应性状”和“繁殖性状”的功能性状及其多样性变化趋势相似,但前者比后者对改造的响应更敏感。将各个时期进行比较,林下植物功能多样性的综合表现大致为:改造前期(S1)>改造中期(S2)>改造后期(S3)>未改造时期(CK)。次生马尾松纯林在经改造后,林下植物的功能多样性大小的动态变化过程,也与前人的报导中,间伐改造对林下植物的影响呈先增大后减小的趋势契合[13]。
从功能多样性的角度来看,马尾松向香樟林的改造加速了森林群落演替的进程,提升了群落的整体功能。在森林改造过程中,对乔木层马尾松的间伐,香樟树种的植入,以及随机树种的入侵,均相对改善了乔木层的林分,实现了由针叶林向阔叶林的转变。随着森林改造时间的推移,乔木层物种数目的增多使群落更加复杂和稳定,提升了群落功能[10]。同时,林下植物的组成、种内种间关系对乔木层的这种变化及其环境影响产生积极响应,其结构和功能趋于新的稳定[13,43]。但研究结果显示,在森林改造的后期出现了林下植物功能多样性降低等问题,这些迹象可能反映改造后期乔木层树木对林下植物存在较为严重的干扰作用。本研究中,改造时在林下种植的香樟密度为2500株/hm2,大于前人研究所认为的合理的香樟种植密度1111—1667株/hm2[44],较高的种植密度可能会增加后期阶段乔木层对林下植物的干扰,这可能是改造后期林下植物功能多样性下降的一个重要原因,应适当调整造林密度。此外,在改造模式的选择上,纯林化改造一般是比混交林改造低效的一种方式[45]。相比纯林,在乔木层选择合适的树种混交改造可以更好利用环境,有利于提升群落整体功能[46-47]。而本文中出现的林下植物的功能多样性在改造后期阶段出现降低等问题则可能是香樟纯林化改造的一种缺陷。在对马尾松次生纯林进行改造时,应提倡选用混交林的改造模式。
4 结论
马尾松次生纯林改造成香樟林过程中,林下植物的功能多样性总体上表现为先增加后减小的趋势,反映了林下植物功能性状对乔木层物种的变化所引起的林下环境变化的积极响应。其中,林下植物“光响应性状”对改造的响应变化比“繁殖性状”更为敏感。马尾松次生纯林向香樟林的改造加速了群落的演替进程,提升了群落功能,但仍存在后期林下植物功能多样性降低等问题,在改造时应尽量选择合适的造林密度和混交林的改造模式。
猜你喜欢
农业灾害研究(2022年6期)2022-12-02
建筑与预算(2022年6期)2022-07-08
西安交通大学学报(社会科学版)(2021年2期)2021-04-06
农村实用技术(2020年1期)2020-12-16
绿色科技(2019年5期)2019-11-29
北方音乐(2019年6期)2019-07-16
中国粮油学报(2018年12期)2018-03-19
现代园艺(2018年14期)2018-01-18
现代园艺(2017年23期)2018-01-18
小小说月刊·下半月(2015年12期)2015-05-14
