
草莓维生素C合成调控机制研究进展
2020-09-08 02:24褚晓洁赵儒楠查笑君高用顺
中国野生植物资源 2020年8期
褚晓洁,赵儒楠,查笑君,高用顺
(1. 浙江师范大学 化学与生命科学学院,浙江 金华 321004; 2. 北京市林业果树科学研究院农业农村部华北地区园艺作物生物学与种质创制重点实验室,北京 100093;3. 南京林业大学 风景园林学院,江苏 南京 210037)
维生素C(Vitamin C, Vc)又称抗坏血酸,是一种水溶性维生素,是生物体中重要的抗氧化剂[1]和辅酶因子[2]。Vc以脱氢抗坏血酸DHA和L-抗坏血酸AsA两种形式广泛存在于植物体内,具有抵抗氧化胁迫、提高植物抗逆性等作用,在植物的生长发育过程中必不可少[3]。Vc作为一种高效抗氧化剂,参与人体多种代谢反应,是人体必不可少的营养物质和抵抗衰老的良好物质。Vc可以清除人体内产生的活性氧自由基,从而起到预防癌症、抗衰老、抗心血管疾病的作用[4-5];亦可促进胶原蛋白的合成[6],从而使人体伤口快速愈合;还具有治疗坏血病,提高人体免疫力等作用,是维持人类生命活动不可或缺的有机物质。近年来随着科技的发展,研究人员发现人与天竺鼠等动物自身不能合成Vc,而其它动物可以在肝脏或肾脏中合成自身所需要Vc[7-8]。随着科技的发展研究人员逐渐发现,由于人体缺乏L-古洛糖酸-1,4-内酯氧化酶而丧失了合成Vc的能力[9],人类只能通过外界食物获取Vc,而水果和蔬菜中富含的Vc,是人类摄取Vc的主要来源。
草莓(Fragaria×ananassaDuch.)为蔷薇科(Rosaceae)草莓属(Fragaria)的多年生草本植物[10-11],其果肉鲜美,含有特殊的浓郁芳香,是世界上最受欢迎的水果之一[12-13]。草莓属植物广泛分布于我国以及世界各地[14-15],据统计全世界现有草莓栽培品种多达2000余个,已成为全球重要的水果与经济作物。草莓的营养价值极高,风味独特,含有丰富的Vc,具有“水果皇后”之称,深受各国人民的喜爱。目前我国各地均有草莓种植,据中国园艺学会草莓分会统计,2018年我国草莓总产量达500万吨,在各农作物与水果中草莓增幅名列前茅[16]。Vc与人类健康密切相关,但是人体自身不能合成,必须从食物中获取,因此研究草莓中Vc的合成及调控机制,改良草莓组织中Vc的含量对草莓的自身生长发育、抗逆生长以及对人类健康都具有重要的意义。目前诸多植物中Vc代谢途径已逐渐清晰,而草莓中Vc合成机制尚不明确,拟对现有Vc合成机制以及草莓Vc合成研究情况加以梳理,以期为今后草莓Vc合成机制的研究提供指导。
1 植物Vc合成途径
早在20世纪初,Holst和Frölich[17]通过“喂饲实验”发现老鼠和其它动物不会患坏血病,但天竺鼠在长期不服用新鲜蔬果后会患坏血病。或许是出于对了解人类自身Vc合成途径的迫切需要,上世纪50年代,Vc在动物体内的生物合成途径已被很好地理解。随后Chatterjee[18]便发现由于人类缺乏L-古洛糖酸-1,4-内酯氧化酶而不能在体内合成Vc。迄今为止动物Vc合成途径就一直领先于植物,或许是植物体内Vc合成途径较为复杂,直到20世纪末植物体内Vc合成途径开始被较清晰地解释。到目前为止植物体内可能存在4条Vc的合成途径,即:L-半乳糖途径(D-甘露糖途径)、D-半乳糖醛酸途径、古洛糖酸途径和肌醇途径(图1)。
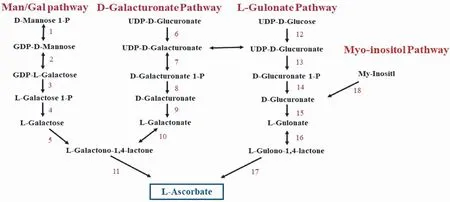
图1 Vc生物合成途径
1.1 L-半乳糖/D-甘露糖途径(L-Gal/D-Man)
该途径是Wheeler等[19]在前人研究基础上,以同位素示踪法并结合生化分析等手段提出来的,随后大量的实验对此途径展开研究与验证,目前L-半乳糖/D-甘露糖途径已是公认的高等植物Vc合成的最重要和最主要且最被接受的途径。L-半乳糖/D-甘露糖途径自葡萄糖开始,合成途径中的所有基因均已被识别[20]。Wheeler等[19]以同位素标记的L-半乳糖培养豌豆和拟南芥,发现D-甘露糖和L-半乳糖是Vc合成的有效前体,并发现在豌豆和拟南芥中L-半乳糖脱氢酶(GalDH)可催化L-半乳糖氧化成L-半乳糖酸-1,4-内酯(GalLDH)。Dowdle等[21]发现该途径是拟南芥中Vc生物合成的主要途径。Conklin等[22]通过抗坏血酸缺陷型突变体,在L-半乳糖/D-甘露糖途径中鉴定了VTC1、VTC2、VTC3、VTC4和VTC5这5个与抗坏血酸相关基因。随着编码 GDP-L-半乳糖磷酸化酶(GGP)的VTC2和VTC5基因被鉴定出来,L-半乳糖/D-甘露糖途径中所有参与基因均被鉴定出来[23-24]。
近年来L-半乳糖/D-甘露糖途径依然是植物Vc合成途径研究的热点,Yoshimura等[25]在连续光照与黑暗处理,发现VTC2过表达导致拟南芥等植物Vc含量增加。Shiroma等[26]发现当编码GGP的VTC2基因表达会导致拟南芥Vc含量增加,进而发现L-半乳糖途径也参与了拟南芥根系中Vc的合成。Qin等[27]发现GDP-甘露糖焦磷酸化酶(VTC1)的几个同源基因OsVTC1-1和OsVTC1-3参与了Vc的合成,它们可能分别参与了叶片和根系中Vc的合成。Vidal-Meireles等[28]在对衣藻Vc生物合成和基因表达调控研究中发现,GGP在衣藻Vc合成中起着关键作用,即使衣藻Vc合成调控机制与陆地植物略有不同,但仍受L-半乳糖/D-甘露糖途径高度调控。Caviglia等[29]利用两个独立的拟南芥突变体,研究乙烯在影响光合作用和生长的氧化还原信号途径中的作用,发现这两个突变体在GDP-L-半乳糖磷酸化酶合成基因中都有缺陷。黄晓婧等[30]克隆并分析了甜樱桃‘佐藤锦’果实中PacGalDH和PacGalLDH基因,分析发现在‘佐藤锦’果实生长发育过程中,PacGalDH表达量与Vc含量变化基本一致。
1.2 D-半乳糖醛酸途径
早在上世纪50年代研究人员便提出了D-半乳糖醛酸途径,但因科研手段有限,缺乏充足的证据而未被及时的证明,D-半乳糖醛酸途径来源于植物细胞壁果胶成分D-半乳糖醛,经若干酶促反应最终合成Vc,但D-半乳糖醛酸途径中仅有GalUR基因被克隆。Isherwood等[31]用D-半乳糖醛甲酯浇灌水芹,结果发现水芹Vc含量明显增加,因而推测植物Vc合成途径可能是D-半乳糖醛酸途径。Agius等[32]在对草莓Vc合成途径的研究中发现,GalUR是草莓中影响Vc合成的关键基因,随后将从草莓中克隆的GalUR在拟南芥中过表达,拟南芥Vc含量提高2~3倍,从分子机制上证实了D-半乳糖醛酸途径的存在。
Melino等[33]在葡萄果实中也检测到了GalUR基因,也进一步证实了D-半乳糖醛酸途径的存在。随后Cruz-Rus等[34]在对葡萄果实发育过程中Vc合成相关基因的表达量进行研究,发现VvGalUR基因的表达量随着Vc含量的增加而上调,且VvGalUR基因的表达受高光照的调节,因而推测D-半乳糖醛酸途径参与了葡萄果实成熟过程中Vc的合成。Jiang等[35]对猕猴桃Vc合成途径中相关基因的表达进行研究,发现猕猴桃Vc合成是由L-半乳糖、D-半乳糖醛酸等途径共同完成的,且GalDH、GMP、GPP、GGP、GalUR等基因对猕猴桃果实Vc合成具有重要的调控作用。虽然D-半乳糖醛酸途径已被广泛证实,但整个路径中的一些编码酶基因以及该途径调控机制仍需进一步研究。
1.3 古洛糖酸途径
Wolucka和Montagu[36]通过GDP-D-甘露糖-3’,5’-差向异构酶(GME)的差向异构反应,提出了古洛糖途径。该途径开始于GDP-L-古洛糖,经若干酶促反应最终生成Vc,但整个反应途径尚未被全面阐明,反应途径中的中间产物也未被充分克隆并分析。早期研究将在老鼠中克隆得到的L-古洛糖酸-1,4-内酯氧化酶转化进莴苣和烟草可显著提高转基因植株的Vc含量,基于此研究人员推测植物体内可能存在一种替代途径,它可能可以绕过L-半乳糖途径中的一些步骤直接合成Vc,这一途径可能与某些动物Vc合成途径相似[37-38]。Maruta等[39]对L-古洛糖-1,4-内酯氧化酶(L-GulL)同系物AtGulLOs在Vc生物合成中的作用研究中发现,在L-GulL处理下转基因烟草中总Vc含量与对照组相比显著升高。Lim等[40]在番茄中过表达老鼠L-古洛糖-γ-内酯氧化酶(GLOase)基因,发现转基因植株Vc含量增加了1.5倍。这些研究都说明该途径可能参与植物体内Vc的合成,而Lisko等[41]通过对拟南芥、马铃薯、烟草、番茄和水稻低Vc突变体进行鉴定,发现拟南芥过度表达含有高抗坏血酸的L-古洛糖酸-1,4-内酯氧化酶,可促进气生和根组织生长,同时Vc总含量明显增加,这一研究证实了该途径的存在。
1.4 肌醇途径
Lorence等[42]在野生型拟南芥中超表达肌醇加氧酶(MIOX),发现Vc含量明显增加,基于此提出了一个可能的生物合成路径-肌醇途径。但Endres和Tenhaken[43]发现野生型和MIOX高表达拟南芥之间Vc生物合成没有差异,因而推测MIOX对Vc的生物合成所起作用不大。Zhang等[44]从转录水平和酶活性两个方面研究了缺氮条件下黄瓜幼苗抗Vc合成和循环途径中的关键酶,发现缺氮诱导的基因参与了黄瓜叶片中Vc合成的肌醇途径等,且MIOX可能在Vc的积累中起作用。因而该途径的有效性尚未被充分证实,该途径还需要更多的证据来支撑。虽然MIOX在Vc合成中的作用仍有待验证,但MIOX在提高植物抗逆性中的作用也引起了研究人员的兴趣,或许肌醇途径并不只参与Vc的合成。Chen等[45]利用实时定量PCR技术,发现GsMIOX1a主要在花中表达,且GsMIOX1a基因正调节植物对碱胁迫的耐受性。
2 草莓Vc合成途径内关键基因
2.1 D-半乳糖醛酸途径与GalUR
20世纪初Agius等[32]首次克隆了GalUR基因并证实了D-半乳糖醛酸途径与GalUR的存在,由于当时在草莓中尚未发现L-半乳糖途径,所以研究人员认为D-半乳糖醛酸途径很有可能是草莓中Vc合成的主要途径。但到目前为止该途径仅有GalUR基因被克隆,且GalUR的作用机理还未被完全阐明。早期研究发现D-半乳糖醛酸途径可能在成熟果实的Vc合成中起作用,Cruz-Rus等[46]研究发现不同品种草莓倍性与Vc含量均明显不同,但Vc含量与倍性关系不大。同时对不同草莓品种GalUR的研究表明,GalUR与Vc含量的相关性在大多数情况下是明显的。此外FaGalUR在草莓果实不同成熟期表达量显著不同,这预示着在草莓果实不同发育阶段调控Vc合成的途径可能是不同的。草莓中D-半乳糖醛酸途径的研究仍处在起步阶段,该途径中的各种中间产物也尚待发掘,GalUR在不用果实发育阶段与不同倍性间的作用机制仍需进一步揭示。
目前D-半乳糖醛酸途径的存在与否在草莓中得到了充足的证明,但该途径在其他植物中是否发挥着同等重要的作用尚待进一步研究。Lim等[47]将GalUR基因转化番茄发现部分转基因番茄Vc含量明显增加,同时转基因植物后代由于Vc含量的增加也上调番茄的抗氧化系统,导致番茄对抗盐等非生物胁迫的耐受性增强。Badejo等[48]研究发现在番茄绿色的果实中主要是L-半乳糖途径调控其Vc的合成,而在成熟果实中D-半乳糖醛酸途径则发挥着主导作用。这不仅说明D-半乳糖醛酸途径不仅在番茄果实Vc合成中发挥着重要作用,也说明不同的植物器官或不同发育阶段或许是不同的合成途径参与Vc的积累。这一推测是否合理尚待证实,同时这也提醒我们在草莓叶片与果实间甚至是果实不同成熟阶段间,可能是不同合成途径参与了Vc含量的积累。此外GalUR在草莓等植物抗逆性中发挥的作用亦不容忽视,这为今后的研究方向也提供了一定借鉴意义。
2.2 L-半乳糖途径与VTC2
早期通过转基因手段提高植物Vc含量多在模式植物如拟南芥、烟草等植物的叶片中展开,同时针对一些水果、农作物如番茄、柑橘、马铃薯、猕猴桃的研究也在有序展开,而草莓Vc合成途径研究却相对迟缓。L-半乳糖途径是目前植物中Vc合成中最重要和最主要的路径,而早期草莓中Vc合成途径仅发现了D-半乳糖醛酸途径。Bulley等[49]利用GDP-L-半乳糖磷酸化酶(GGP/VTC2)转化5个番茄系、9个马铃薯系和8个草莓系,发现所有草莓品系果和叶的Vc含量都增加了两倍,这也似乎证实草莓叶与果实的Vc合成均有L-半乳糖途径的参与。
目前研究发现VTC2在草莓Vc合成中占据重要地位,且该基因似乎是受光照调控的。Dowdle等[21]研究发现L-半乳糖途径是拟南芥幼苗Vc的唯一重要来源,同时发现VTC2基因编码的GDP-L-半乳糖磷酸化酶可将GDP-L-半乳糖转化为L-半乳糖1-P,并发现VTC2基因似乎与光照条件密切相关。Fukunaga等[50]研究发现水稻中Vc的合成受光的调控,且L-半乳糖-1-磷酸磷酸酶(GPPase)、L-半乳糖酸-1,4-内酯脱氢酶(GalLDH)的启动子区域含有光顺式元件。Qin等[27]发现GDP-甘露糖焦磷酸化酶(VTC1)的两个同源基因OsVTC1-1和OsVTC1-3参与了Vc的合成,前者受光诱导表达,而后者受黑暗诱导表达。但因L-半乳糖途径需要利用光合作用产生的葡萄糖进而合成Vc,光照影响VTC2的表达,也极有可能是光照影响了植物自身的光合作用,L-半乳糖途径Vc合成前体的减少,必然影响Vc的合成,因此VTC2在草莓中的光响应机制及作用机理尚有待进一步研究。
此外Baig等[51]给豆角和草莓浇灌L-古洛糖酸-1,4-内酯,发现它们可将该化合物转化为Vc,使Vc含量明显提高。这似乎暗示草莓Vc合成途径中可能存在古洛糖酸途径,但迄今为止尚未有研究人员对此展开研究,所以古洛糖酸途径在草莓Vc合成中是否发挥着作用我们是不得而知的。但就目前而言,研究人员在草莓Vc合成中仅发现了D-半乳糖醛酸途径和L-半乳糖途径(图2)。
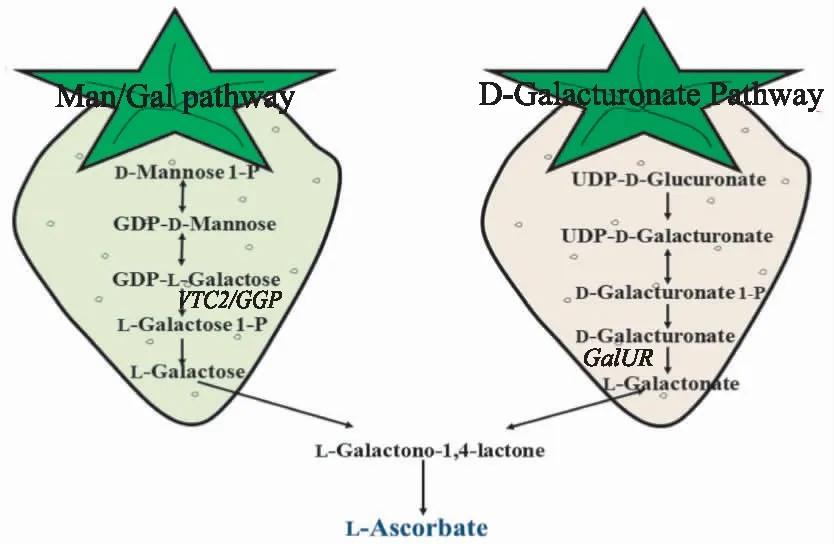
图2 草莓中Vc 生物合成途径
3 展望
目前国内外对于草莓Vc合成调控机制的研究明显落后于拟南芥、烟草、水稻等模式植物,甚至柑橘、番茄、猕猴桃等农作物或水果的研究也明显领先于草莓。最早在草莓中鉴定的D-半乳糖醛酸途径也仅有GalUR被克隆,该途径的具体机制尚未完全明确,甚至GalUR的作用机制也未被完全阐明。目前研究发现D-半乳糖醛酸途径可能在果实发育中起作用,然而在草莓不同倍性、果实不同发育阶段以及草莓叶片和果实中,究竟哪种途径占据主导地位,或是它们间是否存在不同的调控机制,这些都值得进行深入研究。
在草莓中鉴定出了D-甘露糖途径,然而却少有研究者对此展开探索与讨论。现有研究发现D-甘露糖途径主要在植物叶片及青果期发挥作用,然而该途径是否在草莓叶片及草莓青果期中占主导地位还值得深入研究。同时VTC2在草莓Vc合成中占的重要地位也逐渐被发掘,尤其是该基因受光调控现象的发现,也为该路径与该基因的研究提供了新思路。此外GalUR与VTC2可调控Vc合成而在提高植物抗逆性方面表现出独特的优越性,随着现代分子育种技术的兴起,利用优异基因提高水果、作物营养品质及抗逆性具有重要意义。但目前对这两种酶的研究尚不深入,尤其是遗传方面的研究还比较少,调控其表达的转录因子方面研究也存在着一定缺失。
猜你喜欢
食品与生物技术学报(2022年1期)2023-01-11
中草药(2022年6期)2022-03-21
江苏农业学报(2022年1期)2022-03-16
河北果树(2021年4期)2021-12-02
发酵科技通讯(2021年1期)2021-03-18
家庭百事通·健康一点通(2020年12期)2020-12-31
食品与发酵工业(2020年16期)2020-09-03
山东农业科学(2020年1期)2020-07-04
分析化学(2017年12期)2017-12-25
医学美学美容·中旬刊(2015年1期)2015-10-21
