
不同类型接穗银杏苗生长和光合特性的比较研究
2020-09-28 09:00吴孝红汪贵斌曹福亮郁万文
中南林业科技大学学报 2020年9期
吴孝红,任 重,汪贵斌,曹福亮,郁万文
(南京林业大学 a.南方现代林业协同创新中心;b.江苏省农业种质资源保护与利用平台,江苏 南京 210037)
银杏Ginkgo biloba属于银杏科银杏属,集药用、经济和观赏等价值于一体的植物,具有极高的开发价值。银杏幼苗有生长缓慢和优良品种的选育等问题,一定程度上影响了银杏产业的发展,而嫁接技术的应用则可在一定程度上缓解这一问题[1]。嫁接可以保持和发展银杏的优良种性,实现早期丰产和提高果树抗逆境能力,有利于提高产量等。1991年,Del Tredici[2]首次提出银杏的位置效应概念,即银杏无性繁殖的斜向生长现象。1998年,邢世岩等[3]研究发现,银杏嫁接成活后,由于接穗取自母树不同部位和树龄等原因,造成嫁接苗新梢的生长状况和角度不同,进而严重影响到嫁接苗的质量。因此,在银杏嫁接育苗中,会根据目的树形选取不同类型接穗。基于此,对不同类型接穗的嫁接苗进行光合性能的研究具有一定的意义。
光合指标和叶绿素荧光参数能够很好地反映植物的光合能力,在植物生长发育及良种选育等研究中具有重要参考价值。因此,研究砧木或接穗对银杏嫁接苗光合作用的影响具有重要的意义。目前,研究表明嫁接苗的生长发育除受环境因子影响之外,还受接穗和砧木各自遗传特性及砧穗互作效应的影响。优良砧木对嫁接苗的叶绿素含量、光合作用、生长势等有显著的提升[4-5],同时对嫁接苗非光化学淬灭的降低、光能的吸收和利用率均有显著作用[6-7]。李小飞等[8]对美国山核桃的研究发现,砧木年龄越大,嫁接苗光合能力越强。同时,大量研究表明,接穗和砧木之间存在遗传物质的相互交换,如叶绿体基因组在砧穗间的相互传递[9],接穗与砧木的基因互作会形成可遗传性状等[10]。因此,不同类型接穗会由于自身遗传特性及与砧木互作效应影响嫁接苗的光合特性。有关接穗对嫁接苗生长发育和光合特性影响的研究较少,任平等[11]发现不同长势接穗对番茄嫁接苗相关光合基因的表达及光合性能有显著的影响,进而影响植株生长发育。而未曾见有关银杏不同类型接穗对嫁接苗的光合作用与叶绿素荧光参数等方面的研究报告。因此,本试验研究了接穗来自银杏实生树不同部位和年龄的嫁接苗生长指标、光合作用和叶绿素荧光参数等指标的差异,探讨不同类型接穗嫁接苗在一天中的光合能力和光合机构性能的变化,以期为选育优良银杏嫁接苗及其光合特性的研究提供理论依据。
1 材料与方法
1.1 供试材料及处理
2018年3月上旬,选取邳州市银杏种质资源圃内2棵树干基部有大量萌条的10年生和30年生状况良好的实生树作为母树,品种为‘梅核’。采集2棵母树树冠中部与树干基部状况良好的枝条作为嫁接材料,每个接穗上保留2~3个饱满芽,砧木品种为泰兴‘大佛指’3年生银杏实生幼苗,长势及大小基本一致,砧木株行距为45 cm×60 cm。
2018年3月底,在砧木高为60 cm处采用插皮法嫁接,嫁接地点为邳州市银杏种质资源圃。将接穗来源10年生母树树冠、10年生母树树干基部、30年生母树树冠和30年生母树树干基部的嫁接苗分别记为G10嫁接苗、J10嫁接苗、G30嫁接苗和J30嫁接苗,共4种嫁接苗。每个处理分为3个小区,每小区嫁接15株。2019年5月上旬,对各处理进行测定。
1.2 测定项目及方法
1.2.1 新梢生长指标的测量
每个处理每个重复中随机挑选11株,对新梢生长角度、梢长和基径进行测量。角度测量以逆重力方向为0°,使用量角器测量新梢与接穗主干的夹角;使用卷尺测量新梢梢顶至基部的长度,使用游标卡尺测量新梢距离主干1 cm处的基径。
1.2.2 光合参数日变化的测定
测量仪器为英国汉莎科学仪器有限公司的CIRAS-3光合仪,于测量日8:00—18:00,每隔2 h测定1次4种嫁接苗叶片的光合参数,每组处理随机测3株嫁接苗。光合生理参数为:净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间二氧化碳浓度(Ci)等。环境因素包括:光合有效辐射(PAR)、空气温度(Ta)、空气相对湿度(RH)、大气CO2浓度(Ca)等。
1.2.3 光响应曲线的测定
用CIRAS-3光合仪测定4种嫁接苗叶片的光响应曲线,每种嫁接苗随机选取3株进行测定。光源采用CIRAS-3光合仪内置式红蓝光,光照强度梯度设置为0、50、100、150、200、300、400、600、800、1 000、1 200、1 400、1 700、2 000 µmol·m-2s-1,温度控制在(25±0.5)℃之间。
1.2.4 叶绿素荧光参数日变化的测定
测量仪器为英国汉莎科学仪器有限公司生产的FMS-2便携调制式荧光仪,具体测量方法参照文献[12],且与光合指标的测量同时进行。
1.3 数据处理
试验所得数据的分析与作图使用Excel 2016软件,双因素方差分析与显著性差异分析使用SPSS 20.0软件。
2 结果与分析
2.1 嫁接苗新梢生长指标
双因素方差分析表明,接穗部位对嫁接苗新梢的角度和梢长均有显著影响(P<0.05),接穗年龄和二者交互作用对角度和梢长无显著影响(P>0.05),而接穗部位、年龄和二者交互作用对嫁接苗的基径均无显著影响(P>0.05)(表1)。表2表明,J10嫁接苗和J30嫁接苗的新梢角度显著小于G10嫁接苗和G30嫁接苗(P<0.05),J30嫁接苗的梢长显著大于G10嫁接苗(P<0.05),其它各处理之间梢长则无显著差异(P>0.05),4种嫁接苗之间基径亦无显著差异(P>0.05)。新梢角度从大到小依次为G10嫁接苗>G30嫁接苗>J10嫁接苗>J30嫁接苗,梢长和基径从大到小依次为J30嫁接苗>J10嫁接苗>G30嫁接苗>G10嫁接苗。

表1 接穗的年龄和部位对嫁接苗生长指标影响的双因素方差分析Table 1 Two-factor analysis of variance for the effects of scion age and location on growth indexes of grafted seedlings
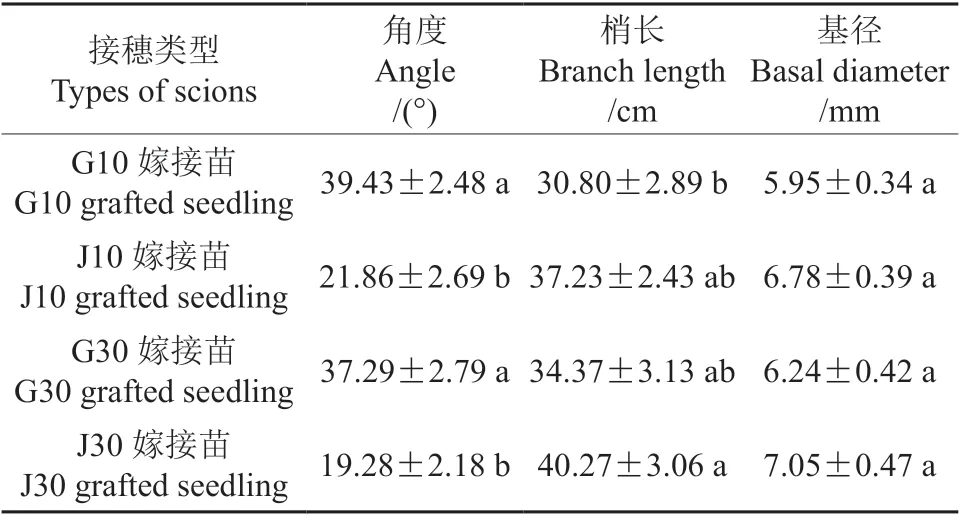
表2 接穗来自不同年龄和部位的嫁接苗的生长指标†Table 2 Growth indicators of grafted seedlings from different ages and parts
2.2 环境因子日变化
由图1可知,PAR和Ta日变化均呈单峰曲线,PAR在12:00左右出现峰值,达到1 569 µmol·m-2s-1;Ta的最大值出现在14:00,达到27.5 ℃;Ca和RH的变化呈V形,分别于12:00和14:00左右出现谷值。
2.3 嫁接苗光合参数日变化的比较
对各时间段进行双因素方差分析可知,在上午和下午,接穗部位对嫁接苗的Pn具有显著影响(P<0.05),而接穗年龄和二者交互作用对嫁接苗的Pn没有显著影响(P>0.05)。
由图2可以看出,上午,J10嫁接苗和J30嫁接苗的Pn分别显著高于G10嫁接苗和G30嫁接苗(P<0.05),12:00后则相反。同时,4种嫁接苗的Pn在12:00有大幅度的下降。J10嫁接苗和J30嫁接苗的Gs呈先下降后上升再下降的趋势,且在14:00达到谷点,16:00有一定幅度的增加,之后逐渐下降。而G10嫁接苗和G30嫁接苗的Gs在10:00有一定幅度的下降,10:00—16:00没有显著变化,在18:00有大幅度的下降。在8:00—18:00之间,4种嫁接苗的Tr均呈先上升后下降的趋势,于12:00达到最高值,且在10:00—14:00期间,G30嫁接苗和J30嫁接苗的Tr始终高于G10嫁接苗和J10嫁接苗。4种嫁接苗的Ci日变化呈W型,G10嫁接苗和J10嫁接苗2个谷值的时间为10:00和16:00,G30嫁接苗和J30嫁接苗则为10:00和14:00。

图1 光合有效辐射(PAR)、大气CO2浓度(Ca)、空气相对湿度(RH)和空气温度(Ta)的日变化规律Fig.1 Diurnal variation of photosynthetic active radiation (PAR), atmospheric CO2 concentration (Ca), air relative humidity (RH) and air temperature (Ta)

图2 接穗来自不同年龄和部位的嫁接苗光合参数的日变化Fig.2 Diurnal variation of photosynthetic parameters of grafted seedlings that scion from different age and parts
2.4 嫁接苗光响应曲线的比较
图3表明,当PAR<200 µmol·m-2s-1时,4种嫁接苗的Pn几乎呈直线上升趋势,而当200 µmol·m-2s-1<PAR<600 µmol·m-2s-1时,Pn上升趋势逐渐放缓,之后随着PAR的增加,G10嫁接苗、J10嫁接苗和G30嫁接苗的Pn变化较小,基本处于稳定状态,而J30嫁接苗的Pn则持续缓慢上升。当PAR<600 µmol·m-2s-1时,G10嫁接苗和J10嫁接苗的Pn均高于G30嫁接苗和J30嫁接苗;当PAR>600 µmol·m-2s-1时,4种嫁接苗中J30嫁接苗的Pn增长幅度最大。嫁接苗Pn在光强>1 200 µmol·m-2s-1后一直为J30嫁接苗>J10嫁接苗>G10嫁接苗>G30嫁接苗。
通过非直角双曲线模型的拟合,得到决定系数R2均大于0.99的Pn-PAR拟合曲线。同时,计算得到4种嫁接苗的最大净光合速率(Pnmax)、表观量子效率(AQY)、暗呼吸速率(Rd)、光补偿点(PLC)、光饱和点(PLS)和初始斜率(a)等相关光合作用参数(表3)。经双因素方差分析可知,接穗部位、年龄和二者交互作用均对嫁接苗的Pnmax、AQY和Rd有显著影响(P<0.05),对PLC和PLS有显著影响的因素只有接穗部位。由表3可知,接穗来自树干基部的J10嫁接苗和J30嫁接苗的Pnmax、Rd、PLS和AQY分别大于G10嫁接苗和G30嫁接苗,且J30嫁接苗与G30嫁接苗之间差异显著,J10嫁接苗与G10嫁接苗之间仅Pnmax差异显著(P<0.05)。比较接穗是同一部位不同年龄的嫁接苗,J30嫁接苗和G30嫁接苗的Pnmax、Rd、PLS和PLC分别大于J10嫁接苗和G10嫁接苗,且J30嫁接苗与J10嫁接苗、G30嫁接苗与G10嫁接苗差异显著(P<0.05)。
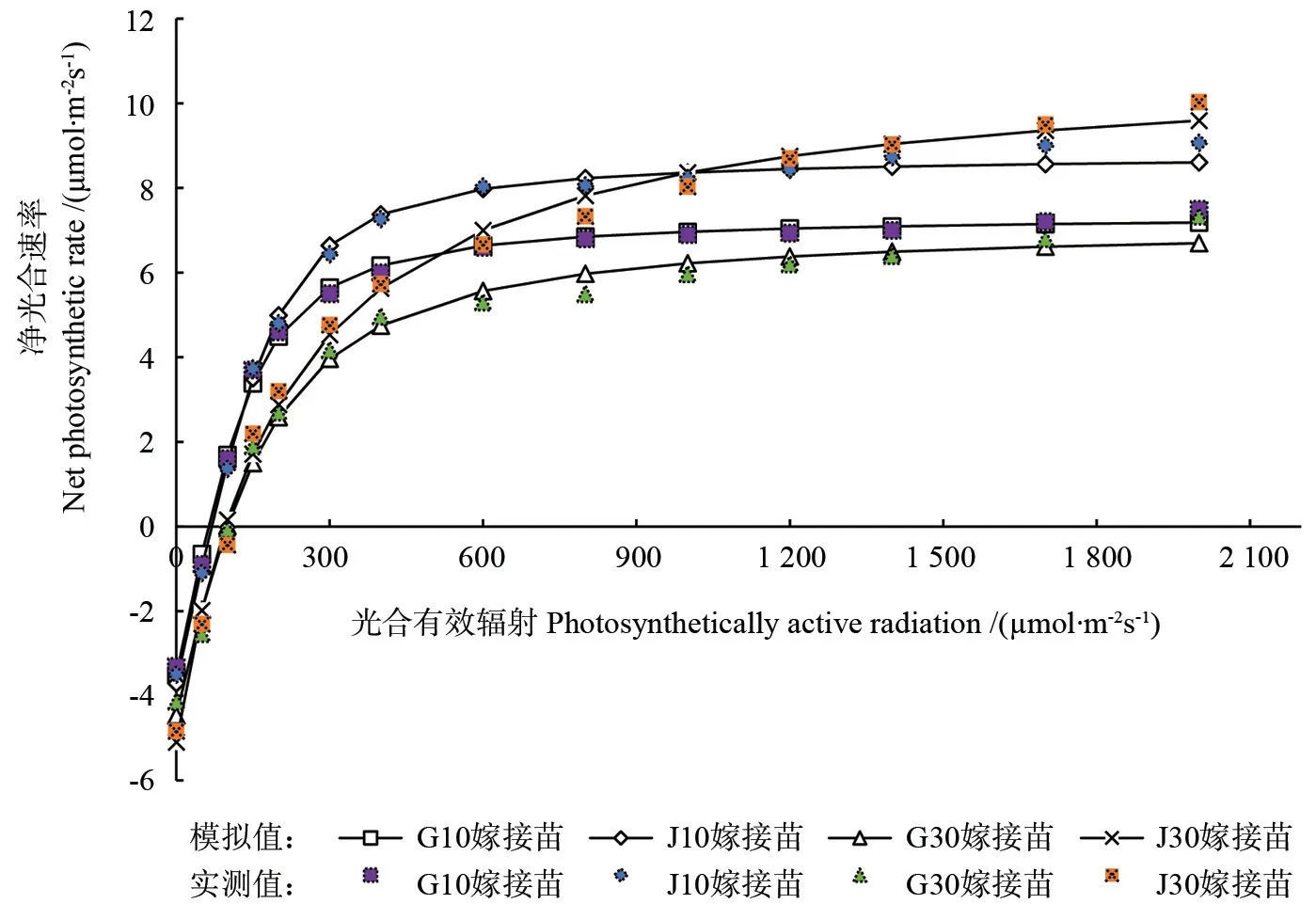
图3 接穗来自不同年龄和部位的嫁接苗Pn-PAR响应曲线的拟合Fig.3 Fitting of Pn-PAR response curves of grafted seedlings that scion from different ages and parts
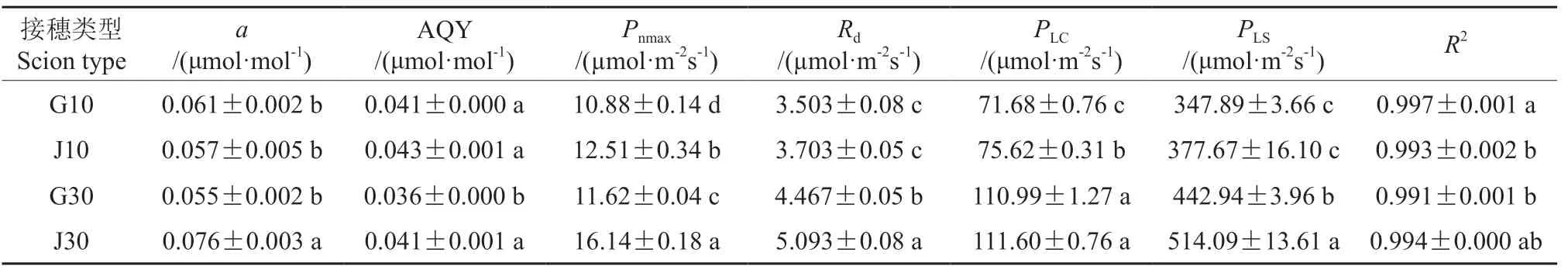
表3 接穗来自不同年龄和部位的嫁接苗光合作用相关参数†Table 3 Relevant parameters of photosynthesis of scion grafted seedlings that scion from different ages and parts
2.5 叶绿素荧光参数日变化的比较
对各时间段进行双因素方差分析可知,在8:00—12:00期间,接穗部位对嫁接苗PSⅡ最大光化学效率(Fv/Fm)和PSⅡ潜在活性(Fv/Fo)有显著影响(P<0.05),接穗年龄对非光化学淬灭系数(NPQ)和PSⅡ实际光化学效率(ΦPSⅡ)有显著影响(P<0.05),因素之间无交互作用。

图4 接穗来自不同年龄和部位的嫁接苗叶绿素荧光参数的日变化Fig.4 Diurnal variation of chlorophyll fluorescence parameters of grafted seedlings that scion from different ages and locations
由图4可知,4种嫁接苗的Fv/Fm和Fv/Fo均呈先下降后上升的趋势。上午,J10嫁接苗和J30嫁接苗的Fv/Fm和Fv/Fo均高于G10嫁接苗和G30嫁接苗,到12:00和14:00则相反,4种嫁接苗的Fv/Fm均于18:00达到最高值。4种嫁接苗的NPQ变化呈单峰型,12:00达到峰值,且此时G10嫁接苗和J10嫁接苗的NPQ高于G30嫁接苗和J30嫁接苗,傍晚达到最低值。上午,J10嫁接苗和J30嫁接苗的光化学淬灭系数(qp)分别高于G10嫁接苗和G30嫁接苗,12:00—14:00则相反,18:00均大幅度下降达到最低值。4种嫁接苗的ΦPSⅡ变化趋势均为双峰型,上午,J10嫁接苗和J30嫁接苗的ΦPSⅡ分别大于G10嫁接苗和G30嫁接苗,且两个峰值出现于10:00和16:00。
3 结论与讨论
接穗来自树干基部的嫁接苗新梢生长较为直立,接穗来自树冠的嫁接苗则为斜向生长。这种斜向生长的现象可能与银杏原始的发育系统有关[3]。同时,4种嫁接苗的新梢梢长和基径之间无显著差异,仅J30嫁接苗的新梢梢长显著大于G10嫁接苗。整体而言,4种嫁接苗的生长量在生长前期未表现出明显的差异,之后的生长受光合作用的影响。
光合作用为植物提供大量营养和干物质,对植物的生长发育及果实的产量、品质起决定性作用[13-14]。研究表明,不同类型接穗嫁接后水分和矿质营养等运输效率不同,导致接穗叶片叶绿素和营养物质等含量存在差异,也会对光合酶Rubisco小亚基和果糖-1,6-二磷酸酶等基因的表达有显著影响,进而引起光合作用的不同[11,15]。本试验结果表明,上午J10嫁接苗和J30嫁接苗的Pn更高,而午后则为G10嫁接苗和G30嫁接苗更高,表明接穗来自基部的嫁接苗在上午光合能力更强,正午后则相反,接穗来自树冠的嫁接苗抗光抑制能力强于接穗来自基部的嫁接苗。
光响应曲线反映了植物净光合速率与光强的关系[16]。参数Pnmax反映了植物叶片光合能力的强弱,Rd的大小则代表了呼吸作用对光合产物消耗的大小,AQY可作为植物对弱光利用效率的指标,其值越高对弱光能利用效率越高,而PLC和PLS为光的补偿点和饱和点,分别反映了对弱光和强光的利用效率[17-18]。本研究表明,接穗来自基部的嫁接苗的Pnmax显著高于来自同棵树树冠的,结果与4种嫁接苗净光合速率大小一致。接穗来自30年生母树的嫁接苗PLS显著高于来自10年生母树,PLC值则相反,说明接穗来自30年生母树的嫁接苗对强光利用能力更高,接穗来自10年生母树的嫁接苗对弱光利用能力更强。
叶绿素荧光是探究植物光合作用效率有效的技术手段,叶绿素荧光参数可作为反映光合内部情况的重要指标[19-20]。Fv/Fm反映了PSⅡ反应中心光能转换效率,其值越低发生光抑制程度越高,而Fv/Fo反映了PSII反应中心的潜在光化学活性[21]。本研究表明,上午接穗来自基部的嫁接苗的Fv/Fm和Fv/Fo高于接穗来自树冠的嫁接苗,正午后则相反,说明上午其光合机构活性优于接穗来自树冠的嫁接苗,而正午则为接穗来自树冠的嫁接苗光合能力更强。推测是母树树冠枝条相比基部所处环境光照更强和温度更高,导致树冠枝条正午时抗光抑制能力更强,而嫁接苗会遗传接穗生活方式[22],所以接穗来自树冠的嫁接苗在中午光合机构活性更高。NPQ反映了以热耗散形式消耗的部分光能,植物受到光抑制后通过热耗散消耗过剩激发能进行自我保护[23-25]。本试验发现,正午接穗来自10年生母树的嫁接苗比来自30年生母树嫁接苗的NPQ值更大,说明前者通过热耗散进行自我保护能力更强。qp代表光化学反应淬灭系数,数值越大说明PSII中心开放程度越大,ΦPSⅡ则代表PSII反应中心的实际光化学效率[26]。本试验发现8:00—10:00,接穗来自基部的嫁接苗的PSII中心开放程度高于来自同棵树树冠的嫁接苗,而正午后则相反。说明上午接穗来自基部的嫁接苗光合机构反应中心活性更强,正午其活性受到光抑制后显著下降。而接穗来自基部和树冠的嫁接苗的叶绿素荧光参数的差异及日动态变化,推测是由砧穗互作效应、接穗遗传特性和环境等因素共同导致。目前,有关接穗对嫁接苗的光合机构影响机理尚不清楚,有待进一步研究。
综上所述,接穗来自树干基部的嫁接苗的新梢生长更直立,其光合能力在上午优于接穗来自同棵树树冠的嫁接苗,午间则相反,接穗来自30年生母树的嫁接苗对强光利用能力更强,接穗来自10年生母树的嫁接苗对弱光利用能力更强。
虽然本研究发现不同类型接穗嫁接苗生长和光合特性等方面有明显差异,但由于影响嫁接苗生长和光合特性的因素有砧木特性、接穗特性、砧穗互作效应和环境因子等多种因素,未能验证具体是什么因素导致及哪种因素起主要影响作用。因此,在未来研究不同接穗对嫁接苗生长时,可以从接穗遗传特性、砧穗之间相关遗传物质的互作和环境因子等方面进行实验设计来探究原因。
猜你喜欢
中国蔬菜(2022年6期)2022-06-22
农业与技术(2021年8期)2021-05-13
河南农业科学(2021年1期)2021-03-09
小小说月刊(2019年12期)2019-12-16
小小说月刊(2019年23期)2019-11-14
热带农业科学(2017年9期)2017-10-23
农家顾问(2016年4期)2016-11-19
农村农业农民·B版(2016年7期)2016-10-21
意林(2009年12期)2009-02-11
