
古田山与中国其他东部典型常绿阔叶林物种组成特征的比较
2020-10-20 04:24张田田王璇金毅余建平任海保钱海源陈声文马克平于明坚
广西植物 2020年8期
关键词:多样性
张田田 王璇 金毅 余建平 任海保 钱海源 陈声文 马克平 于明坚

摘 要:古田山国家级自然保护区保存着大面积低海拔典型常绿阔叶林,为了解不同地点常绿阔叶林物种组成的差异和驱动因素,該研究在古田山常绿阔叶林内共设置29个20 m × 20 m的样地,收集其他14个典型常绿阔叶林的数据,探究古田山常绿阔叶林的物种组成特征,并将15个亚热带东部典型常绿阔叶林进行比较,分析了我国东部典型常绿阔叶林的物种组成差异及驱动因素。结果表明:(1)古田山广泛分布有我国东部典型常绿阔叶林中的栲类林、青冈类林等植被,物种组成在东部典型常绿阔叶林中具有代表性。(2)我国东部典型常绿阔叶林间物种组成差异明显,主要是受环境因素影响而非地理距离,气候因素中最冷月最低温(MTCM)的影响最显著。(3)我国东部典型常绿阔叶林叶生活型组成受最冷月最低温的影响最显著,受最热月最高温(MTWM)的影响是其次,降水量没有显著影响。综上结果发现,生境过滤是我国东部典型常绿阔叶林物种组成差异的主要驱动因素,反映了极端低温对我国亚热带常绿阔叶林物种组成具有重要影响,为常绿阔叶林的保护提供了科学依据。
关键词:中亚热带, 东部典型常绿阔叶林, β多样性, 叶生活型, 生境过滤
中图分类号:Q948
文献标识码:A
文章编号:1000-3142(2020)08-1061-10
Abstract:Gutianshan National Nature Reserve (GNNR) preserves a large expanse of low elevation typical EBLFs. In order to understand how species composition of evergreen broad-leaved forests (EBLFs) vary at the regional scale and the underlying drivers, we investigated species composition of 29 20 m × 20 m plots in GNNR, and collected data of other 14 typical EBLFs that distributed across subtropical eastern China, then we investigated the species composition difference between 15 typical EBLFs, as well as how geographical and physical environments drove these differences. The results were as follows:(1) GNNR EBLFs composed a variety of community types, such as Castanopsis forest and Cyclobalanopsis forest etc. Species composition of GNNR EBLFs was representative in subtropical eastern China. (2) Species composition (β diversity) largely differed between EBLFs. Physical environments, instead of geographical distance, was the most influential factor of EBLF species composition. EBLF species composition was the most influenced by the minimum temperature of the coldest month (MTCM). (3) Composition of leaf life forms of 15 typical evergreen broad-leaved forests was the most strongly influenced by the minimum temperature of the coldest month (MTCM), then by the maximum temperature of the coldest month (MTWM). These results indicate that habitat filtering is the main driver of the difference of woody plants species composition among eastern typical EBLFs, and suggest cold temperature is the most important determinant of eastern typical EBLFs communities, would provide guidance for conservation and restoration of EBLFs.
Key words:mid-subtropics, eastern typical evergreen broad-leaved forests, β diversity, leaf life form, habitat filtering
亚热带常绿阔叶林是世界上主要的森林植被类型之一,主要分布在中国(吴征镒,1980;Jin et al., 2018),以中亚热带低海拔地区分布的常绿阔叶林最为典型,为该地区的地带性植被类型。我国常绿阔叶林的分布范围广,研究者们已在不同地区、不同尺度开展了一些相关研究,如Wang et al.(2011)研究了我国木本植物分布格局,宋永昌等(2015)比较了我国常绿阔叶林分布区8个大型森林动态监测样地的群落特征差异,王希华(2006)研究了我国典型常绿阔叶林物种多样性分布格局。在较小尺度,各地区的研究者对当地有代表性的常绿阔叶林开展了研究。
生态位理论认为群落物种组成主要受环境因素和生物间相互作用的影响,中性过程强调扩散限制对群落物种组成有决定性作用,有研究證明生态位过程和中性过程共同影响群落的生物多样性,二者对群落构建的相对重要性随研究地点和尺度而异(Soininen et al., 2007;牛克昌等,2009)。一般认为,空间尺度越大生态位过程对物种的影响越大,空间尺度越小中性过程的影响越大(Garzon-lopez et al., 2014;Chase, 2014),如我国温带落叶阔叶林之间物种组成差异主要受环境过滤的影响(Liu et al., 2015),在区域尺度上秦岭西部的物种组成受扩散限制的影响更大(赵鸣飞等,2017)。
我国常绿阔叶林以常绿乔木和常绿灌木为主,同时包含有落叶成分(宋永昌等,2015)。常绿物种与落叶物种有着不同的生存策略,植物的落叶现象是植物对季节性干旱、低温气候等不利条件的适应性变化(Wolfe, 1987)。常绿植物和落叶植物对气候变化有不同的响应机制,二者的相对优势度体现群落外貌,是植物群落的重要特征,也是植被类型的分类依据(白坤栋等,2010;Zhang et al., 2010;宋永昌,2016;陶旺兰等,2018)。常绿阔叶林通常指热带以外由常绿阔叶树种占优势的湿润森林,因气候带和地貌等差异,根据优势或指示层片划分为不同植被亚型,其中典型常绿阔叶林分布于中亚热带温暖湿润地区,外貌终年常绿,树冠整齐(宋永昌,2013)。
东部典型常绿阔叶林是典型常绿阔叶林在我国东部中亚热带地区广泛分布的植被亚型(宋永昌,2013)。目前,关于东部典型常绿阔叶林的研究多集中在区域尺度或亚热带较大尺度的比较,而针对我国东部典型常绿阔叶林这一常绿阔叶林亚型的研究却较少。本研究比较了我国亚热带15个有代表性的东部典型常绿阔叶林,探究它们之间物种组成的差异及其驱动因素,并分析气候等因素对常绿植物和落叶植物物种丰富度的影响,以期了解我国亚热带东部典型常绿阔叶林物种多样性格局及其维持机制,为合理保护常绿阔叶林提供科学依据。
1 材料与方法
1.1 研究区概况
我国东部湿润区典型常绿阔叶林(103°—123° E,23°—33° N)分布广泛,北至秦岭、南至云开大山、西至云贵高原、东至东海海岸,位于亚热带季风气候区,夏季湿热,冬季干冷,年温差大,无明显干湿季,年均温度15~18 ℃,年均降水量1 000~2 000 mm(宋永昌,2013)。
古田山国家级自然保护区(118°03′49.7″—118°11′12.2″ E,29°10′19.4″—29°17′41.4″ N)位于浙江省开化县境内,地处黄山、怀玉山、白际山脉交汇处,面积8 107 hm2。古田山处于中亚热带湿润季风气候区,年均温15.3 ℃,最热月均温28.9 ℃,极端最高温38.1 ℃,最冷月均温4.1 ℃,极端最低温-6.8 ℃,年均降水量1 963.7 mm,年均降雨约140 d,无霜期约250 d(于明坚等,2001)。土壤类型有红壤、红黄壤、黄红壤及高山草甸土,由于地形复杂,形成了多种小气候和植被类型(楼炉焕和金水虎,2000;胡正华等,2003),分布有中亚热带地区少见的大面积低海拔常绿阔叶林(于明坚等,2001)。
1.2 常绿阔叶林调查及数据收集
在古田山保护区的常绿阔叶林中选择有代表性、典型性的地点,共设置29个样地。在20 m × 20 m 样地中央位置记录经度、纬度和地形因子,将每个样地完全划分为16个5 m × 5 m小样方,以1 cm为起测胸径(离地1.3 m主干直径),对木本植物进行每木调查,并记录种类、坐标、胸径和植株高度。
搜集整理我国东部典型常绿阔叶林样地调查资料,选择东部典型常绿阔叶林分布范围内不同地区的14个常绿阔叶林群落(表1)。基于以下原则选择样地:(1)无明显干扰历史,能体现地带性植被的特点;(2)具有地理坐标记录,以提取气候因子;(3)为减少取样面积效应,样地面积不小于20 m × 20 m(宋永昌,2016)。为更全面描述地区植被,整合同一保护区内不同样地调查资料。物种学名经校对、同物异名校正,统一参考《中国植物志》(http://frps.iplant.cn/)。本文的15个常绿阔叶林两两之间最近距离为101 km(古田山和黄山),最远距离为1 804 km(天童山和瓦屋山)。
1.3 气候数据
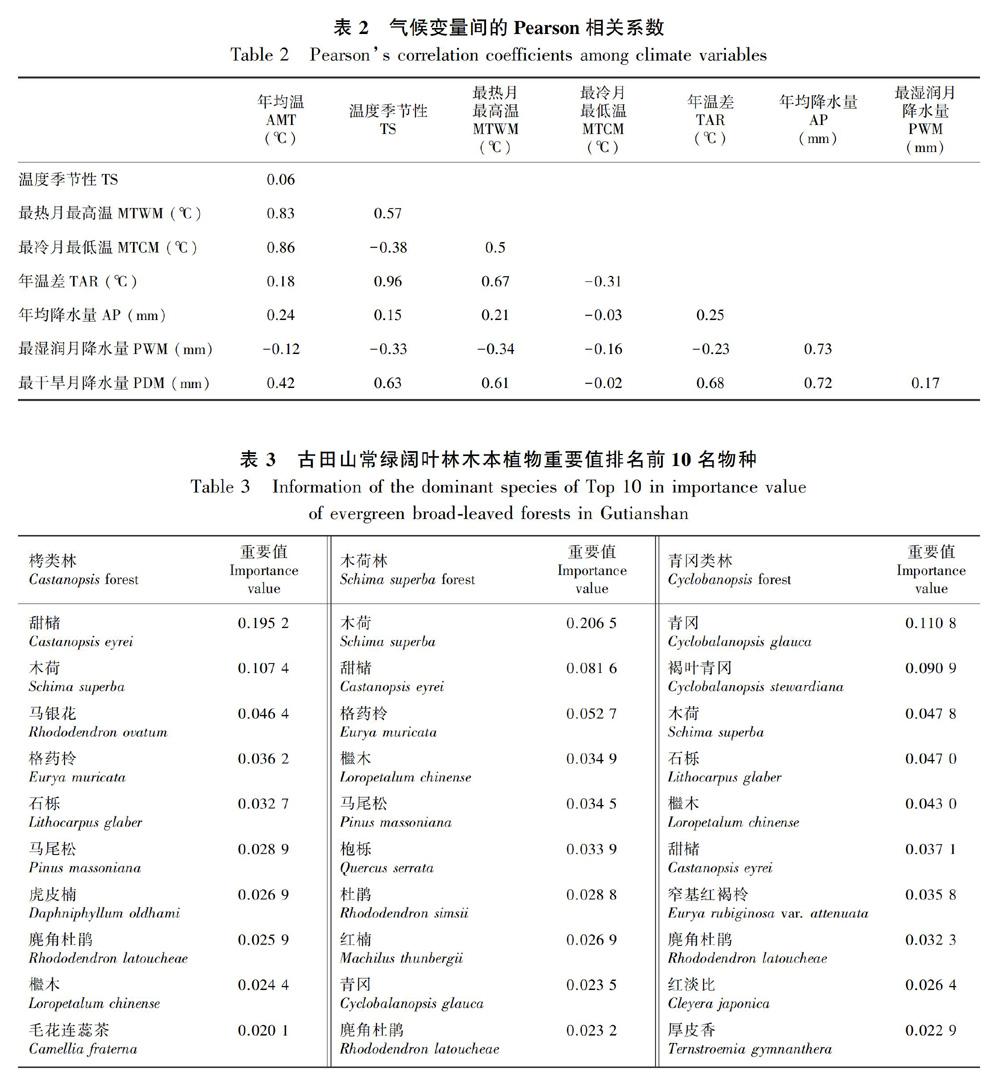
通过DIVA-GIS软件(http://www.diva-gis.org/)从WorldClim(http://www.worldclim.org) 数据库中提取气候数据(Hijmans et al., 2005),分辨率为30角秒(arc-second)。本文选择8个气候变量作为环境变量,分别为年均温(annual mean temperature,AMT)、最热月最高温(maximum temperature of the warmest month,MTWM)、最冷月最低温(minimum temperature of the coldest month,MTCM)、年均降水量(annual precipitation,AP)、最湿润月降水量(precipitation of wettest month,PWM)、最干旱月降水量(precipitation of driest month,PDM)、温度季节性(temperature seasonality,TS;即标准差*100)、年温差(temperature annual range,TAR;即MTWM-MTCM)。15个常绿阔叶林群落分布范围为103~121° E、24~30° N。年均温12~19 ℃,温度季节性606~887,最热月最高温24.5~33.8 ℃,最冷月最低温-1.6~5 ℃,年温差25.6~32.6 ℃,年均降水量1 088~2 062 mm,最湿润月降水量164~367 mm,最干旱月降水量11~51 mm。由于部分气候变量之间有明显的相关性(表2),因此通过方差膨胀因子(variance inflation factor,VIF)检验气候变量间的共线性,去掉VIF值大于4的气候变量(Kabacoff, 2013)。
1.4 数据统计与分析
以古田山样地群落数据为基础,计算各样地的物种重要值(importance value, IV),衡量物种在群落中的数量特征,作为划分群系类型的依据(宋永昌,2016)。计算公式:
IV=(Dr+Pr+Fr)/3。
式中:Dr为相对多度;Pr为相对显著度;Fr为相对频度。
采用Srensen相异性系数(Srensen dissimilarity index, QS)度量常绿阔叶林群落间的物种β多样性。计算公式:
QS=1-(2|X∩Y|)/(|X|+|Y|)。
式中:X,Y为两个群落的物种,不考虑种多度。
采用非加权组平均法(unweighted pair group method with arithmetic mean, UPGMA)对群落组成进行聚类分析。
利用经度、纬度计算不同地区常绿阔叶林群落间的地理距离矩阵。将各环境变量标准化,对各环境变量计算地区间的欧氏距离。
根据赤池信息量准则(akaike information criterion, AIC)选择对物种β多样性有显著影响的气候变量,将所选气候变量和海拔统一计算为环境差异矩阵。由于各样地面积差异较大,为了控制样地面积差异造成的影响,将样地间面积差异作为一个解释变量进行分析。
利用基于距离矩阵的多元回归(multiple regressions on distance matrices, MRM)进行偏回归分析(Lichstein, 2007),在控制样地面积差异的基础上,了解环境距离、地理距离与物种β多样性指数之间的关系。计算各常绿阔叶林群落中常绿阔叶与落叶阔叶物种数量的比值(即常绿阔叶物种数量/落叶阔叶物种数量),作为描述群落叶生活型组成特点的指标。选出AIC值最小的模型作为最优模型,采用线性回归模型了解环境因素对常绿/落叶值的影响。
所有数据分析均通过R 3.2.4軟件进行,其中Srensen相异性系数、聚类分析、模型选择、MRM分析等分别采用Vegan、Stats、MuMIn、Ecodist等软件包进行计算。
2 结果与分析
2.1 物种组成和群系类型
15个常绿阔叶林共记录到木本植物78科233属617种。含10属以上的科有4个,分别为蔷薇科(19属)、豆科(11属)、樟科(10属)和茜草科(10属)。含20种以上的科有5个,分别为蔷薇科(59种)、樟科(54种)、壳斗科(51种)、山茶科(36种)和冬青科(30种);单属科有虎皮楠科、八角枫科、山矾科等32个科;单种科有银杏科、领春木科、伯乐树科等19个科。大戟科、冬青科、壳斗科、山茶科和樟科5个科在15个常绿阔叶林均有分布。五列木科、七叶树科、银杏科等13个科只在一个地区有记录。
不同地区优势物种差异明显(表1),基本涵盖了我国东部典型常绿阔叶林的主要群系类型。
根据重要值将古田山常绿阔叶林样地划分为栲类林(17个)、木荷林(9个)和青冈林(3个)(表3),其中栲类林和木荷林是古田山典型的常绿阔叶林类型,青冈类林为古田山常见的次生林类型(于明坚等,2001)。
2.2 物种β多样性及影响因素
古田山与其他群落的平均Srensen相异性系数最低(0.85),瓦屋山与其他群落的平均相异性系数最高(0.96)。与其他14个地区的常绿阔叶林相比,古田山常绿阔叶林在物种组成上与黄山和天童山的差异最小,而与峨眉山和瓦屋山的差异最大(表4,图1)。
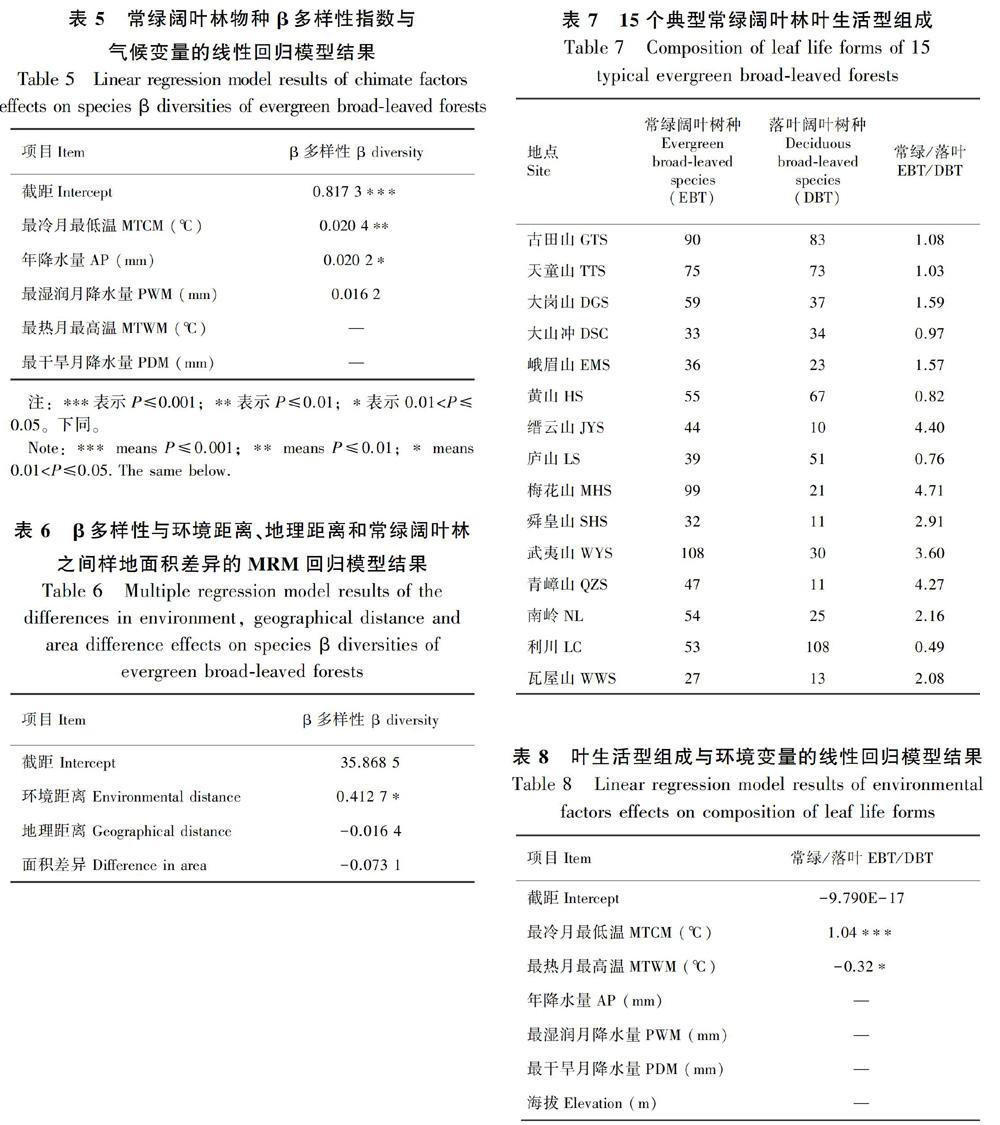
8个气候变量中,去除年均温(AMT)、全年温度变化(TAR)和温度季节性(TS)三个存在多重共线性的变量(VIF>4)。经过模型选择,气候因素中年降水量(AP)、最冷月最低温(MTCM)和最湿润月降水量(PWM)对东部典型常绿阔叶林物种β多样性有明显影响。其中,最冷月最低温(MTCM)对物种β多样性影响最大且显著(P<0.001)(表5)。进一步,环境距离对不同地点间常绿阔叶林群落物种组成差异有显著影响(P<0.05),而地理距离和样地面积影响均不显著(表6)。
2.3 叶生活型组成及影响因素
常绿阔叶种较落叶阔叶种丰富度优势(即常绿阔叶物种数/落叶阔叶物种数)最高的是梅花山(4.71),最低的是利川(0.49)(表7)。影响常绿种优势的主要气候因子为最热月最高温(MTWM)和最冷月最低温(MTCM),其中最冷月最低温(MTCM)的影响最为显著(P<0.01)(表8)。
3 讨论
3.1 不同地区常绿阔叶林物种组成比较
本研究中,有12个地区的常绿阔叶林以栲类林和木荷林为主,锥属和木荷属物种占优势;大山冲和庐山两地的常绿阔叶林以青冈林、石栎林为主,石栎(Lithocarpus glaber)、青冈(Cyclobalanopsis glauca)等常绿阔叶植物占明显优势;利川楠木群落是楠木(Phoebe zhennan)形成的单优群落。
栲类、木荷林分布广泛,是我国东部典型常绿阔叶林分布面积最大、最具代表性的类型(宋永昌,2013)。古田山以甜槠—木荷群落为主体,同时分布有青冈林, 甜槠—木荷群落主要分布在我国亚热带偏南的中低海拔地区,是浙闽赣皖交界山地低海拔常绿阔叶林的主体类型;青冈林是我国东部典型的次生常绿阔叶林,多分布在亚热带中北部低海拔较湿润地区,并作为原生常绿阔叶林在低海拔沟谷地段和中山地带的沟谷和坡地存在(于明坚等,2001)。宋永昌(2013)研究结果表明,梅花山和武夷山常绿阔叶林以少叶黄杞(Engelhardtia fenzlii)、米槠(Castanopsis carlesii)等物种占优势,是亚热带中部中低海拔地区常见的类型;南岭常绿阔叶林分布有雷公青冈(Cyclobalanopsis hui)、猴欢喜(Sloanea sinensis)等华南地区常见种,青嶂山分布有红锥(Castanopsis hystrix)、赤皮青冈(Cyclobalanopsis gilva)等喜温暖湿润气候的树种,代表亚热带偏南的典型常绿阔叶林类型。缙云山以银木荷(Schima argentea)、短刺米槠(Castanopsis carlesii var. spinulosa)为优势种,还分布有四川大头茶(Polyspora speciosa)群落,峨眉山和瓦屋山均以扁刺锥 (Castanopsis platyacantha) 占优势, 三个地点均有东部湿润典型常绿阔叶林向西部半湿润典型常绿阔叶林过渡的特点,这与其他常绿阔叶林群落差异较大。
古田山、天童山和黃山物种组成最为相似,但也有明显差异。天童山的常绿阔叶林主要为木荷林(吴洋洋等,2014),未发现常绿阔叶林常见的建群种甜槠(Castanopsis eyrei);黄山常绿阔林以甜槠林为主,与古田山的优势物种更为相似;古田山和天童山常绿阔叶林中常绿植物物种丰富度均占优势,而黄山温带性质更为明显,表现为落叶物种丰富度高于常绿物种(丁晖等,2016)。因此认为古田山甜槠—木荷群落的物种组成比天童山和黄山更加典型。古田山与其他常绿阔叶林的平均Srensen相异性指数最小,提示古田山的常绿阔叶林的物种组成在我国东部低海拔常绿阔叶林中具有一定代表性;瓦屋山与其他常绿阔叶林的平均Srensen相异性指数最大,可能由于瓦屋山物种组成向西部典型常绿阔叶林过渡的特点明显,与典型的东部典型常绿阔叶林物种组成差异较大。
3.2 影响我国东部典型常绿阔叶林物种组成的主要因素
生态位过程与中性过程对群落构建的相对重要性因研究地区而异,同时受到研究尺度、物种生活型等因素的影响,如温带森林的木本植物更多受到生态位过程的影响,热带森林则主要被中性过程影响(Myers et al., 2013);赵鸣飞等(2017)发现我国温带与亚热带交界处的森林群落内物种组成主要被地理距离解释,但乔木层物种组成受到生境过滤的影响较大。本研究针对我国东部典型常绿阔叶林,发现环境差异比地理距离对木本植物物种组成相似性的影响更为显著,与其他在我国亚热带地区较大尺度的研究结果一致(Liu et al., 2016),表明以生境过滤为代表的生态位过程是影响我国东部典型常绿阔叶林木本植物物种分布的主要因素。气候因素中,最冷月最低温(MTCM)对物种组成影响最显著,验证了大尺度物种分布地理格局的寒冷忍耐假说。
最冷月最低温(MTCM)是我国东部典型常绿阔叶林中常绿阔叶植物物种丰富度优势程度的主要影响因素,降水量则无明显影响。同样,最冷月最低温是影响我国木本植物物种丰富度的最主要因素(Wang et al., 2011);黄永涛等(2015)发现鄂西南亚热带常绿阔叶混交林中常绿和落叶物种丰富度的变化主要受到温度的限制,而非水分;Zhang et al.(2010)发现影响我国常绿阔叶林叶生活型相对重要值的主要气候因素为年均温和年降水量;针对我国亚热带常绿落叶阔叶混交林,常绿阔叶物种和落叶阔叶物种相对重要值主要受到最冷月最低温的影响,同时受到降水量的明显影响(Ge & Xie, 2017)。这表明在不同类型、不同研究尺度的森林群落中对叶生活型组成起主导作用的气候因素不同;这也表明常绿阔叶和落叶阔叶物种丰富度和重要值差异受到不同因素的驱动,常绿阔叶和落叶阔叶植物之间重要值的差异受到温度和降水量的共同影响,而物种丰富度主要受到温度的影响。
本研究探究了东部典型常绿阔叶林的物种组成差异及影响因素,为了解该种植被亚型对气候变化的响应提供一定科学依据。但由于本研究收集到的群落样地资料并没有全面覆盖我国东部典型常绿阔叶林,且不同样地的调查时间和方法存在差异,环境因素主要关注气候因子,缺乏土壤、演替阶段等资料,因此有一定的局限性,更全面深入的研究还有待于进一步完善数据。
致谢 浙江大学王月霞、骆杨青等同学参与古田山野外调查、数据录入等工作;赖正标、赖祯熙等师傅在野外调查工作中付出辛勤劳动,在此致谢。同时,向文中引用的常绿阔叶林样地的研究者致谢!
参考文献:
BAI KD, JIANG DB, CAO KF, et al., 2010. Photosynthetic response to seasonal temperature changes in evergreen and deciduous broad-leaved trees in montane forests of Ailao Mountain and Maoer Mountain [J]. Acta Ecol Sin, 30(4):905-913. [白坤栋, 蒋得斌, 曹坤芳, 等, 2010. 哀牢山和猫儿山中山常绿和落叶阔叶树光合特性对季节温度变化的响应 [J]. 生态学报, 30(4):905-913.]
CHASE JM, 2014. Spatial scale resolves the niche versus neutral theory debate [J]. J Veg Sci, 25(2):319-322.
DING H, FANG YM, YANG XH, et al., 2016. Community characteristics of a subtropical evergreen broad-leaved forest in Huangshan, Anhui Province, East China [J]. Biodivers Sci, 24(8):875-887. [丁晖, 方炎明, 杨新虎, 等, 2016. 黄山亚热带常绿阔叶林的群落特征 [J]. 生物多样性, 24(8) :875-887.]
GARZON-LOPEZ C X, JANSEN PA, BOHLMAN SA, et al., 2014. Effects of sampling scale on patterns of habitat association in tropical trees [J]. J Veg Sci, 25(2):349-362.
GE J, XIE Z, 2017. Geographical and climatic gradients of evergreen versus deciduous broad-leaved tree species in subtropical China:Implications for the definition of the mixed forest [J]. Ecol Evol, 7(11):3636-3644.
HIJMANS RJ, CAMERON SE, PARRA JL, et al., 2005. Very high resolution interpolated climate surfaces for global land areas [J]. Int J Climatol, 25(15):1965-1978.
HUANG YT, YAO L, AI XR, et al., 2015. Quantitative classification of the subtropical evergreen-deciduous broad-leaved mixed forest and the deciduous and evergreen species composition structure across two national nature reserves in the Southwest of Hubei, China [J]. Chin J Plant Ecol, 39(10): 990-1002. [黄永涛, 姚兰, 艾训儒, 等, 2015. 鄂西南两个自然保护区亚热带常绿落叶阔叶混交林类型及其常绿和落叶物种组成结构分析 [J]. 植物生态学报, 39(10):990-1002.
HU ZH, YU MJ, DING BY, et al., 2003. Types of evergreen broad-leaved forests and their species diversity in Gutian Mountain National Nature Reserve [J]. Chin J Appl Environ Biol, 9(4):341-345. [胡正华, 于明坚, 丁炳扬, 等, 2003. 古田山国家级自然保护区常绿阔叶林类型及其群落物种多样性研究 [J]. 应用与环境生物学报, 9(4):341-345.]
JIN Y, RUSSO SE, YU M, 2018. Effects of light and topography on regeneration and coexistence of evergreen and deciduous tree species in a Chinese subtropical forest [J]. J Ecol, 106:1634-1645.
KABACOFF RI, 2013. R in action:Data analysis and graphics with R [M]. GAO T, XIAO N, CHEN G (translator). Beijing: The Peoples Posts and Telecommunications Press:158-197. [KABACOFF RI, 2013. R语言实战 [M]. 高涛,肖楠,陈钢(译). 北京:人民邮电出版社:158-197.]
LICHSTEIN JW, 2007. Multiple regression on distance matrices:A multivariate spatial analysis tool [J]. Plant Ecol, 188(2):117-131.
LIU M, 2010. Community analysis of low hill evergreen broad-leaved forest in Nanling National Nature Reserve, Guangdong Province [J]. Guangdong For Sci Technol, 26(3): 65-70. [劉敏, 2010. 南岭国家级自然保护区低山常绿阔叶林植物群落分析 [J]. 广东林业科技, 26(3):65-70.]
LOU LH, JIN SH, 2000. Spermatophyta flora of Gutianshan Nature Reserve in Zhejiang [J]. J Beijing For Univ, 22(5):33. [楼炉焕, 金水虎, 2000. 浙江古田山自然保护区种子植物区系分析 [J]. 北京林业大学学报, 22(5):33.]
LIU Jl, QIAN H, JIN Y, et al., 2016. Disentangling the drivers of taxonomic and phylogenetic beta diversities in disturbed and undisturbed subtropical forests [J]. Sci Rep, 6:35926.
LIU Y, TANG Z, FANG J, 2015. Contribution of environmental filtering and dispersal limitation to species turnover of temperate deciduous broad-leaved forests in China [J]. Appl Veg Sci, 18(1):34-42.
MYERS JA, CHASE JM, JIMENEZ I, et al., 2013. Beta-diversity in temperate and tropical forests reflects dissimilar mechanisms of community assembly [J]. Ecol Lett, 16(2):151-157.
NIU KC, LIU YN, SHEN ZH, et al., 2009. Community assembly:the relative importance of neutral theory and niche theory [J]. Biodivers Sci, 17(6):579-593. [牛克昌, 刘怿宁, 沈泽昊, 等, 2009. 群落构建的中性理论和生态位理论 [J]. 生物多样性, 17(6):579-593.]
SONG YC, 2013. Evergreen broad-leaved forests in China [M]. Beijing:Science Press:3-177. [宋永昌, 2013. 中国常绿阔叶林 [M]. 北京:科学出版社:3-177.]
SONG YC, 2016. Vegetation ecology [M]. 2nd ed. Beijing:Higher Education Press:39-356. [宋永昌, 2016. 植被生态学 [M]. 2版. 北京:高等教育出版社:39-356.]
SONG YC, YAN ER, SONG K, 2015. Synthetic comparison of eight dynamics plots in evergreen broad-leaf forests,China [J]. Biodivers Sci, 23(2):139-148. [宋永昌, 阎恩荣, 宋坤, 2015. 中国常绿阔叶林8大动态监测样地植被的综合比较 [J]. 生物多样性, 23(2):139-148.]
SOININEN J, MCDONALD R, HILLEBRAND H, 2007. The distance decay of similarity in ecological communities [J]. Ecography, 30(1):3-12.
TAO WL, HU G, ZHANG ZH, et al., 2018. Latitudinal patterns in geographical elements of woody plants in a karst area of southwestern China [J]. Plant Sci J, 36(5):667-675. [陶旺兰, 胡刚, 张忠华, 等, 2018. 西南喀斯特木本植物区系成分的纬度变异格局 [J]. 植物科学学报, 36(5):667-675.]
WANG XH, 2006. The phytogeography and species diversity of typical evergreen broad-leaved forest in China [D]. Shanghai:East China Normal University:1-111. [王希华, 2006. 中国典型常绿阔叶林植物地理与物种多样性研究 [D]. 上海:华东师范大学:1-111.]
WANG ZH, FANG JY, TANG ZY, LIN X, et al., 2011. Patterns, determinants and models of woody plant diversity in China [J]. Proc Biol Sci, 278(1715):2122-2132.
WU YY, GUO CZ, NI J, 2014. Dynamics of major forest vegetations in Tiantong National Forest Park during the last 30 years [J]. Chin J Appl Ecol, 25(6):1547-1554. [吴洋洋, 郭纯子, 倪健, 2014. 天童国家森林公园主要森林植被过去30年的动态变化 [J]. 应用生态学报, 25(6):1547-1554.]
WU ZY, 1980. Vegetation of China [M]. Beijing:Science Press:306-356. [吴征镒, 1980. 中国植被 [M]. 北京:科学出版社:306-356.]
YU MJ, HU ZH, YU JP, et al., 2001. Forest vegetation types in Gutianshan Nature Reserve in Zhejiang [J]. J Zhejiang Univ (Agric Life Sci Ed), 27(4):375-380. [于明堅, 胡正华, 余建平, 等, 2001. 浙江古田山自然保护区森林植被类型 [J]. 浙江大学学报(农业与生命科学版), 27(4):375-380.]
ZHANG L, LUO TX, ZHU HZ, et al., 2010. Leaf life span as a simple predictor of evergreen forest zonation in China [J]. J Biogeogr, 37:27-36.
ZHAO MF, WANG GY, XING KX, et al., 2017. Patterns and determinants of species similarity decay of forest communities in the western Qinling Mountains [J]. Biodivers Sci, 25(1): 3-10. [赵鸣飞, 王国义, 邢开雄, 等, 2017. 秦岭西部森林群落相似性递减格局及其影响因素 [J]. 生物多样性, 25(1):3-10.]
(责任编辑 蒋巧媛)
猜你喜欢
小学教学参考(语文)(2016年12期)2017-01-04
未来英才(2016年20期)2017-01-03
理论导刊(2016年12期)2016-12-27
甘肃教育(2016年22期)2016-12-20
新教育时代·教师版(2016年26期)2016-12-06
求知导刊(2016年30期)2016-12-03
戏剧之家(2016年22期)2016-11-30
散文百家·下旬刊(2016年9期)2016-11-23
人间(2016年27期)2016-11-11
农业与技术(2016年15期)2016-11-09
