
猕猴桃软腐病主要病原菌的分离、鉴定及其生长特性研究
2020-12-04 06:53王仁才王芳芳卜范文
核农学报 2020年11期
石 浩 王仁才 王芳芳 王 琰 卜范文 周 倩
(1 湖南农业大学园艺园林学院, 湖南 长沙 410128;2 湖南应用技术学院农林科技学院, 湖南 常德 415100;3 湖南省园艺研究所,湖南 长沙 410125)
猕猴桃(kiwifruit)是一种在国内外都十分受欢迎的经济作物,富含维生素C、可溶性膳食纤维、原花青素、黄酮等功能物质[1-2],具有较高的食用价值。近几年来,随着其栽种规模的逐年增加,各种病害也接踵而至[3]。其中对猕猴桃采后危害最严重的是猕猴桃软腐病,在我国陕西、贵州、四川、湖南、江西等地病害现象严重,发病率高达35%以上,对猕猴桃商品果的质量造成了具大的影响[4],对农户也造成了一定的经济损失。1982年新西兰人Hawthorne等[5]首次对猕猴桃软腐病进行了报道,随后意大利、韩国和中国也陆续进行了相关报道[6-9]。猕猴桃软腐病病状主要表现为果实病灶部出现1~3 cm直径大小、灰暗色的凹陷病斑,病斑处果肉腐化较为严重,果肉呈现乳白色。果实发病后期,在较高温湿度环境下,果实病部处出现菌丝,同时表皮渗出一定的组织液。研究表明,无任何损伤的猕猴桃果实也能发病,且各部位均有病害出现的情况。损伤果发病情况更严重,发病时间提前,在无损伤果实软化前期病害较难发现,而一旦出现病害后,7~9 d内可导致整个果实完全腐烂[10]。因此通过分子技术手段,在果实未发病时及时发现病原菌,并进行相应病害防治十分必要。目前有关猕猴桃果实软腐病病原菌鉴定的研究报道不多,同时国内外学者对引起软腐害病原菌的报道存在一定的争议,且不同品种间病原菌状况也有所差异[11-12],为此有必要对湖南地区猕猴桃软腐病害进行进一步的研究。目前常采用对病菌基因间隔序列(internal transcribed spacer, ITS)、β-微管蛋白基因(β-tubulin)和延伸因子(EF-1α)测序结果构建发育树的方法来判定病原菌种类[13-14]。本试验通过对软腐病原菌分离,ITS测序鉴定病原菌并对病原菌生长特性研究,以期进一步了解猕猴桃软腐病的主要病菌,间接确定猕猴桃安全贮藏和影响病害发生的关键因子,为研发猕猴桃采后病害防控及保鲜技术提供参考依据。
1 材料与方法
1.1 材料
猕猴桃(品种:红阳)软腐病病菌感染果和好果,于2017年采自湖南省湘西州猕猴桃园,将果实装入保鲜袋中,保藏于4℃冰箱备用。
1.2 主要仪器与设备
CP223C电子分析天平,奥豪斯仪器(上海)有限公司;SX-500高压蒸汽灭菌锅,海珂淮仪器有限公司;DSZ2000X倒置生物显微镜,北京中显恒业仪器仪表有限公司;GZ-400-GII恒温光照培养箱,韶关市广智科技设备有限公司;PHS-25型PH计,上海仪电控股(集团)公司;T100TM Thermal Cycler PCR仪,美国Bio-Rad公司。
1.3 试验方法
1.3.1 猕猴桃软腐病病原菌的分离与纯化 取刚产生软腐病病状的猕猴桃果实,用无菌水冲洗杂质后除去果实表面水分,然后用已高温灭菌的解剖刀刮取猕猴桃发病果实病健交界处的组织,用75%乙醇喷洗消毒约10 s,再用大量无菌水洗去残余乙醇,用已灭菌的滤纸吸干无菌水,挑取少许待分离的材料组织采用点接法接种于PDA平板上,在27℃培养箱中避光培养3~5 d后,可在平板表面观察到系列猕猴桃软腐病病原菌的混合菌落[15]。
从混合菌落中挑取较规则的单一菌落接种在PDA平板上,在27℃培养箱中避光培养3~5 d,重复纯化3次,得单菌落。单菌落培养7~10 d后,观察菌丝和孢子形态。同时取出菌落孢子,用无菌水冲洗后稀释孢子,在PDA平板上将孢子均匀涂抹培养,待长成菌落后挑取部分菌落培养在PDA斜面上,4℃冰箱保存[15-16]。
1.3.2 猕猴桃软腐病病原菌形态学鉴定与致病性验证 将已经分离好的单株病原菌,重新接种在新鲜培养基上,27℃条件下培养一个月,期间每隔1~2 d观察一次菌丝的生长情况,包括菌落的形态、长度,菌丝的密集度、颜色和形状等。培养一个月后,观察菌产生孢子的数量、结构、形态等,结合文献[17-18]初步判断病原菌的种类。
取10个完好的猕猴桃果实(未染病),其中4个果实接种分离得到的软腐病病原菌菌丝,4个果实接种软腐病病原菌孢子悬浮液,2个果实接种无菌水(对照)。采用针刺接种法接种,培养10 d后观察果实发病情况,并对症状与原发病果进行比较。同时对接种后猕猴桃病果重新进行分离、纯化及鉴定病原菌,确定是否为同一种病菌[19]。
1.3.3 猕猴桃软腐病病原菌的分子鉴定 培养7~10 d后,应用CTAB法[20]提取纯化后病原菌的DNA,以真菌通用ITS4、ITS5为引物。总PCR反应体系为24 μL,包括2 μL含有20~30 ng DNA模板、1 μL 10 μmol·L-1引物、12.5 μL Premix Ex Taq@Version 2.0及8.5 μL双蒸水。PCR扩增程序:94℃预变性30 s;94℃变性30 s,55℃退火30 s,72℃延伸30 s,共32个循环;72℃延伸7 min[15]。回收PCR产物送至生工生物工程(上海)股份有限公司测序,再根据所得序列,采用MEGA7软件构建系统发育树。
1.3.4 猕猴桃软腐病病原菌生物学生长特性分析 参考丁仁惠等[21]的方法并稍作修改。将直径为5 mm左右的菌饼分别接种于下列不同培养基、碳源、氮源、pH值、培养温度的环境中培养,3 d后采用十字交叉法测量菌落直径并记录菌丝密集度,设3次重复。无菌水为对照(CK),除pH值处理外,其余均为自然pH值,除温度处理试验外,其余处理均27℃培养。菌丝生长抑制率计算公式如下:
菌抑制率=(菌丝长对照-菌丝长处理)/菌丝长处理×100%。
培养基:土豆葡萄糖培养基(potato dextrose agar, PDA)、马铃薯蔗糖培养基(potato saccharose agar, PSA)、水-琼脂培养基(water agar, WA)、沙氏培养基(sabouraud agar, SDA)、察氏培养基(czapek agar, CA)、高氏1号培养基(Gauze,s Medium No.1)、燕麦培养基(oat meal agar, OMA)、玉米粉培养基(corn meal medium, CMM)、牛肉膏蛋白胨培养基(luria bertani, LB)、酵母蛋白胨葡萄糖培养基(yeast extract peptone glucose agar, YPGA)。
碳源:甘露醇、麦芽糖、蔗糖、乳糖、D-山梨醇、可溶性淀粉、木糖、肌醇、D-半乳糖9种碳源代替PDA培养基中的葡萄糖。
氮源:氯化铵、酵母粉、甘氨酸、尿素、硝酸钾、蛋白胨、硝酸铵、L-丙氨酸、L-赖氨酸9种氮源代替CA培养基中的硝酸钠。
pH值分别为4、5、6、7、8、9、10、11、12、13的PDA培养基。
培养温度分别为4、5、10、15、20、25、28、30、35℃。
菌饼置离心管中,离心管分别置于60~90℃(温度梯度为5℃)的恒温水浴锅中处理30 min,冷却后接种至PDA培养基上,确定病原菌致死温度。
1.3.5 数据处理 采用WPS 2019对试验数据进行处理和分析。采用Statistix 8.0软件对试验数据进行统计分析。
2 结果与分析
2.1 猕猴桃软腐病病原菌培养形态学观察
从软腐猕猴桃果实中分离出两种主要病原菌,分别接种于PDA培养基上,图1-a-1~4为其中一种菌(占总菌落数54.8%),图1-b-1~4为另一种菌(占总菌落数34.4%)。图1-a-1和图1-b-1均为培养5~7 d的菌丝,其中图1-a-1中菌丝为纯白色的密集卷毛状,病原菌的菌丝较稀疏,菌落圆形,绒毛状,无特殊气味;图1-b-1中菌丝非常稠密,长圆形,灰色、近棒状,无特殊气味。图1-a-2和图1-b-2分别为两种菌培养30 d后生长形态,其中,图1-a-2中产生黑色的零散子座,有孔口,上部有不平整凸起;图1-b-2中菌丝干枯,呈现灰黑色。显微镜观察发现,图1-a-3菌丝有隔膜和分枝,白色;图1-a-4分生孢子有两种形态,α形分生孢子透明椭球,β分生孢子透明,线状和一端有钩状结构,分生孢子以α形为主初步鉴定为间座壳菌(Diaporthephaseolorum),记作NJDMJ[15];图1-b-4菌丝同样有隔膜和分枝,黑色,菌丝直径为1.0~4.9 μm,可初步鉴定为葡萄座腔菌(Botryosphaeriadothidea),记作PTZQJ[18]。

注:a、b分别表示同一种病原菌,其中,1为培养5~7 d的病原菌菌丝; 2为培养30 d时的病原菌形态,3和4均为显微观察到的病原菌形态。Note: a and b show the same pathogenic bacteria. Among them, 1 is the mycelium of pathogenic bacteria cultured for 5 to 7 days, 2 is the mycelium of pathogenic bacteria cultured for 30 days, 3 and 4 are microscopic observation of pathogenic bacteria morphology.图1 软腐病病状的猕猴桃果实中分离的病原菌形态Fig.1 Morphology of pathogens isolated from kiwifruit fruits with soft rot disease
2.2 猕猴桃软腐病病菌感染果及接种后猕猴桃好果的发病症状
图2-a、b为猕猴桃软腐病病菌感染果的发病症状,有明显的暗褐色病斑圈。图2-c~e分别为接种白色菌丝病原菌,黑色菌丝病原菌,以及白色菌丝病原菌、黑色菌丝病原菌混合孢子后的猕猴桃发病症状。可知,在室温27℃和湿度80%左右环境下培养7~10 d,接种菌丝(图2-c、d)或菌液(图2-e)侵染均能使果实发病。接种病原菌后,猕猴桃果实最显著特征是果实表面有明显的凹陷和似圆形的水渍状病斑,中央病斑呈暗褐色圆形,拇指按压后尤为明显,触摸时有类似气囊触感,剥除果皮后果实内部可见乳白色圆形或椭圆形病斑,同时有组织液渗出,后期还长出肉眼可见的菌丝,隐约有腐臭气味。进一步对接种后发病猕猴桃中的病原菌进行分离纯化,得到的病原菌生长形态、菌丝、孢子均与接种的病原菌一致,可初步鉴定为同一病原菌。
2.3 猕猴桃软腐病病原菌分子鉴定
将分离得到的两种病原菌株所得序列登录NCBI进行BLAST相似度比对,并将病原菌株的序列与同源性为99%的相关序列在MEGA7软件上进行系统发育树的构建。由图3可知,黑色菌株PTZQJ与Botryosphaeriasp(葡萄座腔菌)聚在同一枝上,同源性达到99%,这与形态学鉴定结果一致。白色菌株NJDMJ与Phomopsissp(拟茎点霉菌)、Diaporthephaseolorum(间座壳菌,拟茎点霉菌有性态)聚在同一枝上,同源性达到99%,该菌株为大豆拟茎点种腐病菌,这与上述形态学鉴定结果也一致。
2.4 猕猴桃病原菌生态学特性
2.4.1 不同培养基对猕猴桃病原菌生长菌丝的影响 由表1可知,在相同培养基下葡萄座腔菌的生长速度总体较间座壳菌快,且菌丝更稠密。如在PSA上葡萄座腔菌的生长速度为17.7 mm·d-1,较间座壳菌的生长速度高2.4 mm·d-1;在YPGA上葡萄座腔菌的生长速度为16.4 mm·d-1,较间座壳菌的生长速度高2.5 mm·d-1。在PSA、YPGA、PDA、SDA上两种病原菌的生长速度均较快,且菌丝比较稠密,而在LB和WA两种无糖培养基上,病原菌均无法生长,说明糖是这两种菌生长的必要条件。此外,两种病原菌都能在CMM、OMA、高氏1号、CA上生长,但生长效果较差,生长速率在6.1~10.2 mm·d-1之间。

图3 基于ITS序列构建的猕猴桃病原菌系统发育树Fig.3 The phylogenetic tree of kiwifruit pathogens based on ITS sequences
2.4.2 不同碳源对猕猴桃病原菌菌丝生长的影响 碳源是微生物生长中不可或缺的营养物,是构成微生物细胞的骨架,同时也在微生物生长发育中提供能量。由表2可知,猕猴桃两种病原菌对糖的依耐性都很强,其中,在未添加碳源的培养基(CK)上两种病原菌的生长速率仅为2.3、2.7 mm·d-1,几乎不生长,而在分别以可溶性淀粉、蔗糖和麦芽糖为碳源的培养基上两种病原菌生长的效果较好,菌丝生长速率在12.4~17.7 mm·d-1之间,在以D-半乳糖、乳糖、D-山梨糖为碳源的培养基上两种病原菌菌丝生长速率在9.2~11.9 mm·d-1之间。不同类型的碳源对两种菌的生长具有较大的差异,可能是因为病原菌对不同碳源的吸收利用不同,如以乳糖为碳源的培养基上间座壳菌的生长速度高于葡萄座腔菌,而以麦芽糖为碳源的培养基上间座壳菌的生长速度低于葡萄座腔菌。
2.4.3 不同氮源对猕猴桃病原菌菌丝生长的影响 由表3可知,以酵母粉、蛋白胨、甘氨酸、L-丙氨酸和L-赖氨酸为氮源的培养基均较适宜猕猴桃两种病原菌的生长,葡萄座腔菌生长速率在10.9~11.2 mm·d-1之间,间座壳菌生长速率在9.0~9.7 mm·d-1之间;以氯化铵、硝酸钾、硝酸铵、尿素为氮源的培养上,葡萄座腔菌生长速率在9.6~10.3 mm·d-1之间,间座壳菌生长速率在8.6~9.2 mm·d-1之间。此外,未添加氮源的培养基(CK)上猕猴桃两种病原菌的生长速率也较高,在8.8~10.2 mm·d-1之间,可能是因为琼脂中含有一定量的氮源,但也间接说明两种病原菌对氮源的吸收利用量较低。
2.4.4 不同pH值对猕猴桃病原菌生长的影响 真菌对环境pH值要求较高,一旦环境中的pH值出现异常,会在一定程度改变营养物质的供给状态,给菌体细胞膜电荷性质和稳定性带来负面影响,继而影响菌体对营养物质的吸收能力。由表4可知,猕猴桃两种病原菌菌丝在pH值4.0~13.0的培养基上均可生长,其中,pH值为6.0~9.0时,两种病原菌菌丝生长速度均较快,葡萄座腔菌菌落生长速率达到了18.1~19.3 mm·d-1, 菌丝也非常浓密,间座壳菌菌落生长速率达到16.2~17.2 mm·d-1。而当pH值达到12时,与pH值6时相比,菌丝生长速率相差5 mm·d-1以上。综上,猕猴桃病原菌菌丝适合在pH值6左右的中酸性环境中生长,而在强酸强碱环境中生长缓慢。
2.4.5 不同培养温度对猕猴桃病原菌菌丝生长的影响 由表5可知,猕猴桃两种病原菌在低温(4~5℃)条件下几乎不生长,而培养温度高于10℃时,病原菌菌丝生长速率开始缓慢增加,在培养温度为25~30℃条件下两种病原菌生长最快,其中,培养温度为28℃时葡萄座腔菌生长速率最快,达到18.8 mm·d-1,培养温度为25℃时间座壳菌生长速率最快,达到17.4 mm·d-1。 继续升高培养温度至35℃时,两种病原菌的生长速率均有一定程度的下降,表明培养温度达到35℃后开始抑制猕猴桃病原菌的生长。

表1 不同培养基对猕猴桃病原菌菌丝生长的影响Table 1 The effects of different mediums on mycelial growth of kiwifruit pathogens
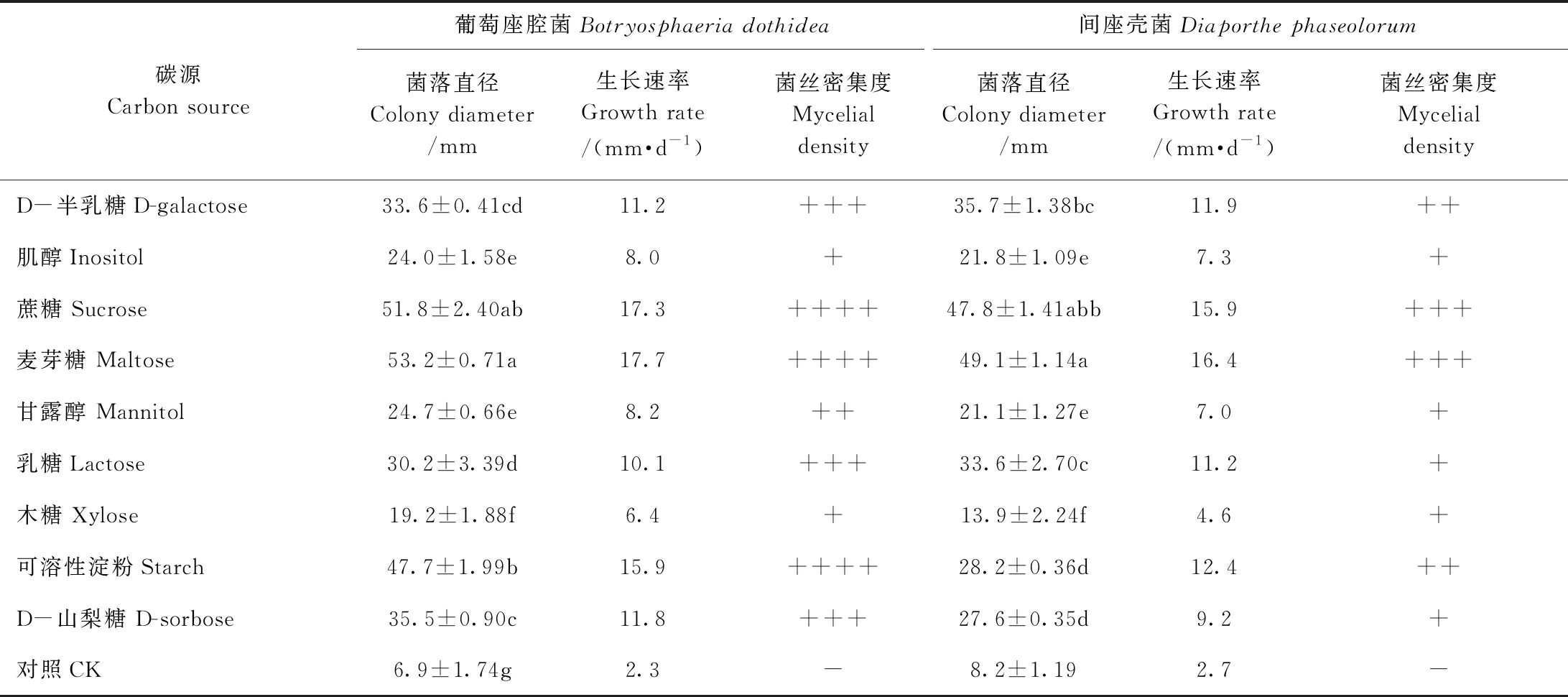
表2 不同碳源对猕猴桃病原菌菌丝生长的影响Table 2 The effects of different carbon sources on mycelial growth of kiwifruit pathogens
2.4.6 猕猴桃病原菌致死温度分析 由表6可知,在80℃水浴处理30 min后,接种于PDA培养基上的猕猴桃两种病原菌均得到了较大程度的抑制,相对于60℃水浴处理30 min,葡萄座腔菌生长抑制率达到了60.29%,间座壳菌生长抑制率达到了82.31%;85℃水浴处理30 min后,间座壳菌死亡,葡萄座腔菌几乎死亡;90℃水浴处理30 min后,两种病原菌均无法生长。因此,确定猕猴桃采后软腐病原菌的致死温度为85℃。

表3 不同氮源对猕猴桃病原菌菌丝生长的影响Table 3 The effects of different nitrogen sources on mycelial growth of kiwifruit pathogens
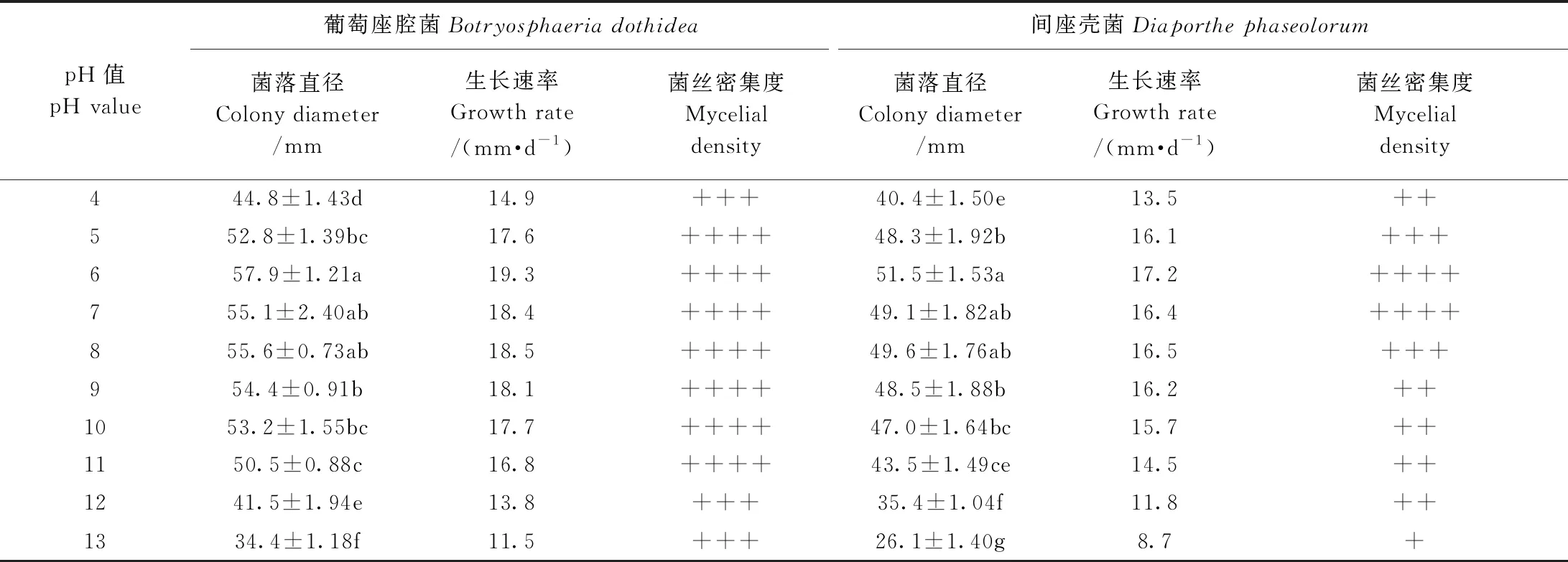
表4 不同pH值对猕猴桃病原菌菌丝生长的影响Table 4 The effects of different pH value on mycelial growth of kiwifruit pathogens

表5 不同培养温度对猕猴桃病原菌菌丝生长的影响Table 5 The effects of different temperatures on mycelial growth of kiwifruit pathogens
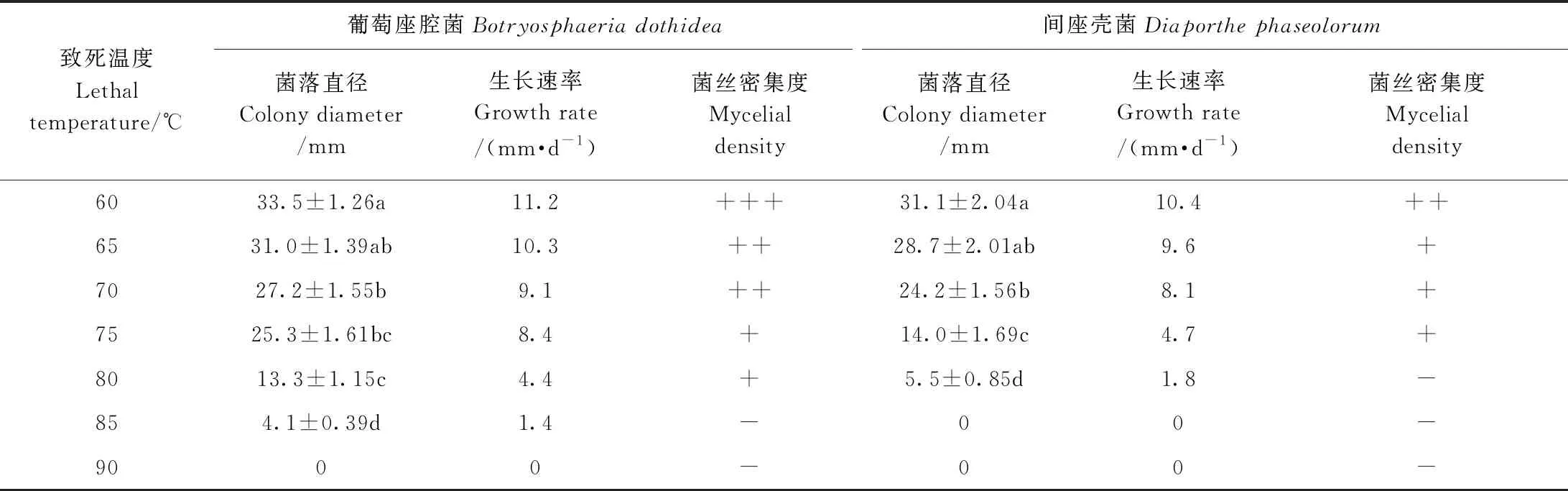
表6 猕猴桃病原菌的致死温度Table 6 The lethal temperature of pathogenic bacteria
3 讨论
病原菌鉴定是病害防治的关键。通过分离猕猴桃采后软腐病果上的菌体达到对病原菌初步形态学鉴定的目的,观察菌丝、孢子形态特征,孢子囊的产生方式,孢子释放方式及分子鉴定,是确定菌类所属的主要方式[22-23]。本试验采用上述方法确定猕猴桃软腐主要病害为葡萄座腔菌和间座壳菌。本试验结果与前期软腐病害果微生物多样性的结果基本一致,进一步验证了这两种病菌是造成猕猴软腐的主要病菌。本研究还发现,将两种病原菌回接于猕猴桃果实上,均能引起果实发生类似的病斑,且在后期能产生丰富的分生孢子,这与前人相关研究结果基本一致[11,17]。相对于陆训等[15]从软腐猕猴桃中发现的病原菌,大豆拟茎点种腐病菌(Phomopsislongicolla)有一定的差别,可能是因为猕猴桃品种(翠玉)不一样,或来自不同地区果园所致,在今后试验中可进一步对来自不同地区的软腐红阳猕猴桃进行病原鉴定,在验证上述假设的同时以期得到更多不同新种类的病原菌。相对于陆训等[15]、王小洁等[18]的研究发现,中华系列猕猴桃感染软腐病的症状(病斑)更加明显,可能是因为美味系列猕猴桃有毛。
中华系列猕猴桃口感佳、鲜食效果更好,经济效益更高,因此今后应着重对中华系列软腐猕猴桃病原菌进行研究。同时王小洁等[18]研究表明这两种菌具有较强抵抗低温的能力,能产出大量的孢子,尤其是间座壳菌,因此猕猴桃种植过程中,需要进行严格的冬季清园处理,杀灭这些软腐病原菌,以免对果子造成侵染。
本研究通过改变外部环境因子对猕猴桃葡萄座腔菌和间座壳菌的生长特性进行分析,发现在pH值5~11范围内病原菌能正常繁衍,同时在中性偏酸的条件下生长最好,这与其寄主猕猴桃果实存在的环境条件基本吻合。菌株对于恶劣温度和pH值环境的适应性较强,存活能力高,在80℃水浴处理30 min或者在pH值4或13的环境下依然能生长。说明病原菌对外界环境具有较强的抗性,这与病原菌自身的因素有关,通过改变环境因素杀灭真菌有一定难度,今后研究可开发相关植物源杀菌剂,对此类病原菌进行杀菌试验。本研究结果表明,在温度25~30℃的环境下,菌丝生长旺盛,菌丝密集度较高,且其生长发育的最适温度和果实采收时期的气温相近,与同属内其他菌种大致相似[21,24]。在采果期间应尽量选择清晨或傍晚温度较低时,一方面减少果实呼吸代谢,一方面钝化病原菌的生长,延长保鲜时间。病菌更喜营养成分均衡且多含有葡萄糖、蔗糖、麦芽糖的培养基,如PSA、YPGA、PDA、SDA。对于氮源来说,两种菌种均偏爱酵母粉、蛋白胨、甘氨酸等,两种菌均能利用多种营养,今后对这两种菌进行扩繁时,可参考此理论依据。此外,未来研究可进一步补充惰性气体、紫外光照、辐射等物理手段对病原菌生长特性的影响[25-26],以期得到防控软腐病原菌的良好措施。
在研究猕猴桃软腐病原菌生物学特性的基础上,需进一步鉴定出属内种间的差异性,从而精准鉴别出每种植物病害的病原菌,分析和完善地域间种群的亲缘关系[27];深入探究致病机理,了解和把握每个环节的致病过程,形成一套系统而全面的认识体系,构建抗病分子育种技术体系,从根本上防控病害的发生[28]。
4 结论
本研究鉴定出造成猕猴桃果实采后软腐的主要病害菌为葡萄座腔菌(Botryosphaeriadothidea)和间座壳菌(Diaporthephaseolorum),这两种病原菌在PSA、YPGA、PDA、SDA上生长速度较快,菌丝较稠密。同时,两种病原菌对葡萄糖、淀粉、蔗糖和麦芽糖等碳源利用效果好,对酵母粉、蛋白胨、甘氨酸等氮源利用效果好;最适pH值为5~7,最适温度为25~30℃,致死温度为85℃。猕猴桃采后保存时,减少猕猴桃储运、贮藏过程的中的机械损伤可减轻和延缓果实软腐,适当的碱性环境及0.5~4℃的低温贮藏,有助于抑制病原菌,从而延长果实的贮藏期。本研究结果为进一步阐明猕猴桃果实软腐病害主要病原菌的种类及对猕猴桃贮藏过程中病害的绿色防控供了一定理论依据。
猜你喜欢
油气田地面工程(2022年9期)2022-10-26
中国土壤与肥料(2021年5期)2021-12-02
昆钢科技(2021年6期)2021-03-09
当代水产(2020年11期)2021-01-04
农业与技术(2017年12期)2017-07-11
农民致富之友(2017年7期)2017-04-27
湖北农业科学(2016年23期)2017-03-17
农家顾问(2016年12期)2017-01-06
分析化学(2014年10期)2014-10-24
中国科技纵横(2012年2期)2012-02-27
