
三种豆科绿肥作物茎和叶角质层蜡质化学组成分析
2020-12-11 02:47姚露花王登科张学风肖前林杨瑞吉郭彦军
作物学报 2020年1期
栗 扬 姚露花 郭 欣 赵 晓 黄 蕾 王登科 张学风 肖前林 杨瑞吉 郭彦军
三种豆科绿肥作物茎和叶角质层蜡质化学组成分析
栗 扬 姚露花 郭 欣 赵 晓 黄 蕾 王登科 张学风 肖前林 杨瑞吉 郭彦军*
西南大学农学与生物科技学院, 重庆 400716
植物角质层蜡质是一类覆盖于植物表层的疏水有机化合物, 在保护植物免受生物与非生物逆境胁迫中发挥着重要作用。为了更好地了解和认识角质层蜡质在夏季绿肥作物抗逆性中的作用, 选择柽麻()、田菁()和竹豆() 3种夏季豆科绿肥作物, 鉴定茎和叶蜡质组分, 并分析蜡质总量、各组分含量及碳链分布特征。共鉴定出8类化合物, 包括脂肪酸、初级醇、醛、烷烃、烷基酯、二醇、萜类和固醇类化合物, 其中前4种以同系物形式存在且为所有植物茎和叶共有成分(柽麻茎中未检出脂肪酸), 说明烷合成和醇合成途径是主要的2种蜡质合成途径。田菁茎中鉴定出二醇化合物, 其结构初步解析为1,18-30烷醇和1,16-30烷醇。3种绿肥作物茎和叶蜡质总量存在显著种间及部位差异, 其中柽麻茎蜡质总含量为16.33 µg cm-2, 显著高于田菁茎(6.45 µg cm-2)和竹豆茎(0.72 µg cm-2)。就茎和叶比较, 柽麻茎显著高于叶片, 其他2种植物茎和叶之间无显著差异。柽麻茎蜡质中, 烷烃为优势成分, 占蜡质总量的57.38%; 叶片以初级醇为优势成分, 占蜡质总量的50.12%。田菁茎、叶蜡质中的优势成分均为初级醇, 分别占总蜡质的30.12%和71.21%。竹豆茎、叶蜡质中的优势成分均为烷烃, 分别占总蜡质的40.79%和39.27%。各组分优势化合物的碳链长度在不同物种、不同部位也存在一定差异, 说明参与蜡质合成的基因在物种、器官间有所不同。这些结果为今后从分子水平上揭示角质层蜡质参与夏季绿肥作物抗逆机制提供了理论基础。
角质层蜡质; 绿肥作物; 柽麻; 田菁; 竹豆
绿肥作物种植于主要作物种植间隙, 是一类以改良土壤质量为目的的植物, 其中以豆科绿肥作物利用较广[1]。豆科绿肥作物既可通过生物固氮增加土壤中的氮素, 也可通过翻埋入土, 以绿肥的形式改良土壤质量。然而, 相较于粮食作物, 对夏季绿肥作物基本理化特性的认识尚不足, 相关研究文献也少, 制约了研究者对这些绿肥作物的开发利用。因此, 了解夏季绿肥作物的抗性生理指标, 可为提高这类作物的抗逆性、降低田间管理成本提供理论依据。
植物角质层蜡质是植物与外界环境的第一接触面, 在保护植物免受生物及非生物胁迫的侵害、提高抗逆性方面发挥着非常重要的作用[2]。如干旱胁迫下, 植物叶角质层蜡质含量的增加被认为与提高植物的抗旱性有关[3]。随着紫外线辐射强度的变化, 角质层蜡质晶体结构也随之发生变化, 以通过反射减少紫外线伤害[4]。角质层蜡质中的醛类物质被认为与病原菌的侵染有关[5]。此外, 朱命炜等[6]和韦存虚等[7]应用扫描电镜观察发现成熟芦荟()和星星草()叶表面分布有密集的瘤状蜡质, 能有效地减少水分散失和增强叶片反射光辐射的能力。目前的研究结果表明, 植物角质层蜡质主要由长链脂肪酸及其衍生物组成, 如醛类、初级醇、酯类、烷烃、次级醇、酮, 及支链醇、支链烷烃、三萜醇、双酮、烯烃、二醇和酮醇等物质[8]。植物种类不同, 其所含的角质层蜡质组分也不同, 如油菜()和番茄()以烷类物质为优势成分, 紫花苜蓿()、玉米()以初级醇为优势成分[9], 羊草()以二酮为优势成分[10]。蜡质组分的品种差异说明参与角质层蜡质生物合成及转运的途径也可能存在差异[11]。角质层蜡质的合成, 首先在细胞质中由可溶性的脂肪酸合成酶催化从头合成16C/18C的饱和脂肪酸, 然后在内质网中由脂肪酰-CoA延长酶催化16C/18C的脂肪酸延长产生不同链长的长链脂肪酸。脂肪酰-CoA延长酶是一个多酶复合体, 包括β-酮脂酰-CoA合酶(3-ketoacyl- CoA synthase, KCS)、β-酮脂酰-CoA还原酶(3-ketoacyl-CoA reductase, KCR)、β-羟脂酰-CoA脱水酶(3-hydroxacyl-CoA dehydratase, HCD)和反式烯脂酰-CoA还原酶(trans-2,3-enoyl-CoA reductase, ECR)。基于蜡质组分分析及基因功能验证, 目前已证实, 角质层蜡质生物合成主要包括脱羰基途径(烷合成途径)、酰基还原途径(初级醇合成途径)和聚酮合酶途径(polyketide synthase, PKS)[12]。如在拟南芥()中已鉴定出参与蜡质合成的21个基因、参与合成次级醇和酮的MAH1基因及参与烷基酯合成的基因[12], 在小麦()与大麦()中鉴定出参与β-二酮合成的基因[13]。挖掘蜡质合成基因已被作为选育抗逆作物材料的方法之一[9]。如在紫花苜蓿中的超表达, 显著增加了烷烃含量, 提高了植株的抗旱性[13],在水稻()中参与了初级醇的合成, 并可提高水稻抗旱性[15]。
柽麻()、竹豆()、田菁()是生产中常用的几种夏季豆科绿肥作物。柽麻与玉米间作, 可显著提高玉米地上部产量[16]。在松嫩平原盐碱土, 随着田菁生物量增加, 土壤pH、可溶性盐含量逐年降低, 有机质含量、碱解氮含量逐年升高[17]。幼龄果园套种竹豆绿肥与清耕相比, 可减少径流70%、泥沙80%以上, 对改善园地土壤水肥, 促进幼树生长有明显的促进作用[18]。目前尚无这些绿肥作物角质层蜡质方面的研究报道。鉴于此, 本试验从田间采集柽麻、田菁和竹豆叶片及茎秆, 利用色谱质谱联用仪鉴定了蜡质组分, 用气相色谱对蜡质组分进行了定量分析, 并计算了各组分相对含量及碳链分布特征, 为进一步挖掘这些植物的蜡质合成基因, 提高夏季豆科绿肥作物抗逆性提供理论基础。
1 材料与方法
1.1 试验地点
试验地位于西南大学农学与生物科技学院实验农场(29°48′N, 106°24′E), 属亚热带季风性湿润气候,年平均气温为18.2°C, 降水量为1084.6 mm。夏季绿肥作物生长季(4月至9月)月平均气温分别为18.9°C、22.5°C、25.3°C、28.6°C、28.5°C和24.3°C; 平均降水量分别为94.6、144.2、206.3、169.4、128.6和126.6 mm; 月平均日照时数分别为96.8、103.8、97.2、162.5、164.3和104.7 h (1988—2017年)。供试土壤为粘质黄壤, 0~20 cm土壤耕层pH 6.37, 含有机质 12.4 g kg-1、全氮0.73 g kg-1、全磷0.12 g kg-1、全钾3.32 g kg-1、碱解氮67.4 mg kg-1、速效磷8.6 mg kg-1、速效钾82.5 mg kg-1。
1.2 绿肥作物
供试绿肥作物包括柽麻()、竹豆()和田菁()。种子经消毒(10% H2O2)后, 直接播在田间。设置9个小区, 小区面积10 m2(2 m×5 m), 每个品种3个小区(重复), 随机区组排列。播种时, 行距40 cm, 株距30 cm。等幼苗出土1周后, 开始间苗, 每小区留苗76株。定期除杂, 不浇水(靠雨水)。待植物进入现蕾期开始采样分析。因竹豆在重庆地区开花较迟且少, 采样时只有少数植株有花蕾。
1.3 蜡质提取
分别采集每个小区各植物不同植株相同叶位叶片5片, 同时按留茬5 cm刈割后, 剪取茎段。提蜡前, 利用WinFOLIA专业组织图像分析系统(Regent Instrument Inc, Canada)和数字化扫描仪(EPSON V750, Japan)测定茎、叶的正投影面积, 而3种豆科绿肥作物的茎可近似看作圆柱体, 故茎、叶面积可通过以下公式进行计算。
叶片面积= 2×投影面积
茎秆表面积= π×投影面积
每个小区叶片和茎秆样品, 在室温条件下分别用含5 µg内标(24烷)的氯仿(5 mL)涡旋浸提3次, 每次30 s。提取液混合后经40°C氮气吹干, 加入20 µL BSTFA [N,O-Bis (三甲基硅烷基)三氟乙酰胺])和20 µL吡啶, 于70°C衍生反应45 min, 多余衍生剂用氮吹仪再次吹干。将所得提取物溶于1 mL氯仿, 进行气相色谱和质谱分析。
1.4 气相色谱-质谱(GC-MS)分析
采用9790II气相色谱仪(浙江福立)分析蜡质含量。色谱柱为DM-5 (5%二苯基 + 95%二甲基聚硅氧烷固定液), 毛细管柱长30 m, 直径0.32 mm, 液膜厚度0.25 µm。氮气作为载气; 进样量为1.0 µL; 柱膜和FID检测器的温度分别为300°C和320°C; 分流比为10︰3。初始温度为80°C, 以每分钟15°C的速度升温至260°C, 并保持10 min, 之后以每分钟2°C升温至290°C, 保持1 min; 再以每分钟5°C的速度升温至320°C, 并保持10 min。利用气相色谱—质谱联用仪(GCMS-AOC-5000 Plus)鉴定蜡质组分, 程序升温方式同色谱分析。鉴定成分时主要根据离子峰质荷比(m/z)结合已发表相关成分质谱图, 解析化合物结构。基于FID峰值量化蜡质, 根据质谱所鉴定的标准确定色谱中各种化学成分的出峰位置, 用内标计算出植物单位面积的实际蜡质含量, 单位为µg cm-2。
1.5 数据分析
试验数据为3个重复的平均值±标准误。采用SPSS(17.0)统计软件, 利用单因素方差分析(One- Way ANOVA)比较不同作物茎、叶的蜡质总含量、蜡质各组分含量及链长分布特征, 显著性检验水平为= 0.05 (l.s.d.)。
2 结果与分析
2.1 蜡质组分鉴定
以质谱分析获得化合物离子峰质荷比(m/z), 再结合文献中对蜡质成分的鉴定质谱图, 从柽麻茎中鉴定出5类物质, 包括脂肪酸、初级醇、醛类、烷烃和萜类化合物, 叶片中鉴定出4类物质, 包括初级醇、醛类、烷烃和萜类化合物。田菁茎中发现7类物质, 包括脂肪酸、初级醇、烷基酯、醛类、烷烃、二醇、萜类和固醇类化合物, 叶片中鉴定出5类物质, 包括脂肪酸、初级醇、醛类、烷烃和萜类。其中二醇为1,18-30烷醇和1,16-30烷醇, 其中1,16-30烷醇离子峰构成为m/z 73(100%)、103(38%)、149(59%)、285(23%)、299(63%)、313(25%)、387(8%)、401(15%)、415(6%)、598(质量峰)(图1)。竹豆茎和叶中均鉴定出5类物质, 包括脂肪酸、初级醇、醛类、烷烃和萜类。柽麻和田菁茎和叶均有低于10%左右的成分未能被鉴定, 而竹豆茎和叶中未鉴定成分高达30%左右。
2.2 茎、叶蜡质总量及组分相对含量分析
柽麻、田菁、竹豆3种绿肥作物的茎、叶蜡质总含量存在显著种间及部位差异, 其中柽麻茎蜡质总含量为16.33 µg cm-2, 显著高于田菁茎(6.45 µg cm-2)和竹豆茎(0.72 µg cm-2)(图2)。就茎和叶比较, 柽麻茎显著高于叶片, 其他2种植物茎和叶之间无显著差异。
柽麻茎蜡质中, 烷烃为优势成分, 占蜡质总量的57.38%, 其次是初级醇, 占14.23%, 脂肪酸占11.23%, 醛类和萜类化合物均低于7%, 未鉴定蜡质组分约占2.96% (图3)。叶片中初级醇为优势成分, 占蜡质总量的50.12%, 醛类和萜类化合物分别占12.77%和15.95%, 而烷烃比例只有5.26%, 约12.45%的成分未能鉴定出来。

图1 田菁茎蜡质中双醇化学结构分析
A: 1,18 -30双醇; B: 1,16 30-双醇。
A: C30 1,18-alkanediol; B: C30 1,16-alkanediol.
田菁茎、叶蜡质中的优势成分均为初级醇, 其中茎中占总蜡质的30.12%, 而叶中高达71.21%。其次是醛类, 茎和叶中分别占总蜡质的22.89%和19.39%(图4)。茎中发现的二醇, 所占比例为3.58%, 烷基酯比例为0.44%。脂肪酸、烷烃和三萜类物质在茎和叶中比例分别为7.99%和3.86%、6.59%和3.29%、14.63%和0.11%。另外, 茎和叶中均有8.55%和2.13%的未知成分。

图2 柽麻、田菁、竹豆的茎和叶蜡质总量
数据柱上不同小写字母表示品种内差异显著(< 0.05)。
Different lowercase letters above the data bar within each species represent significant difference at< 0.05.

图3 柽麻茎和叶中主要蜡质组分实际含量
竹豆茎、叶蜡质中的优势成分均为烷烃, 分别占总蜡质的40.79%和39.27%。其次茎中分别为醛(25.28%)、初级醇(12.18%)和三萜类(2.61%), 叶中三类组分的比例分别为8.34%、3.75%和12.69% (图5)。茎和叶中均有小于2%的脂肪酸, 以及17.22%和22.66%的未知成分。

图4 田菁茎和叶主要蜡质组分实际含量

图5 竹豆茎和叶主要蜡质组分实际含量
2.3 碳链分布特征分析
不同绿肥作物中发现的蜡质组分多数以同系物的形式存在, 如脂肪酸和初级醇均为不同链长的偶数碳分子, 而醛和烷烃为奇数和偶数碳共存的化合物, 且烷烃表现为明显的奇数优势, 醛表现为偶数优势。
2.3.1 柽麻主要蜡质组分碳链分布特征 柽麻茎中脂肪酸碳链分布在C20~C32之间, 其中以C30为优势组分, 占总脂肪酸的33.33%, 其次为C32, 占22.20% (图6)。初级醇碳链分布茎中在C22~C34之间, 叶中在C24~C32之间, 二者均以C28为优势组分, 分别占总初级醇的45.34%和68.66%; 茎中含有较高比例的C32, 达到31%, 而叶中只有0.14%。茎和叶中醛的碳链分布存在较大差异, 茎中以C32为优势组分(52.13%), 叶中以C30为优势组分(70.83%)。烷烃链长在C25~C33之间, 茎和叶均以C31为优势组分, 分别占总烷烃的79.49%和59.56%。萜类物质, 茎和叶中均以羽扇豆醇为最高(80%左右), 其中茎中还有20%左右a-香树脂醇, 叶中有15%左右的乙酰化羽扇豆醇。
2.3.2 田菁主要蜡质组分碳链分布特征 田菁茎和叶中的脂肪酸碳链分布范围在C20~C32, 其中茎中以C30为优势组分(68.35%), 而叶中以C26为优势组分(30.15%)(图7)。初级醇碳链分布在茎和叶中差异较大, 其中茎中在C26~C32, 叶中在C22~C32, 但2个部位均以C30为优势组分, 分别占83.05%和73.25%。醛碳链分布茎中在C28~C34, 叶中在C28~C32, 2个部位均以C30为优势组分, 分别占46.96%和52.53%。烷碳链分布茎中在C25~C33, 叶中在C25~C31, 2个部位均以C29为优势组分, 分别占41.77%和51.18%。茎中所鉴定的二醇为C30同分异构体, 其中1,18-30烷醇占27.91%, 1,16-30烷醇占72.09%。茎中发现的萜类物质为β-香树脂醇和羽扇豆醇, 而叶中只有β-香树脂醇。
2.3.3 竹豆主要蜡质组分碳链分布特征 竹豆茎和叶中脂肪酸碳链分布均在C22~C30之间, 且均以C28为优势组分(60%左右)(图8)。初级醇碳链分布, 茎中在C24~C30, 叶中在C24~C34, 其中茎中以C28为优势组分(68.75%), 叶中以C32为优势组分(60.38%)。茎中醛碳链在C28~C32, 其中C30为优势组分(88.75%), 而叶中只有C28醛。茎和叶中烷烃碳链分布均在C25~ C33之间, 且均以C31为优势组分, 分别占40.75%和60.26%。茎和叶中的萜类物质均为β-香树脂醇和羽扇豆醇, 其中茎以β-香树脂醇为优势组分(73.69%), 叶以羽扇豆醇为优势组分(72.97%)。
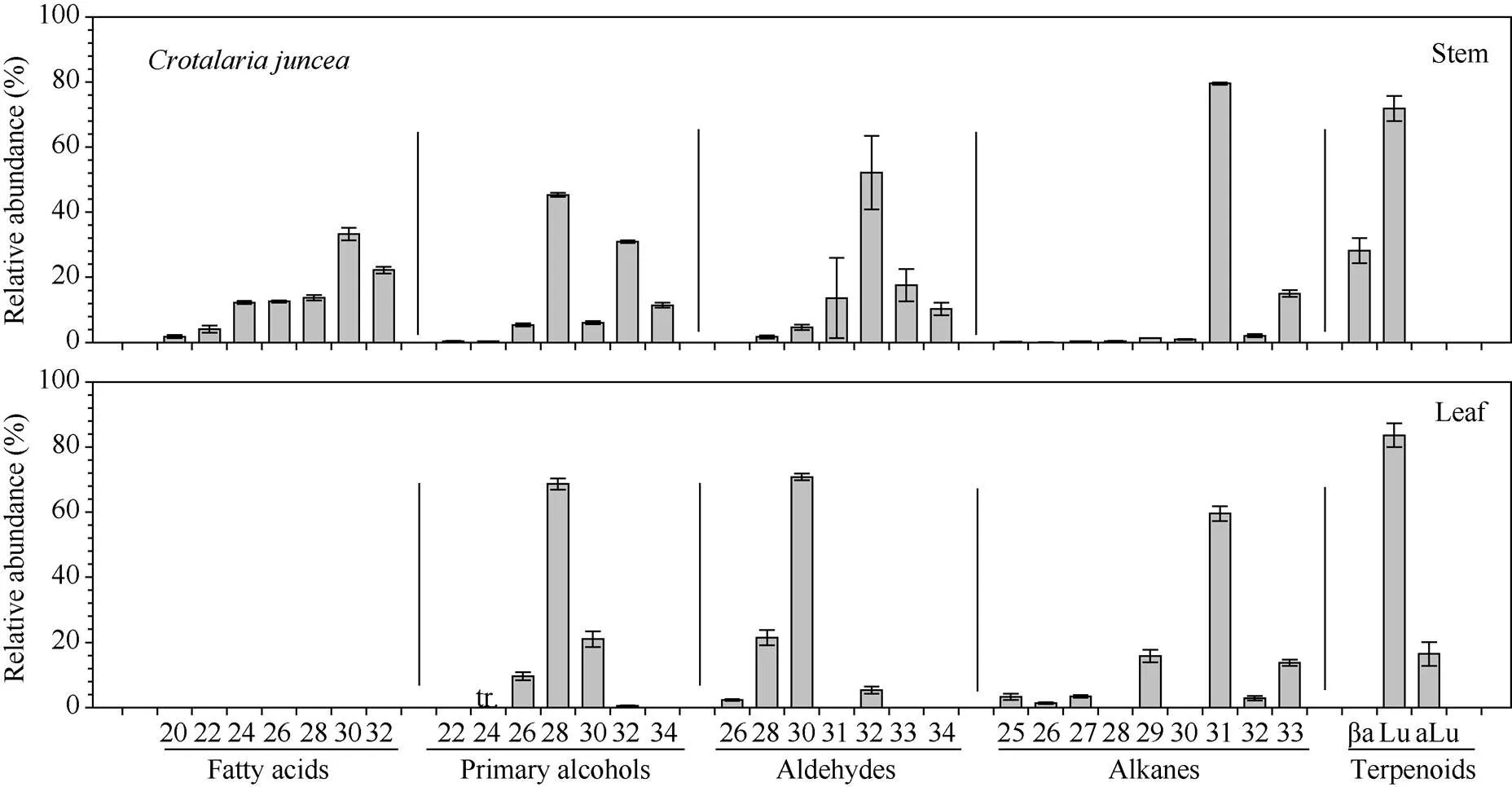
图6 柽麻茎和叶蜡质各组分同系物相对含量

图7 田菁茎和叶蜡质各组分同系物相对含量
3 讨论
3种豆科绿肥作物的茎和叶中均含有脂肪酸、初级醇、醛类和烷烃,且这些组分含量总和占总蜡质含量的比例超过60%。说明烷合成途径和醇合成途径是这些植物合成角质层蜡质的主要途径。这与在其他主要作物上的发现是一致的[9]。然而,蕨类植物叶角质层蜡质以烷基酯为优势成分[19], 羊草以二酮为优势成分[10]。这些结果暗示了, 在长期进化过程中, 不同植物为适应不同的生存环境其角质层蜡质生物合成产生了不同的进化方向。此外,我们还发现田菁茎和叶中的蜡质组分存在较大差异, 有些成分只出现在叶片, 如二醇和固醇类物质, 其中二醇为1,18-30烷醇和1,16-30烷醇。在高山罂粟()叶片中也鉴定出C-30的二醇同系物, 但其羟基位置在1,11[20], 在蓖麻()叶片中,鉴定出链长C-22到C-28、羟基位置在1,3的二醇同系物[21]。Busta和Jetter[22]认为这些碳链一头为初级醇、碳链中间存在另一个羟基的二元醇结构,其生物合成过程可能是由P450加氧酶参与完成的。在拟南芥()中,MAH1作为羟化酶参与了二醇的合成[23]。田菁叶中二醇物质的发现,为今后研究二醇生物合成途径提供了新的材料。

图8 竹豆茎和叶蜡质各组分同系物相对含量
3种豆科绿肥作物茎和叶中蜡质含量存在较大差异, 其中茎蜡质含量最高的是柽麻, 其次是田菁, 竹豆蜡质含量最低。一般情况下, 植物蜡质含量的增加与其抗旱性的提高相联系[2]。尽管都是夏季绿肥作物, 蜡质含量较低的竹豆为藤本植物, 群体高度低、叶片数量多、地表覆盖度高, 在进化过程中其生长环境可能相对湿润, 不需要过高的蜡质沉积就能适应生长环境。而柽麻与田菁株型直立, 地表覆盖度低, 生长环境相对干燥, 叶片可能需要沉积较多的蜡质来抵御逆境胁迫。当然, 影响植物抗逆性的因素较多, 今后有必要进一步分析几种植物蜡质含量与其抗旱性的关系。就茎和叶角质层蜡质含量比较, 拟南芥茎秆高于叶片[24], 马铃薯()无显著差异[25], 蒲公英()叶片高于茎秆[26], 本试验中柽麻茎高于叶片, 而田菁和竹豆茎和叶蜡质无显著差异。这些结果说明不同植物茎和叶角质层蜡质的沉积存在器官特异性, 且这可能与各自不同的生理功能有关。对12个大麦品种的比较发现, 蜡质沉积较多的品种具有较高的抗旱性[3]。转录因子MYB96在亚麻荠()中的超量表达可显著增加蜡质沉积, 并提高植物抗旱性[27]。
在茎和叶蜡质组分方面, 柽麻茎以烷烃为优势组分, 而叶以初级醇为优势组分, 说明参与蜡质合成的主效基因在2个器官的表达存在差异, 如拟南芥参与初级醇合成的CER4在叶中较高表达, 而参与烷合成的CER1在茎中高表达[28]。田菁茎和叶中的优势组分均为初级醇(30.12%和71.21%), 且烷烃组分比例不足10%, 说明田菁角质层蜡质以醇合成途径为主。竹豆茎和叶中的优势成分均为烷烃, 说明其角质层蜡质合成以烷合成途径为主。在其他作物上也有类似的研究结果, 如紫花苜蓿叶以初级醇为主, 茎以烷为主[14]; 亚麻荠()叶以烷基酯为主, 茎以三萜类化合物为主[29]; 而玉米叶以初级醇为主, 茎以醛为主[30]。尽管目前尚无法确定这些优势组分是否在植物抗逆方面起主要作用, 然而干旱胁迫下烷类含量的增加被认为与多数植物的抗旱性提高有关[2]; 植物叶表长链醛类的缺失可抑制病原真菌孢子的萌发[31]。
3种绿肥作物茎和叶中主要的蜡质组分均以同系物形式存在, 且整体表现出脂肪酸、初级醇和醛以偶数碳占优势、而烷烃以奇数碳占优势的碳链分布特征。这与绝大多数植物上的研究结果是一致的[32]。就蜡质组分内同系物优势化合物碳链长度分析, 柽麻和竹豆茎和叶均以C31为优势化合物, 而田菁以C29为优势化合物。说明参与烷烃生物合成的同源基因在茎和叶中是一致的。茎和叶初级醇优势化合物在柽麻和田菁是一致的, 分别为C28和C30; 而竹豆茎中以C28为优势化合物, 叶中以C32为优势化合物。出现这一结果的原因可能在于链长延长酶(FAE)及参与初级醇合成的CER4在竹豆不同器官存在差异[33]。醛被认为是烷合成途径和醇合成途径的中间产物[9]。几种绿肥作物茎和叶中醛的优势化合物存在较大差异, 暗示不同器官的醛可能来自不同的合成途径。
4 结论
从3种夏季绿肥作物茎和叶中共鉴定出8类化合物, 包括脂肪酸、初级醇、烷基酯、醛类、烷烃、二醇、萜类和固醇类化合物。尽管主要蜡质组分在品种及部位间一致, 但不同作物及部位又有各自独特的组分存在, 如柽麻茎中未检测出脂肪酸, 田菁茎中鉴定出固醇类和二醇化合物。其中田菁茎中的二醇化合物初步结构解析为1,18-30烷醇和1,16-30烷醇。此外, 不同物种、不同部位所积累的优势组分及优势化合物也存在一定差异, 说明参与蜡质合成的基因在物种、器官间有所不同。这为今后从分子水平上揭示角质层蜡质参与夏季绿肥作物抗逆机制提供了理论基础。
[1] N’Dayegamiye A, Tran T S. Effects of green manures on soil organic matter and wheat yields and N nutrition., 2001, 81: 371–382.
[2] Yeats T H, Rose J K C. The formation and function of plant cuticles., 2013, 163: 5–20.
[3] Gonzalez A, Ayerbe L. Effect of terminal water stress on leaf epicuticular wax load, residual transpiration and grain yield in barley., 2010, 172: 341–349.
[4] Skorska E, Szwarc W. Influence of UV-B radiation on young triticale plants with different wax cover., 2007, 51: 189–192.
[5] Zhu M, Riederer M, Hildebrandt U. Very-long-chain aldehydes induce appressorium formation in ascospores of the wheat powdery mildew fungus., 2017, 121: 716–728.
[6] 朱命炜, 王红星, 李建军, 李俊英, 王太霞, 李景原. 木立芦荟发育过程中叶表皮角质膜和蜡质的变化. 电子显微学报, 2004, 23: 670–673. Zhu M W, Wang H X, Li J J, Li J Y, Wang T X, Li J Y. Changes in the cuticle and wax of the leaves during the development of aloe (Mill.)., 2004, 23: 670–673 (in Chinese with English abstract).
[7] 韦存虚, 王建波, 陈义芳, 周卫东, 孙国荣. 盐生植物星星草叶表皮具有泌盐功能的蜡质层. 生态学报, 2004, 24: 2451–2456. Wei C X, Wang J B, Cheng Y F, Zhou W D, Sun G R. Epicuticular wax of leaf epidermis: a functional structure for salt excretion in a halophyte., 2004, 24: 2451–2456 (in Chinese with English abstract)
[8] Jetter R, Riederer M. Composition of cuticular waxes onFronds., 2000, 26: 399–412.
[9] Lee S B, Suh M C. Advances in the understanding of cuticular waxes in Arabidopsis thaliana and crop species., 2015, 34: 557.
[10] 李晓婷, 赵晓, 王登科, 黄蕾, 姚露花, 王党军, 和玉吉, 郭彦军. 天然草地植物叶角质层蜡质的化学组成及其对自由放牧的响应. 草业学报, 2018, 27(6): 137–147.Li X T, Zhao X, Wang D K, Huang L, Yao L H, Wang D J, He Y J, Guo Y J. Chemical profiles of cuticular waxes in arid steppe plant species and their responses to continuous grazing., 2018, 27(6): 137–147 (in Chinese with English abstract).
[11] Bernard A, Joubès J. Arabidopsis cuticular waxes: advances in synthesis, export and regulation., 2013, 52: 110–129.
[12] Kunst L, Samuels L. Plant cuticles shine: advances in wax biosynthesis and export., 2009, 12: 721–727.
[13] Kosma D K, Rowland O. Answering a four decade-old question on epicuticular wax biosynthesis., 2016, 67: 2538–2540.
[14] Zhang J Y, Broeckling C D, Blancaflor E B, Sledge M K, Sumner L W, Wang Z Y. Overexpression of WXP1, a putativeAP2 domain-containing transcription factor gene, increases cuticular wax accumulation and enhances drought tolerance in transgenic alfalfa ()., 2005, 42: 689–707.
[15] Zhou L Y, Ni E D, Yang J W, Zhou H, Liang H, Li J, Jiang D G, Wang Z H, Liu Z L, Zhuang C X. Rice OsGL1-6 is involved in leaf cuticular wax accumulation and drought resistance., 2013, 8: 12.
[16] 杜青峰, 王党军, 于翔宇, 姚露花, 和玉吉, 王瑞, 马生兰, 郭彦军. 玉米间作夏季绿肥对当季植物养分吸收和土壤养分有效性的影响. 草业学报, 2016, 25(3): 225–233.Du Q F, Wang D J, Yu X Y, Yao L H, He Y J, Wang R, Ma S L, Guo Y J. The effects of corn and green manure intercropping on soil nutrient availability and plant nutrient uptake., 2016, 25(3): 225–233 (in Chinese with English abstract)
[17] 张国发, 吴园园, 徐太海, 梁彦涛. 田菁秸秆还田对松嫩平原盐碱土改良效果的研究. 大庆师范学院学报, 2018, 38(3): 48–50. Zhang G F, Wu Y Y, Xu T H, Liang Y T. The effects of returningstraw on improving soil quality in alkalized soils of Songnen Plain.2018, 38(3): 48–50 (in Chinese).
[18] 杨湘如, 郑永发. 幼龄果园套种竹豆生态效益试验研究. 中国水土保持, 1991, (11): 29–30. Yang X R, Zheng Y F. The ecological profits ofintercropping with young orchard., 1991, (11): 29–30 (in Chinese).
[19] Guo Y, Li J J, Busta L, Jetter R. Coverage and composition of cuticular waxes on the fronds of the temperate fernsand., 2018, 122: 555–568.
[20] Jetter R, Riederer M, Seyer A, Mioskowski C. Homologous long-chain alkanediols from Papaver leaf cuticular waxes., 1996, 42: 1617–1620.
[21] Vermeer C P, Nastold P, Jetter R. Homologous very-long-chain 1,3-alkanediols and 3-hydroxyaldehydes in leaf cuticular waxes ofL., 2003, 62: 433–438.
[22] Busta L, Jetter R. Moving beyond the ubiquitous: the diversity and biosynthesis of specialty compounds in plant cuticular waxes., 2018, 17: 1275–1304.
[23] Wen M, Jetter R. Composition of secondary alcohols, ketones, alkanediols, and ketols incuticular waxes., 2009, 60: 1811–1821.
[24] Hegebarth D, Buschhaus C, Wu M, Bird D, Jetter R. The composition of surface wax on trichomes ofdiffers from wax on other epidermal cells., 2016, 88: 762–774.
[25] Guo Y J, Jetter R. Comparative analyses of cuticular waxes on various organs of potato (L.)., 2017, 65: 3926–3933.
[26] Guo Y J, Busta L, Jetter R. Cuticular wax coverage and composition differ among organs of., 2017, 115: 372–379.
[27] Lee S, Kim H, Kim R, Suh M. Overexpression of Arabidopsis MYB96 confers drought resistance invia cuticular wax accumulation., 2014, 33: 1535–1546.
[28] Rowland O, Zheng H, Hepworth S R, Lam P, Jetter R, Kunst L. CER4 encodes an alcohol-forming fatty acyl-coenzyme a reductase involved in cuticular wax production in Arabidopsis., 2006, 142: 866–877.
[29] Razeq F M, Kosma D K, Rowland O, Molina I. Extracellular lipids of: characterization of chloroform- extractable waxes from aerial and subterranean surfaces., 2014, 106: 188–196.
[30] Javelle M, Vernoud V, Depege-Fargeix N, Arnould C, Oursel D, Domergue F, Sarda X, Rogowsky P M. Overexpression of the epidermis-specific homeodomain-leucine zipper IV transcription factor Outer Cell Layer1 in maize identifies target genes involved in lipid metabolism and cuticle biosynthesis., 2010, 154: 273–286.
[31] Hansjakob A, Riederer M, Hildebrandt U. Appressorium morphogenesis and cell cycle progression are linked in the grass powdery mildew fungus., 2012, 116: 890–901.
[32] Jetter R, Kunst L, Samuels A L. Composition of plant cuticular waxes. In: Riederer C. Muller, eds. Biology of the Plant Cuticle. Oxford: Blackwell Publishing Ltd, 2006. pp 145–181.
[33] Haslam T M, Mañas-Fernández A, Zhao L, Kunst L. Arabidopsis ECERIFERUM2 is a component of the fatty acid elongation machinery required for fatty acid extension to exceptional lengths., 2012, 160: 1164–1174.
Chemical compositions of cuticular waxes on stems and leaves of three legume green manure crops
LI Yang, YAO Lu-Hua, GUO Xin, ZHAO Xiao, HUANG Lei, WANG Deng-Ke, ZHANG Xue-Feng, XIAO Qian-Lin, YANG Rui-Ji, and GUO Yan-Jun*
College of Agronomy and Biotechnology, Southwest University, Chongqing 400716, China
Cuticular waxes are hydrophobic compounds covering plant surfaces and play vital roles in protecting plants from various biotic and abiotic stresses. However, less is known as related to the cuticular wax on leaves or stems of summer legume green manures. In the current study, stems and leaves from three summer legume green manure crops grown in the fields were sampled, including,, andRoxbwere.Using gas chromatography-mass spectrometer (GC-MS), we identified most of the chemical compounds on these plant species. In total, eight classes of wax compounds were identified, including fatty acids, primary alcohols, aldehydes, alkanes, alkyl esters, diols, terpenoids and sterols. Among these compounds, fatty acids, primary alcohols, aldehydes and alkanes with serial homologs could be observed in stems and leaves of all tested plant species excepting fatty acids in leaves of, with their weight proportions accounting for more than 60% in total wax. This suggested that alkane forming and alcohol forming pathways were two major biosynthesis pathways in these plants. In stems of, two compounds were identified as diols with the functional hydroxyl group located at 1,18 and 1,16 positions. Using gas chromatography with flam ionized detector (GC/FID), we quantified the total wax and wax compounds. Total wax coverage differed among three plant species and between two organs. The total stem wax coverage was the highest in(16.33 µg cm–2), followed by(6.45 µg cm–2) and(0.72 µg cm–2). The total wax coverage on stems ofwas significantly higher than that on leaves, whereas no significant difference in total wax coverage between stems and leaves was observed inand. The predominant wax class differed in plant species and organs. Foralkanes were the predominant composition in stems accounting for 57.38% of total wax, whereas primary alcohols were the predominant composition in leaves accounting for 50.12%. For, both stems and leaves were dominated by primary alcohols, accounting for 30.12% and 71.21% of total wax, respectively. For, both stems and leaves were dominated by alkanes, accounting for 40.79% and 39.27% of total wax, respectively. We further analyzed the chain length distributions of the wax classes in stems and leaves. Generally, fatty acids, primary alcohols and aldehydes were consisted of serials of even carbon number homologs, whereas alkanes were consisted of both even- and odd-carbon number homologs with odd number predominance over even number. The predominant compound in each wax class also differed between plant species and organs. The dominant fatty acids on stems and leaves in,, andwere C30, C30and C26, and C28, respectively; the dominant primary alcohols were C28, C30, and C28and C32respectively; the dominant aldehydes were C32and C30, C30, and C30and C28, respectively; while the dominant alkanes were C31, C29, and C31, respectively. The terpenoids identified in current study were β-amyrin,a-amyrin and lupenol. The variations of predominant wax class and wax compound between plant species and organs implied that genes involved in wax biosynthesis might also be different. These results provide basic knowledges in studying the molecular mechanisms of cuticular waxes in legume green manure crops fronting biotic and abiotic stresses.
Cuticular wax; green manures;;;
2019-03-24;
2019-05-12;
2019-07-22.
10.3724/SP.J.1006.2020.94048
郭彦军, E-mail: qhgyj@126.com
E-mail: 286923599@qq.com
本研究由国家自然科学基金项目(31670407)和中央高校基本科研业务费(XDJK2018B021)资助。
This study was supported by the National Natural Science Foundation of China (31670407) and the Fundamental Research Funds for the Central Universities (XDJK2018B021).
URL: http://kns.cnki.net/kcms/detail/11.1809.S.20190719.1721.008.html
猜你喜欢
纺织标准与质量(2022年3期)2022-08-10
核农学报(2022年6期)2022-05-20
辽宁农业科学(2021年6期)2021-12-02
科学与财富(2021年31期)2021-03-08
作文周刊·小学二年级版(2021年40期)2021-01-05
伴侣(2020年4期)2020-04-27
伴侣(2020年2期)2020-04-13
干旱地区农业研究(2020年6期)2020-02-03
健康必读(2016年10期)2016-11-14
数理化学习·高一二版(2009年7期)2009-11-23
