
鳜·斑鳜及杂交鳜鳜♀×斑鳜♂的染色体组型研究
2020-12-11 09:18吕青兰薛源史杨白
安徽农业科学 2020年21期
吕青兰 薛源 史杨白
摘要 鳜鱼(Siniperca chuatsi)、斑鳜(Siniperca scherzeri)及其杂交鳜(鳜♀×斑鳜♂)为研究对象,采用植物血球凝集素(PHA)和秋水仙素体内注射获得大量有丝分裂中期相的头肾细胞,直接制片法制备染色体标本后进行染色体组型的研究和分析。结果表明:3种鳜鱼染色体数目皆为2n=48,均未观察到随体、次缢痕或其他特殊标志的染色体。鳜、斑鳜和杂交鳜的核型公式分别为2n=8sm+10st+30t(NF=56)、2n=2m+8sm+10st+28t(NF=58)和2n=2sm+12st+34t(NF=50)。该研究鳜和斑鳜的染色体组型与其他水系种群相比有所差异,呈现多态性。首次报道的杂交鳜染色体组型与父母本皆有不同,显示了杂交后代复杂的染色体变异。
关键词 鳜;斑鳜;杂交鳜;染色体组型
中图分类号 S917.4 文献标识码 A
文章编号 0517-6611(2020)21-0095-05
Abstract Taking Siniperca chuatsi,Siniperca scherzeri and their hybrids[S.chuatsi(♀)×S.scherzeri(♂)]as the research objects,a large number of head kidney cells in mitotic metaphase were obtained by PHA and colchicine injection in vivo,and then the chromosome samples were prepared by direct production method,the karyotypes were studied and analyzed.The results showed that the chromosome number of three species of mandarin fish were all 2n = 48,and no satellite,secondary constriction or other special markers were observed.The karyotype formulae of S.chuatsi,S.scherzeri and their hybrids were 2n=8sm+10st+30t(NF=56),2n=2m+8sm+10st+28t(NF=58) and 2n=2sm+12st+34t(NF=50),respectively.In this experiment,the karyotypes of S.chuatsi and S.scherzeri were different from those of other river populations,showing polymorphism.The karyotype of hybrid mandarin fish reported for the first time was different from that of its parents,which showed the chromosomal intricately mutated in hybrid offspring.
Key words S.chuuatsi;S.scherzeri;Hybrid mandarin fish;Karyotype
基金项目 江苏省重点研发计划(现代农业)项目(BE2017311,BE2018398);苏州大学校级大学生创新创业训练计划项目(2019xj069);江苏省渔业科技项目(cx(18)2012-01)。
作者简介 吕青兰(1997—),女,江苏盐城人,从事鱼类生物学研究。*通信作者,副教授,博士,从事水生生物学研究。
收稿日期 2020-05-06
鳜(Siniperca chuatsi)(也称鳜鱼)和斑鳜(Siniperca scherzeri)在分类学上均隶属于鲈形目 (Perciformes)真鲈科(Percichthyidae)鳜亚科 (Siniperinae)鳜属(Siniperca),广泛分布于我国除青藏高原外的其他水系[1-2]。鳜生长速度快、味鲜肉嫩、营养价值高,但抗逆性较低,以活体小型鱼虾类为食。斑鳜营养味美,被誉为“淡水石斑”[3],具有较强的抗病力,并能摄食冰鲜鱼虾,但生长速度较慢[1,3]。杂交鳜兼备双亲生长快、抗病力强、饲养简单等优良性状[3-4],且营养价值较双亲高[5]。
染色体组型(也称染色体核型,karyotype),一般是染色体数目、大小、形态、着丝点位置、随体、次缢痕以及带型(G带、C 带等)[6]所有染色体可测性状的总称。目前,已有对鄱阳湖鳜鱼[7]、浙江乌溪江水库斑鳜[8]等染色体组型的研究报道。研究表明,斑鳜自然群体遗传多样性较高,且因区域、水系的不同而存在明显的遗传分化现象,例如福建与广西的斑鱖存在遗传差异;鳜鱼群体遗传多样性略低于斑鳜,但南北方鳜鱼的遗传差异也比较明显[2,9]。笔者首次分别研究来自于长江水系鳜和黑龙江水系斑鳜及其杂交鳜(鳜♀×斑鳜♂)的染色体组型,以期较全面地掌握不同地域鳜鱼、斑鳜的染色体组型特征,并了解种间杂交后代染色体组型的变异情况,进而为鱼类杂交新品种的选育、种质鉴定等方面提供遗传学基础资料。
1 材料与方法
1.1 材料 试验用鱼均取自江苏省淡水水产研究所常州水产良种繁养基地,鳜来自于长江水域,体长20~30 cm,体重360~400 g;斑鳜来自于黑龙江水域,体长15~20 cm,体重50~100 g;杂交鳜(鳜♀×斑鳜♂)是以上述2种鱼为亲本的杂交后代,体长22~26 cm,体重200~300 g。每种鱼均随机取样4尾。水族箱内恒温(23 ℃)、充氧暂养12 d后用于试验。
1.2 方法
1.2.1 染色体玻片的制备。
采用林义浩[10]的体内注射植物血球凝集素 (PHA) 、活体肾细胞直接制染色体玻片的方法,并参照朱健等[11]、Lu等[12]、刘良国等[13]的方法加以改进。分2次腹腔注射PHA溶液,第一次注射剂量为10 μg/g,继续暂养15 h后进行第2次注射,剂量为8 μg/g;5 h后按3 μg/g的剂量注射秋水仙素。2.5 h后,剪断鱼尾、腮丝血管放血15 min,然后取头肾组织于0.75%生理盐水中破碎组织,用4层纱布将细胞悬液过滤入50 mL离心管中,加入适量生理盐水,吹打均匀(100次以上),制成头肾细胞提取液,2 000 r/min离心8 min,弃上层清液;用 8 mL预热好的0.075 mol/L KCl低渗液吹打2次,置于37 ℃水浴中处理30 min;加入1 mL固定液预固定1 min;以1 500 r/min转速离心10 min,弃上清液;加9 mL固定液在37 ℃水浴中固定30 min;以1 500 r/min离心10 min,弃上清液;重复上述步骤固定3次;最后,弃去大部分上清液,留约1 mL制成细胞悬液,用滴管轻轻吹打均匀;用吸管吸取细胞悬液并从约1 m高处滴在经预冷、倾斜30°的洁净玻片上,轻吹玻片上的细胞悬液使之扩散;自然干燥后用10% Giemsa染色液染色10~15 min;蒸馏水冲洗干净,空气中自然干燥;显微镜观察染色体形态。
1.2.2 染色体组型分析。
显微镜观察后,选取50个分散良好、形态清晰的染色体中期分裂相进行染色体计数,统计染色体众数;选取状态良好的中期分裂相的二倍体染色体,在油镜下进行显微摄影。选取3个染色体数目完整、形态清晰、分散良好的中期分裂相原始图,放大并编号,并依据形态、大小等特征进行配对;使用Image J 1.8.0软件将照片放大,進行染色体配对和数据测量,计算染色体相对长度[染色体相对长度=(每条染色体单体长度/染色体组总长度)×100%]和臂比(臂比=长臂长/短臂长),按照Levan等[14]的命名和分类方法(表1),对每对染色体进行分类、排列,锐化处理后获得染色体组型图。
2 结果与分析
2.1 染色体数目
油镜下分别统计鳜、斑鳜和杂交鳜染色体中期分裂相各50个。由表2可知,鳜、斑鳜和杂交鳜的染色体众数分布率分别为86%、84%和78%,因而可确定鳜、斑鳜、杂交鳜这3种鳜鱼的二倍体染色体数目皆为48。
2.2 染色体组型分析
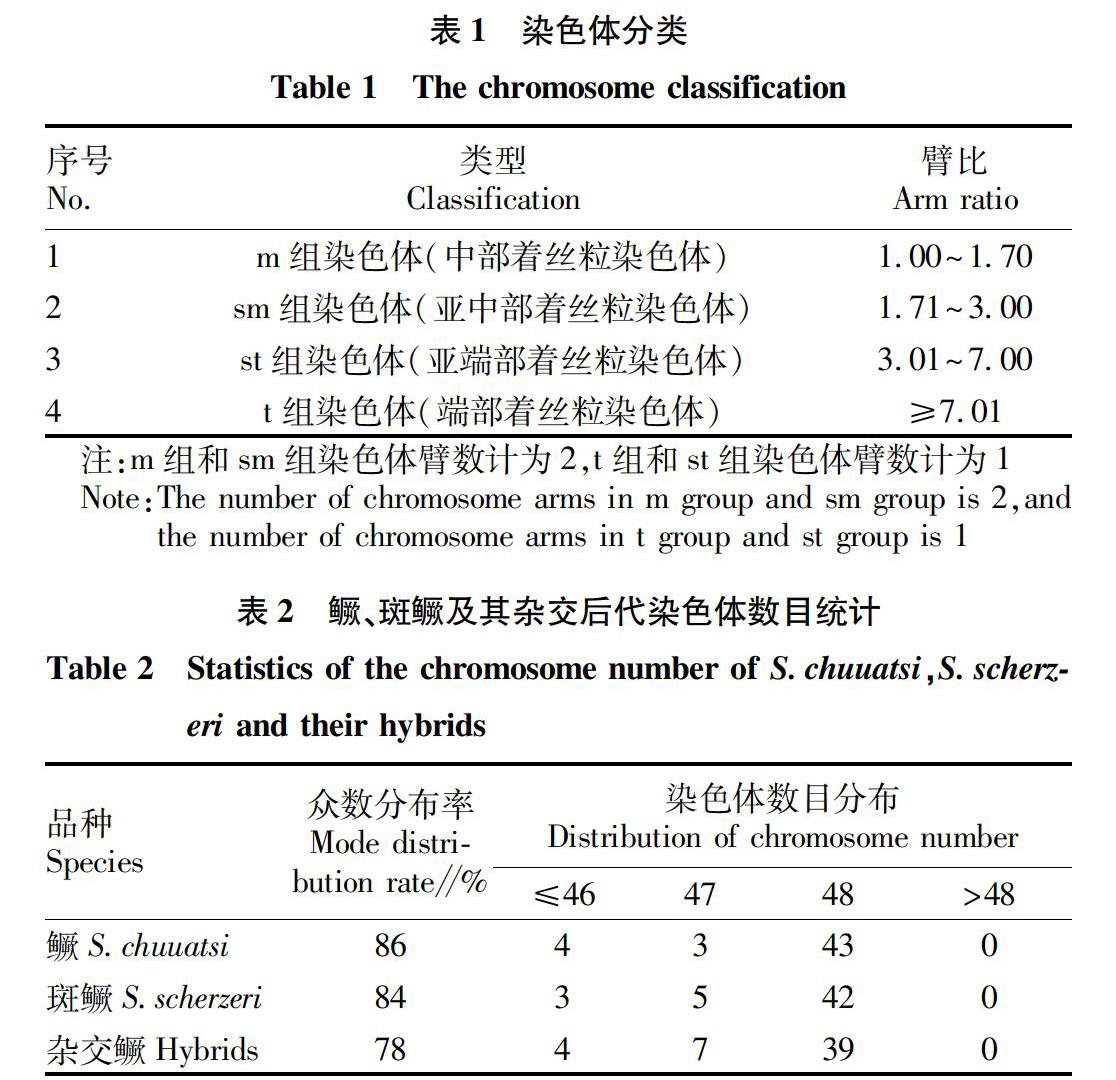
2.2.1 鳜染色体组型。
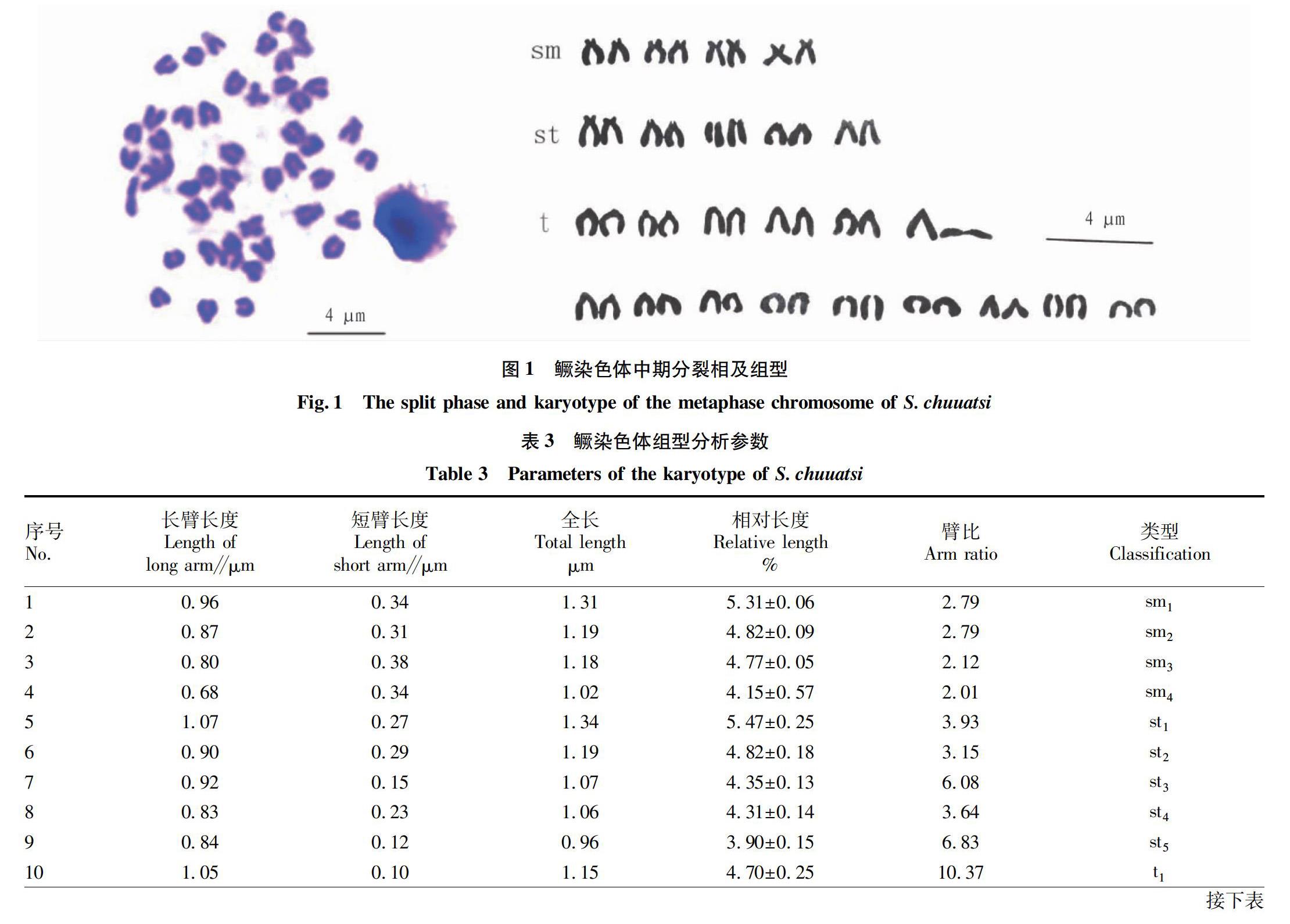
根据鳜染色体大小和形态,结合臂比和相对长度进行同源染色体配对,共配成24对染色体(图1,表3)。按照Levan命名法,鳜的染色体可分为3组,其中4对亚中部着丝粒染色体(sm组)、5对亚端部着丝粒染色体(st组)和15对端部着丝粒染色体(t组)(图1,表3),其核型公式为2n=48=8sm+10st+30t,染色体臂数(NF)为56。
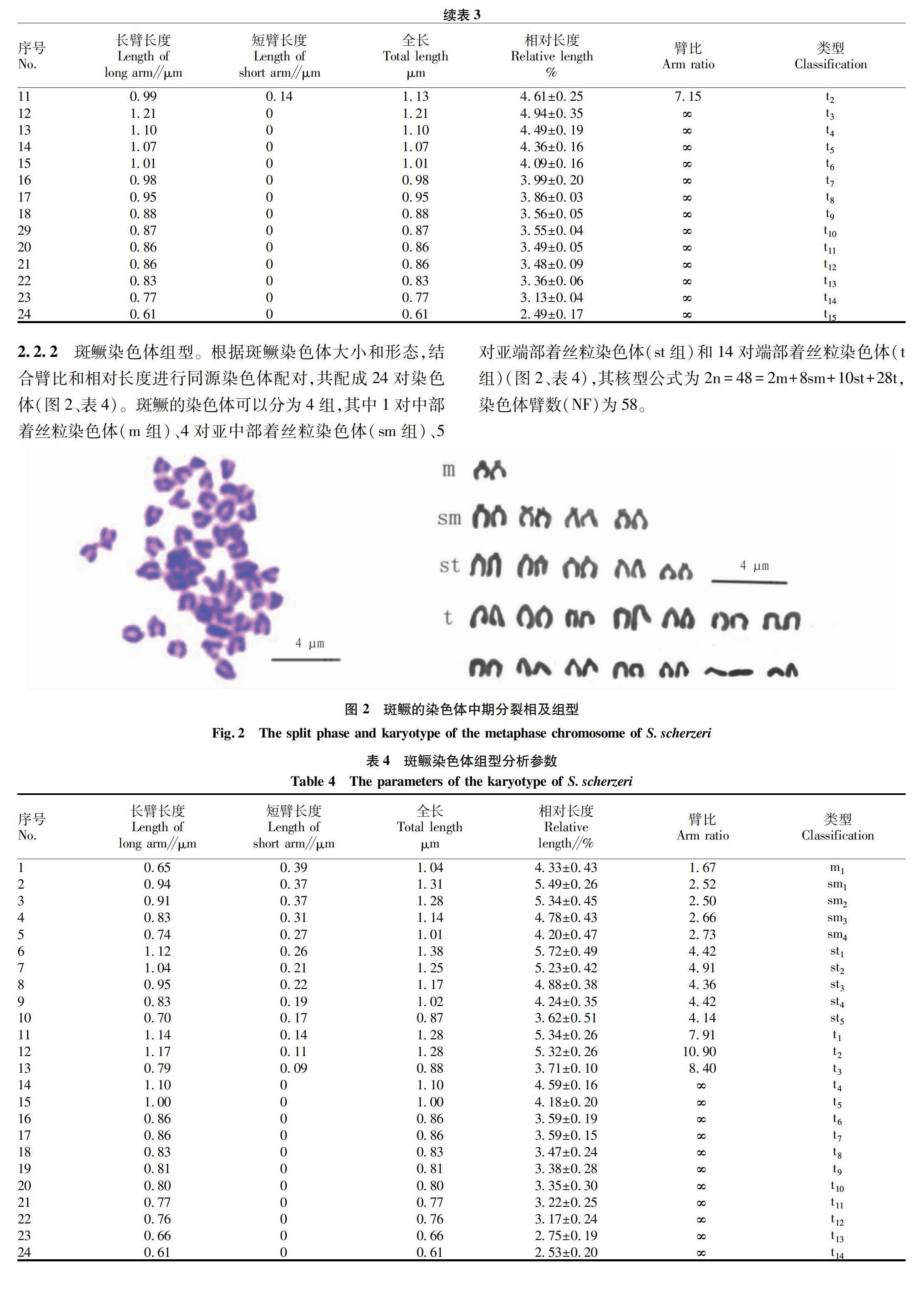
2.2.2 斑鳜染色体组型。
根据斑鳜染色体大小和形态,结合臂比和相对长度进行同源染色体配对,共配成24对染色体(图2、表4)。斑鳜的染色体可以分为4组,其中1对中部着丝粒染色体(m组)、4对亚中部着丝粒染色体(sm组)、5对亚端部着丝粒染色体(st组)和14对端部着丝粒染色体(t组)(图2、表4),其核型公式为2n=48=2m+8sm+10st+28t,染色体臂数(NF)为58。
2.2.3 杂交鳜染色体组型。
根据杂交鳜染色体大小、形态,结合臂比和相对长度进行同源染色体配对,共配成24对染色体(图3、表5)。杂交鳜的染色体可以分为3组,其中1对亚中部着丝粒染色体(sm组)、6对亚端部着丝粒染色体(st组)和17对端部着丝粒染色体(t组)(图3、表5),其核型公式为2n=48=2sm+12st+34t,染色体臂数(NF)为50。
3 讨论
3.1 鳜、斑鳜染色体组型特征
鳜染色体组型在国内外已有不少报道[7,11,15-18]。该研究结果显示,鳜染色体众数为2n=48,核型公式为2n=8sm+10st+30t(NF=54)。染色体数目、染色体配对与臂数的结果与已有报道的结论是一致或相似的,但在着丝点位置的确定和染色体分组上存在一定的差异。该研究的长江水系鳜核型公式与朱健等[11]报道的苏州太湖鳜(同属长江水系)染色体组型2n=48(6sm+12st+30t,NF=56)相似,但与杨春等[7]报道的鄱阳湖鳜染色体组型2n=48(24sm,st+ 24t,NF=72)差异较大,存在着sm 染色体减少的现象。这可能是由于鳜的地理差异引起的遗传分化现象,但还有待采用分子遗传学等方法做进一步验证。其中臂数的巨大差异可能是由于与杨春等[7]报道的st组染色体臂数确定标准不一样导致的。该研究结果表明黑龙江水系斑鳜染色体组型为2n=48(2m+8sm+10st+28t,NF=58),与张晓辉等[8]报道的浙江乌溪江水库斑鳜染色体组型2n=48(4m+10sm+4st+30t,NF=62)相似,但与余先觉等[17]报道的斑鳜染色体组型2n=48(6sm+14st+28t,NF=54)相比存在m染色体增多的现象,这一较大的差异是否表明斑鳜存在亚种或地理种群有待进一步研究。李树深[19]指出,在特定的分类阶元中,具有较多中部着丝粒染色体(m)和亚中部着丝粒染色体(sm)为较特化类群,具有较多端部着丝粒染色体(t)为较原始类群,因此可认为鳜、斑鳜均属于较特化类群。通过该研究的大量试验以及查阅相关研究报道后发现,造成同种鱼类染色体组型特征差异的原因是多方面的,主要包括:①观察时间点差异。若染色体分裂相不处于中期,而是偏向染色体缩短的中后期时,就很容易把亚端部和端部着丝粒染色体错误分类,因此必须采用多时间点、多样本采样加以纠正。②不同研究者采用试验方法、滴片方法、测量手段、确定着丝点的位置等方法的不同都可能造成实验数据的差异。因此,需要制定统一标准,以便用于科学的种质分类。③地域环境差异。鱼类在演化过程因地域环境差异,可能发生了染色体臂间易位、倒位等结构重排[20],从而引起染色体多态性。因此,需要利用更多的生物学方法进行深入研究。该研究结果显示,鳜和斑鳜的染色体组型公式较为相似,因此杂交亲和性高,不存在生殖隔离,容易获得正常发育的胚胎,并能培育出发育良好的杂交鳜。已有杂交鳜培育成功的报道[3-4,21]也证实了这一假设。
3.2 杂交鳜与其双亲染色体组型特征的比较 笔者首次报道了杂交鳜的染色体组型特征。结果显示,染色体数目与双亲相同,且短臂上均未见随体、次缢痕;但缺少父本斑鳜的中部着丝粒染色体,且比双亲少了3对亚中部着丝粒染色体;而比双亲多了1对亚端部着丝粒染色体,且分别比母本鳜和父本斑鳜分别多了2对和3对端部着丝粒染色体。这表明,杂交鳜的染色体组型是由2个亲本的染色体经过较复杂的遗传重组后形成的。从杂交鳜的表型特征来看,除体型和生长速度更接近母本鳜鱼外,其抗病力强、肉质细嫩、体色花纹等均较接近父本斑鳜[3]。这表明杂交鳜兼具双亲的遗传性状,显示出良好的杂交优势。但是,关于杂交鳜与双亲更深入的亲缘关系还有待进一步研究。
3.3 鱼类有丝分裂中期分裂相的获取
与哺乳动物相比,鱼类有丝分裂中期染色体具有形态小、数量多、分裂少等特点。因此,制备具有大量、形态清晰、染色体数目完整的中期分裂相玻片具有较大难度。在该试验过程中发现,将饲养水温等环境条件控制在试验鱼的最适生理状态下,并采用2次注射PHA的方法,可获得较多理想的中期分裂相;此外,PHA和秋水仙素的作用剂量和时间也对染色体制片效果产生影响。据有关文献记载[22],PHA剂量在8~10 μg/g是安全的,超过20 μg/g可能会导致某些鱼类死亡;秋水仙素通常的作用剂量范围为1~5 μg/g,作用时间为2~3 h,若用量过大或处理时间过长,会导致染色体的过分凝缩或着丝点裂解,最终导致染色体形态不正常,甚至被破坏或溶解;浓度过低或处理时间过长,则分裂相过少[22]。该研究进行了预固定操作,因为预固定有利于细胞提前适应溶液的酸性环境,预先固定染色体形态,有利于获得理想的染色体分裂相[23]。冰乙酸的纯度以及甲醇与冰乙酸的配制比例对分裂相的获得也有很大的影响。该研究所用固定液为甲醇∶冰乙酸(分析纯)按1∶2(V/V)配比混合;若冰乙酸是优级纯,则甲醇∶冰乙酸应按3∶1(V/V)配制才可以获得较多的分裂相[23]。另外,选取合适的组织是制备染色体标本的前提,该研究优选了生命活动旺盛、分裂增殖较快的头肾组织,取得了较满意的效果。
参考文献
[1]陈文军,何贤臣,尹海富.鳜的生物学特性和流行病学初探[J].黑龙江水产,2018(2):31-33.
[2]宋书莉,赵金良,李晨虹.鳜属鱼类的进化和遗传多样性研究[C]//中国水产学会.2017年中国水产学会学术年会论文摘要集.北京:中国水产学会,2017:1.
[3]郝忱,丁淑燕,刘国兴,等.杂交鳜人工繁殖与苗种培育技术初探[J].水产养殖,2019,40(3):1-2.
[4]王德忠,朱珠,张学师,等.翘嘴鳜(♀)×斑鳜(♂)的人工繁殖及苗种培育试验[J].科学养鱼,2019(2):8-10.
[5]李燕,史建华,李永强,等.翘嘴鳜、斑鳜和杂交鳜鱼体营养成分和氨基酸、脂肪酸组成的比较分析[J].水产科技情报,2015,42(5):246-250.
[6]牛文涛,蔡泽平.中国海水鱼类核型研究概述[J].厦门大学学报(自然科学版),2006,45(S2):125-131.
[7]杨春,李达,徐光龙,等.鄱阳湖鳜鱼染色体组型的研究[J].江西农业学报,1999,11(1):52-55.
[8]张晓辉,朱扬玲,杜建明,等.斑鳜染色体核型研究[J].浙江农业科学,2007(2):226-227.
[9]李传阳,许淼洋,赵金良,等.斑鳜(♀)×鳜鱼(♂)杂交后代遗传特征的微卫星分析[J].水产科学,2014,33(2):97-102.
[10]林義浩.快速获得大量鱼类肾细胞中期分裂相的PHA体内注射法[J].水产学报,1982,6(3):201-208.
[11]朱健,张成锋,闵宽洪,等.尼罗尖吻鲈和鳜鱼染色体组型分析及比较[J].水生生物学报,2009,33(2):195-199.
[12]LU H,MENG Q L,YANG L.Karyotype analysis of Cyprinus carpio in Dongping Lake[J].Animal husbandry and feed science,2017,9(6):394-397.
[13]刘良国,邹万生,杨春英,等.洞庭湖水系黄颡鱼的形态差异及染色体组型(英文)[J].Agricultural science & technology,2011,12(10):1521-1524.
[14]LEVAN A,FREDGA K,SANDBERG A A.Nomenclature for centromeric position on chromosomes[J].Heresitas,1964,52(2):201-220.
[15]杨慧一.鱖鱼(Siniperca chuatsi)染色体组型的研究[J].遗传学报,1982,9(2):143-146.
[16]凌均秀.八种鱼的染色体组型的研究[J].武汉大学学报(自然科学版),1982(2):109-112.
[17]余先觉,周敦,李渝成,等.中国淡水鱼类染色体[M].北京:科学出版社,1989.
[18]曹丽萍,丁炜东,贾永义,等.奥利亚罗非鱼(♀)×鳜(♂)杂交后代及其双亲的染色体核型分析[J].农业生物技术学报,2006,14(2):187-190.
[19]李树深.鱼类细胞分类学[J].生物科学动态,1981(2):8-15.
[20]李康,桂建芳,洪云汉,等.中国鲤科鱼类染色体组型的研究Ⅴ.鮈亚科10种鱼的染色体组型[J].武汉大学学报(自然科学版),1984,30(3):113-122.
[21]古丽.长珠杂交鳜[J].农村百事通,2018(3):29.
[22]黎玉元,孙念,李伟,等.鱼类染色体制备方法概述[J].湖南饲料,2013(4):28-30.
[23]杨秋月,赵俊鹏,王玉君,等.预固定在淋巴细胞染色体制备中的量化分析[J].医学动物防制,2019,35(5):509-512.
