
水稻成蘖特性及相关氮同化酶活性、基因表达分析
2020-12-30 06:20季雯珺何永刚章志宏
湖北农业科学 2020年23期
安 琪,季雯珺,何永刚,章志宏
(武汉大学生命科学学院/杂交水稻国家重点实验室(武汉大学),武汉 430072)
水稻(Oryza sativa L.)是世界上最重要的粮食作物之一,养活了全球一半以上的人口。水稻产量的构成因素包括单位面积有效穗数、每穗颖花数、千粒重和结实率等[1]。而水稻分蘖直接决定有效穗数,并对每穗颖花数、结实率和千粒重有重要影响。
水稻分蘖的形成过程主要包括分蘖芽的形成与分蘖芽的伸长[2-4]。在每个叶位的叶腋里一般都能形成一个腋芽,也叫分蘖芽,分蘖芽能够进一步伸长生长形成分蘖。影响水稻成芽、成蘖的因素包括环境因素和遗传因素。环境因素包括光、温、水、肥等[5,6]。有研究表明,水稻分蘖芽在三幼一基期对环境最为敏感[7]。氮素是植物生长所必需的大量元素之一,也是决定植物生物量和作物产量的主要限制因素[8,9]。在水稻营养生长期,充足的氮素供应可以促进分蘖[10],氮供应不足会导致有效分蘖数大量减少[11]。遗传因素包括调控腋芽形成和调节分蘖芽伸长的基因、植物激素、碳/氮同化相关基因等。影响腋生分生组织分化形成腋芽的基因有TE/TAD1、MOC1、MOC3、LAX1 等。 水 稻TILLERS ABSENT1(TAB1)与拟南芥WUSCHEL(WUS)基因同源,参与腋生分生组织的发育起始[12]。MOC1(monoculm1)是第一个被确认为控制水稻分蘖的基因,该基因突变体的叶腋处不能形成分蘖芽,在成熟期只有一个主茎秆[2]。单蘖突变体MOC3(monoculm3),由于一个点突变而导致TAB1 基因的提前终止,从而妨碍了分蘖芽的形成。TE/TAD1 通过调控MOC1 的降解来调控水稻分蘖。TAD1 编码含有细胞分裂后期促进复合体/细胞周期体(APC/C)和多亚基E3 连接酶的一个共激活蛋白,同APC/C 复合体的亚基CDC27 直接相互作用[13,14]。TE 与MOC1 在叶腋中共表达,真核生物中功能高度保守的E3 泛素连接酶APC/C 与TE 形成复合体APC/CTE,通过26S 蛋白酶体泛素化降解MOC1[13,14],从而使水稻分生组织特异性基因homeobox1 的表达下调,抑制腋生分生组织的起始与形成[13]。APC/CTE能够保守性地降解细胞周期中的关键调控因子,从而特异性地调控分蘖发生,影响水稻株型和子粒产量[13,15]。
SLs 信号通路在植物分蘖形成中具有重要的调控作用,其中涉及到的基因包括SLs 合成途径的基因D27、D17、D10 和SL 信号传导的基因D14、D3、D53。如水稻的D10(DWARF10)与拟南芥的MAX4是直系同源基因,编码一个类胡萝卜素裂解双加氧酶,参与SL/SLs 的生物合成,控制水稻侧芽向外生长[16]。 HTD1(HIGH-TILLERING DWARF1)/D17 编码水稻中类胡萝卜素裂解双加氧酶OsCCD7,在类胡萝卜素衍生物的合成中发挥作用,负调控水稻侧芽生长[17]。突变体d27 中生长素的极性运输能力增强,表现出分蘖增加和株高变矮的表型。D3 通过控制分蘖芽的休眠性从而抑制分蘖芽的活性。d3突变体中有观察到对于下胚轴的生长缺陷,因此D3可能调控多个发育阶段中不同蛋白质的降解过程,也对分蘖阶段进行调控[18]。HTD2/D14/D88 编码一个α/β 折叠水解酶超家族的蛋白,抑制水稻分蘖的发生,负调控水稻分蘖数[19-21]。HTD2 的突变不影响分蘖芽的形成,但对随后分蘖芽的向外伸长生长有影响,导致分蘖增加、植株矮化的表型[19]。IPA1/OsSPL14 作为SPL 基因家族的一员,起着转录因子的作用,在营养生长阶段,IPA1 控制水稻分蘖;在生殖生长阶段,IPA1 的高表达促进了穗分枝。IPA1 能与一些基因的启动子直接结合,如直接正向调控水稻株型的重要基因DEP1 以及负向调控水稻分蘖芽生长的关键基因OsTB1,进而分别抑制水稻分蘖发生和调节水稻穗长和株高[22]。
调控水稻分蘖的激素包括生长素、细胞分裂素和独角金内酯等。生长素和独角金内酯抑制分蘖芽的生长发育[23,24],而细胞分裂素则促进分蘖芽的生长[25]。解释植物激素调控分蘖发生的2 个模型[26],一个是生长素第二信使模型:IAA 通过正调控SL、负调控CK 起作用,而SL 和CK 进一步调控蘖芽伸长的基因从而调控蘖芽生长。另一个是生长素运输通道模型:IAA 运输通道被SL 所抑制,SL 会抑制IAA 运输蛋白PIN,从而抑制IAA 从分蘖芽中输出,进而抑制腋芽的生长[26]。
水稻育种和生产实践表明,低节位、早发分蘖在拔节期具有与主茎媲美的生物量,在成熟期穗部性状上更接近主茎,对植株产量的形成具有更大贡献。目前专注于水稻低节位、早发分蘖的深入研究很少。本研究在考察水稻品种低节位、早发分蘖遗传变异的基础上,较深入地探究了水稻低节位、早发分蘖的分子遗传基础。
1 材料与方法
1.1 试验材料及种植
选用具有不同分蘖特性的水稻品种天优华占、岳4B、明恢63、扬两优6 号、Y 两优800、中嘉早17、黄 华占、HR901、鄂早18、Basmatis、Lemont 和Star Bonnet 为材料,采用水培法培养水稻秧苗,营养液配方为改良后的国际水稻研究所(IRRI)常规营养液配方[27]。水稻种子浸种10 h、沥水2 h,再用强氯精2 g 对水800 mL,浸种10~12 h,后用清水冲洗多次,于8 月18 日播种于潮湿的蛭石中,室外遮阴待种子发芽。第1 叶展开后,移至自然条件下光照培养,用1/4 营养液浇灌,2 叶开始用1/2 营养液。8月27 日选取2 叶1 心的水稻秧苗移栽到水培泡沫盘中,每穴1 苗,株、行距为6 cm×9 cm,每个品种在2 个水培盆(60 cm×36 cm×12 cm)中种植42 株。移栽后用全营养液培养,每隔2 d 更换一次营养液,每个培养盆用营养液10 L;5~6 叶期,每盆用营养液15 L。每天用HCl 或NaOH 调节营养液的pH 为5.0~5.5,并适当补充挥发掉的水份。各叶龄期间隔取样,以保证每个水稻品种相对生长环境一致。
1.2 水稻秧苗分蘖特性及相关农艺性状的观测
在水稻秧苗培养过程中,每天观察记录分蘖发生情况。新的分蘖发生,以该分蘖的第1 片叶伸出着生叶鞘1 cm 为记录标准。
每个水稻品种3 叶1 心至5 叶1 心时期,每个叶龄取样5 株,测定各叶龄时期的苗高、根长、各叶片的叶长、叶宽、SPAD 值。随后,对取样材料用105 ℃杀青10 h 左右,80 ℃烘干至恒重,测定各器官及全株的干物重。
1.3 水稻秧苗生理生化指标的测定
晴天上午10:00—11:00,每个水培盆随机取3~4 叶期水稻秧苗5 株,选择完全展开叶,将叶片放入液氮中保存,用于酶活性检测;叶鞘置于牛皮纸袋中105 ℃杀青,80 ℃烘干至恒重,用于可溶性糖含量测定。
粗酶液提取及保存参考卢永恩[28]的方法,谷氨酰胺合成酶(GS)活性检测参考Rhodes 等[29]的方法,NADH-GOGAT(谷氨酸合酶)活性测定参照Leyser[25]的方法进行。NADH-GDH(谷氨酸脱氢酶)活性测定参照Turano 等[30]的方法进行。可溶性糖的测定参考张志良等[31]的《植物生理学实验指导》。
1.4 水稻秧苗基因表达的测定
晴天上午10:00—11:00 取3~4 叶期水稻秧苗,用于铵态氮同化相关酶合成基因、生长素和细胞分裂素合成基因的表达检测;每个水培池随机取5株,取分蘖节和芽放入液氮中保存,用于分蘖相关基因的表达检测。
RNA 提取采用TRIzol(Invitrogen 公司)提取法;DNA 消化用DNaseⅠ购自Thermo 公司;反转按照THUNDERBIRD SYBR®qPCR Mix QPS-201 说明书操作,试剂购自上海硕盟生物科技有限公司。
1.5 数据处理
利用Excel 软件记录整理数据,采用SPSS 20.0软件进行相关性分析。
2 结果与分析
2.1 水稻秧苗分蘖发生动态
从水稻秧苗在播种后18~34 d 的分蘖发生动态(表1)来看,12 个供试品种的分蘖特性具有较大变异。其中,天优华占、扬两优6 号、岳4B 和明恢63 4 个品种,在播种后18 d 时单株分蘖数达0.9~1.3个,随后一直保持较高的单株分蘖数,属于分蘖早发类型的品种。Y 两优800、中嘉早17、黄华占和Basmatis 4 个品种,在播种后18 d 时的单株分蘖数为0~0.2 个,但随后快速增长,单株分蘖数仅次于上述分蘖早发类型的品种,属于蘖性居中类型的品种。HR901、鄂早18、Lemont 和Star Bonnet 4 个品种在播种后18 d 时均未发生分蘖,播种后22 d 也只有HR901 的单株分蘖数为0.1 个,此后的分蘖增长一直较慢,这几个品种属于分蘖迟发类型的品种。从表1 显著性分析也可以看出,播种后18~26 d,分蘖早发类型的4 个水稻品种分蘖数与其余8 个水稻品种一直都有显著性差异。播种后22~30 d,分蘖早发类型、蘖性居中类型的水稻品种与分蘖迟发类型的水稻品种间平均分蘖数一直都有显著性差异。

表1 水稻不同品种苗期单株分蘖数
2.2 不同水稻品种苗期第1~3 叶位成蘖率
为研究水稻早发分蘖的发生情况,本研究观察统计了上述12 个水稻品种第1~3 叶位的成蘖率。统计结果如图1 所示,12 个水稻品种中只有天优华占、扬两优6 号、岳4B、Y 两优800 在第1 叶位上长出分蘖,其成蘖率分别为80%、50%、20%、10%。第2 叶位除黄华占、HR901 和鄂早18 成蘖率为0 外,其余水稻品种都形成分蘖且成蘖率都在30% 以上。天优华占、扬两优6 号、岳4B、明恢63 和Lemont 第2 叶位成蘖率为60%~100%。在第3 叶位上仅有鄂早18 的成蘖率是10%,其余水稻品种的成蘖率都在60% 以上,其中天优华占、扬两优6 号、岳4B 和明恢63 的成蘖率达100%。
2.3 水稻第1~3叶位成蘖率与分蘖数的相关性分析
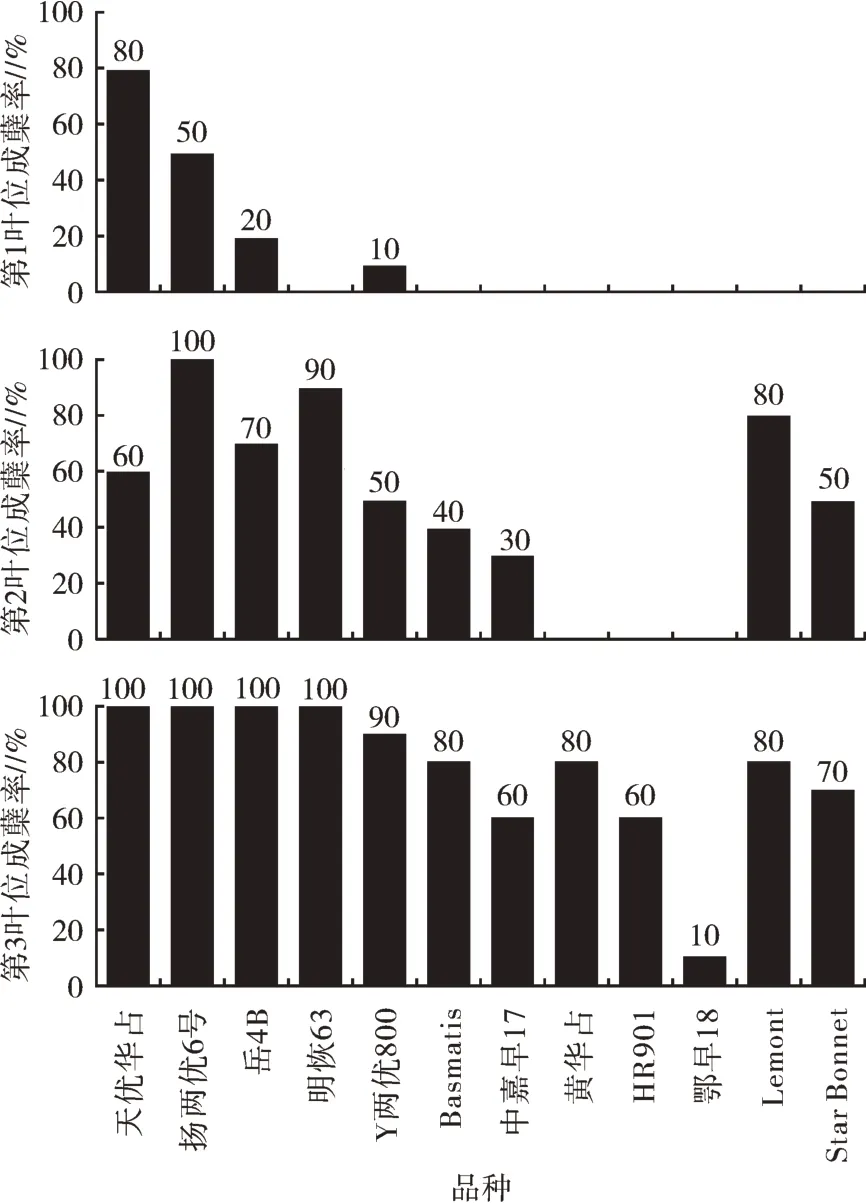
图1 水稻不同品种苗期第1~3 叶位成蘖率
为分析分蘖数与第1~3 叶位成蘖率之间的关系,本研究将12 个水稻品种1~3 叶位成蘖率分别与单株分蘖数进行了相关性分析(表2),由表2 可知,水稻第2、3 叶位成蘖率与分蘖数呈显著正相关,特别第2 叶位成蘖率与分蘖数之间的正相关性达极显著水平(P<0.01),且相关系数高达0.754~0.931。水稻播种后18~34 d 的单株分蘖数与第1 叶位成蘖率之间相关性不显著,说明第1 叶位成蘖率对单株分蘖数的影响不大。
2.4 水稻第1~3 叶位成蘖率与苗期叶片铵态氮同化酶活性、叶鞘可溶性糖含量的相关性分析
氮素水平是影响水稻分蘖发生的重要因素之一,充足的氮素供应可以促进分蘖[10],供应不足会导致有效分蘖数大量减少[11]。在生长发育过程中水稻植株从稻田吸收无机氮的主要形式为NH4+,氮素同化酶GS、GOGAT 和GDH 在将无机氮NH4+转化为有机氮的过程中起着关键作用。由表3 可知,第1 叶位成蘖率与GS、NADH-GOGAT 和NADHGDH 的活性均呈显著正相关,而第2 叶位成蘖率只与NADH-GDH 酶活性呈极显著正相关(P<0.01),第3 叶位成蘖率只与NADH-GOGAT 酶活性呈显著正相关(P<0.05)。此外,表2 结果还显示,水稻第1~3 叶位成蘖率均与秧苗叶鞘的可溶性糖含量之间的相关性不显著。

表2 水稻第1~3 叶位成蘖率与单株分蘖数的相关性分析
2.5 水稻第1~3 叶位成蘖率与分蘖相关基因表达量的相关性分析
由表4 可知,水稻秧苗第1 叶位成蘖率与分蘖芽发育相关基因(MOC1 和OGR1)、IAA 合成相关基因YUCCA6、CK 合成基因IPT5 以及SL 合成及调控通路相关基因(HTD1、D10、D3、D14、IPA1 和TB1)的表达量之间的相关性均不显著,表明水稻第1 叶位成蘖率基本不受水稻分蘖芽发育和分蘖形成的遗传调控系统的调控。第2 和第3 叶位的成蘖率均与分蘖芽发育相关基因(MOC1 和OGR1)和CK 合成基因IPT5 的表达量之间相关性不显著,但分别与IAA 合成相关基因YUCCA6 和SL 合成及调控通路相关的多数基因(HTD1、D10、D3、D14、IPA1 和TB1)的表达量呈显著或极显著负相关,表明水稻第2 和第3 叶位成蘖率不受分蘖芽发育的制约,IAA 合成和SL 合成及调控是负调控水稻第2 和第3 叶位成蘖率的主要遗传基础。

表3 水稻秧苗第1~3 叶位成蘖率与苗期叶片铵态氮同化酶活性、叶鞘可溶性糖含量的相关性分析

表4 水稻第1~3 叶位成蘖率与秧苗分蘖节分蘖相关基因表达量的相关性分析
3 小结与讨论
水稻分蘖是影响水稻产量的重要农艺性状之一,受水稻植株遗传基础及其生长发育环境的共同调控。不论是“超级杂交稻”“超高产新株型”还是“水稻理想株型”,其核心内容都是控制水稻的无效分蘖、提高成穗率,进而提高水稻产量。成穗数是水稻产量的重要构成之一,而单株有效分蘖数又是决定成穗数的重要因素,过低或过高的分蘖数都会影响成穗数和成穗率,进而影响到最终的产量形成[32,33]。在水稻生产中,人们习惯上将成穗率高和穗形较大的分蘖叫做优势分蘖[34]。优势的强弱一般是一级分蘖>二级分蘖>三级分蘖,同是一级分蘖的,其所处的节位比较低或者出生时间比较早,常常有较高的成穗率,穗形也比较大,因此生产上强调栽后早发分蘖。黄耀祥等[35]提出了水稻的“早发”概念,即早在营养生长前期就长出较为强壮的分蘖,为重穗型遗传特性的表达和孕育大穗提供了有效保证。蒋彭炎等[36]的“三高一稳栽培法”中也提出要促进早期分蘖的发生。在水稻生产中,低节位(早发)分蘖往往能够形成有效分蘖,在成熟期其穗部性状更接近主茎,对植株产量的形成具有重要贡献。在小麦中,同样也发现低节位分蘖穗的每穗粒数和每穗可育小穗数更多、每穗不育小穗数更少、千粒重更高,低节位分蘖(早发分蘖)比高节位分蘖(迟发分蘖)具有更高的单穗产量[37,38]。
本研究发现低节位早发分蘖特性在水稻品种资源中具有较大的遗传变异,受到多种遗传因素的调控,包括生长素、独脚金内酯、可溶性糖含量以及氮同化相关酶活性等。水稻茎鞘中可溶性糖含量是决定其总茎蘖数和存活率的重要因素[39],在本研究中发现叶鞘中可溶性糖与水稻第1~3 叶位成蘖率无显著相关性,说明在水稻低节位的分蘖芽初步生长阶段,对可溶性糖的要求比较低,只要维持低水平的可溶性糖含量水稻就可以形成分蘖芽。生长素合成相关基因的表达水平与水稻第2、3 叶位成蘖率呈极显著相关,说明在水稻苗期分蘖节中YUCCA6 的表达是影响水稻不同品种低节位分蘖的关键因素之一。SLs 能够抑制植物的分枝和侧芽的生长[40],HTD1、D10 都是SLs 合成途径上的基因,D14、D3 是SLs 信号传导过程中的基因,IPA1 和TB1 是SLs 下游调控基因[18-21],这些基因通过SLs 信号通路抑制水稻侧芽向外生长[41,42]。研究还发现低节位成蘖与SLs 信号通路及其下游调控基因IPA1 和TB1 密切相关,第2 叶位成蘖率与D10、D14、IPA1 和TB1 的相对表达量呈显著相关,第3 叶位成蘖率与HTD1、D14 和TB1 的相对表达量呈显著相关,第2、3 叶位成蘖率与D3 的相对表达量呈极显著相关。说明当腋芽处于分蘖原基状态时,SLs 信号通路上HTD1、D10、D3 和D14 基因以及其下游基因IPA1 和TB1的表达已经开始对苗期低节位分蘖芽的生长起作用。
通过品种改良和栽培调控促进低节位、早发分蘖是水稻高产的重要途径,本研究为探究影响早发分蘖相关因素提供了更多的理论依据,但更细致的机理还需进一步研究。
猜你喜欢
中外葡萄与葡萄酒(2023年4期)2023-07-07
干旱地区农业研究(2022年4期)2022-07-28
四川蚕业(2022年1期)2022-06-06
塔里木大学学报(2021年1期)2021-03-31
蔬菜(2020年1期)2020-01-17
农家科技中旬版(2018年2期)2018-05-05
现代农业(2018年7期)2018-02-18
西南农业学报(2016年5期)2016-05-17
长江蔬菜(2011年2期)2011-03-22
