
深圳七娘山黄桐和乌檀所在风水林群落特征分析
2021-01-04 05:58陈志晖谷超关开朗陈京锐刘逸嵘凡强廖文波
生态科学 2020年6期
陈志晖, 谷超, 关开朗, 陈京锐, 刘逸嵘, 凡强, 廖文波,*
深圳七娘山黄桐和乌檀所在风水林群落特征分析
陈志晖1, 谷超2, 关开朗1, 陈京锐1, 刘逸嵘1, 凡强1, 廖文波1,*
1. 中山大学生命科学学院, 有害生物控制与资源利用国家重点实验室/广东省热带亚热带植物资源重点实验室, 广州 510275 2. 深圳大鹏半岛国家地质公园管理处, 深圳 518002
基于样方调查, 对深圳七娘山高岭古村风水林群落进行1500 m2的样地调查, 分析其物种组成和群落结构特征, 旨在为该群落的保护、利用与发展提供科学依据。结果如下: (1)群落种类丰富, 共有维管植物117种, 其中蕨类植物5科7属8种, 裸子植物1科1属1种, 被子植物49科78属108种。种子植物属的地理成分以热带分布占优, 高达91.13%; (2)群落外貌终年常绿, 无明显季相变化和林冠起伏, 层间结构丰富。其中乔木层以黄桐()、肉实树()和银柴()为主要优势种, 次之有乌檀()、胭脂()等; 灌木层以九节()、大沙叶()为主要优势种, 而草本层以金毛狗()为优势种; (3)群落Raunkiaer频度规律为A>B>C>D>E, C级和D级仅占3.65%, 无E级分布, 可见群落偶见种多, 多优势种相互制约; (4)物种多样性方面, Simpson 多样性指数、Shannon-Wiener 指数和Pielou 均匀度指数均表现为乔木层>灌木层>草本层, 与邻近保存较好的大鹏半岛香蒲桃风水林群落和钝叶假蚊母树群落, 以及东莞的臀果木-黄桐风水林群落相比有着更高的物种多样性和均匀度; (5)优势种群年龄结构分析表明, 乌檀种群为衰退状态, 黄桐和肉实树种群处于增长阶段, 群落在未来的发展中可能会演化成以黄桐和肉实树等中生性树种为主的中生常绿阔叶林。
风水林; 群落特征; 七娘山; 高岭古村
0 前言
风水林是指受传统的文化信仰和风水理论的影响而营建、管理或封育、保护起来的森林[1], 多为原生低海拔阔叶树种的残余部分[2], 是华南地区多数村落极具地域特色的群落类型, 保存有丰富的乡土树种。与西部地区的神山文化相似, 风水林的保护主要也受信仰文化的影响, 所以在一定程度上也反映了该地区的文明和自然环境的变化。风水林作为一种独特的群落类型, 往往比周边的人工林群落有着更高的物种丰富度、植物多样性[3], 此外作为社会历史文化现象, 也有着重要的文化价值。
大鹏半岛位于深圳市东南部海岸, 三面环海, 受近海特殊地理环境的影响, 岛上植被类型多样, 丰富的植物资源多分布在村庄附近的风水林中。张永夏等[4]对岛上的风水林香蒲桃群落与深圳南山假苹婆群落和澳门假苹婆群落作对比, 也显示其物种多样性水平较高。但随着城市化进程的加快, 村庄的变迁以及人们文化观念改变等因素影响, 岛上原始的常绿季雨林和常绿阔叶林已被破坏殆尽, 仅剩部分较为完好的次生林[5]。因发展旅游业, 这类分布于城市中的风水林受人为干扰严重, 其生境破碎化日趋严重。七娘山北侧东风岭的高岭古村为大鹏半岛上古村原始风貌保存较为完好的一个客家村落, 其村落东边保存有一片典型的风水林群落。本文通过对该风水林进行样方调查, 进而对该群落的组成结构、地理成分性质、群落外貌、物种多样性、年龄结构等进行分析, 在掌握该风水林的结构和演替特征的基础上, 为该风水林群落资源的保护开发和促进其可持续发展提供科学依据。
1 材料和方法
1.1 研究地概况
七娘山位于深圳大鹏半岛中南部, 西与香港隔海相望, 北与惠州海域相接, 地理位置为114°26′— 114°37′ E, 22°27′—22°36′ N, 主峰海拔867.4 m, 为深圳第二高峰。七娘山属南亚热带海洋性季风气候, 日照充足, 热量丰富, 降水充沛。夏季高温多雨, 冬季干旱稍冷, 年平均气温为22.3 ℃, 最高气温达36.6 ℃, 最低气温为1.4 ℃, 年平均降雨量为2000毫米。受海风影响, 地形主要以中低山地、丘陵为主。土壤多为酸性的赤红壤和红壤, 土质较粘重。所调查的风水林群落位于七娘山北侧东风岭高岭古村附近, 海拔184 m, 地理坐标为114°32′35.15″ E, 22°32′49.27″ N。
1.2 样地设置与调查
在对整个群落进行全面踏查的基础上, 选择典型地段设置面积为1500 m2的样地, 采用相邻格子法将样地划分为15个10 m×10 m样方, 用每木记账调查法进行调查, 起测径阶≥1.3 cm, 或高度≥1.5 m, 记录树种名、胸径、树高、冠幅等; 并在每个样方内设置1个5 m×5 m的灌木样方, 记录种名、株数、高度和种的盖度; 再设置1个2 m×2 m的草本样方, 记录种名、株数和盖度[6]。
1.3 数据分析
根据群落调查结果, 对七娘山风水林群落的种类组成、群落结构、种群分布格局和物种多样性进行分析。
(1) 计算重要值: 根据《陆地生态系统生物观测规范》[6], 计算群落中各种群的相对密度(D)、相对显著度(P)、相对频度(F)、相对投影盖度(C), 计算出重要值(I)。其中乔木层重要值= D+P+ F, 灌木和草本层重要值= D+C+ F。
(2) 种群年龄结构分析: 以立木级代替年龄级进行分析[7], 采用5级立木划分标准, I级为苗木,<33 cm, II级为小树;≥33 cm,<2.5 cm; III级为壮树, 2.5≤≤7.5 cm; IV级为大树, 7.5≤≤22.5 cm; V级为老树,≥22.5 cm。
(3) 频度分析: 依据Raunkiaer 的频度分类方法, 将频度分为5个等级, 即: 1—20%为A级, 21%—40%为B级, 41%—60%为C级, 61%—80%为D级, 81%—100%为E级。
(4) 物种多样性分析: 采用Margalef 丰富度指数(d)、Simpson 多样性指数()、Shannon-Wiener 多样性指数()和Pielou 均匀度指数(J)进行测度[8]。计算公式分别为:
Margalef 指数:=(-1) / ln
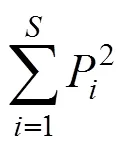
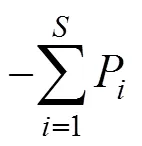
Pielou 均匀度指数:=/ ln
式中为乔木层、灌木层或草本层各层的物种数;P=N/,N为第个物种的个体数量,为乔木层、灌木层或草本层各层的个体数量。
2 结果与分析
2.1 群落物种组成和地理成分分析
根据样地调查统计, 该群落共有维管束植物117种, 隶属55科86属(蕨类植物依据PPG I, 裸子植物依据郑万钧系统, 被子植物依据APG IV)。其中蕨类植物5科7属8种, 裸子植物1科1属1种, 被子植物49科78属108种(表1)。其中国家II级保护植物3种, 分别是白桂木()、土沉香()和金毛狗()。群落组成的主要优势科有: 桑科(Moraceae)3属17种, 叶下珠科(Phyllanthaceae)5属9种, 茜草科(Rubiaceae)6属8种, 樟科(Lauraceae)4属7种, 大戟科(Euphorbiaceae)5属4种, 这些科的种数占总种数的38.46%, 优势科现象明显。
按照吴征镒中国种子植物属的分布区类型划分原则[9], 可将本群落的79属划分为10种类型和2种分布变型(表2)。泛热带分布及其变型占比最大, 有26属, 占总属数的32.91%。该类型的属在群落中多为乔木或藤本, 在群落中起重要作用的有榕属()、杜英属()和苹婆属()。热带亚洲和热带美洲间断分布有3属, 占总属数的3.8%, 多为次优势树种, 如泡花树属()。旧世界热带分布及其变型有18属, 占总属数的22.78%, 仅次于泛热带分布, 在乔木层中起重要作用的有蒲桃属()和血桐属()。热带亚洲(印度-马来西亚)分布有15属, 占总属数的18.99%, 该分布区类型属对群落的影响最大, 如银柴属()、黄桐属()、肉实树属()和波罗蜜属()均为该群落的优势种, 表现出显著的热带性质。此外, 温带分布类型有6属, 占总属数的7.6%, 多为伴生树种, 对群落所起作用较小。由此可见, 本群落中热带亚热带成分占到总属数的91.13%, 且在群落中占据优势地位, 相比于深圳大鹏半岛的香蒲桃风水林群落[4]和香港岛黄桐群落[10]具有更强的热带性质, 但低于东莞凤岗镇臀果木-黄桐风水林群落[11]。这可能与该群落受人为干扰后的物种入侵有关。
2.2 群落外貌和垂直结构特征
该群落为南亚热带低地常绿阔叶林, 以常绿阔叶树种占绝对优势, 混有1株针叶树杉木(), 无落叶树种。群落林冠浓密, 无明显起伏, 终年常绿, 伴有部分山杜英()和中华杜英()的红叶点缀。群落垂直结构分层现象明显, 可划分为乔木层、灌木层、草本层以及由藤本植物和附生植物所构成的层间结构, 不同层间存在着空间重叠和交错分布现象。
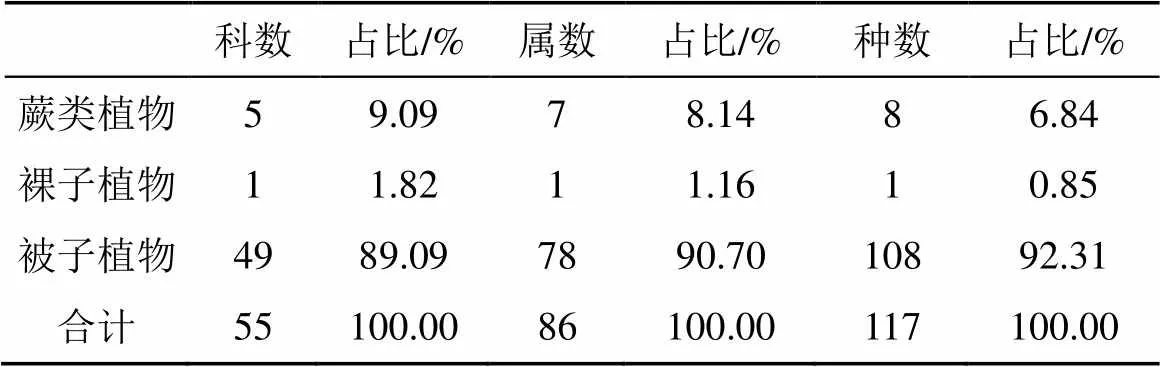
表1 七娘山风水林群落物种组成
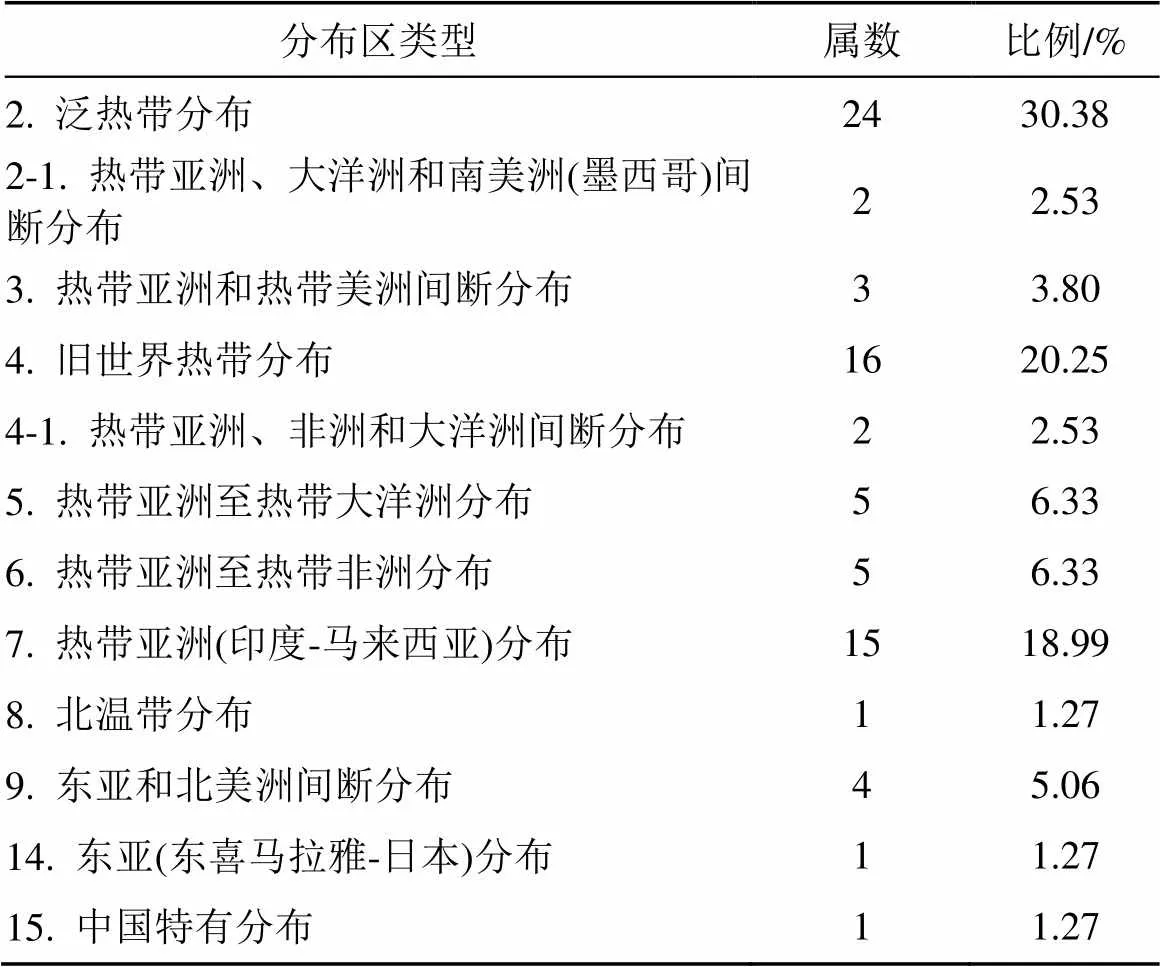
表2 七娘山风水林群落种子植物属的分布区类型
乔木层植物有57种, 以银柴()、黄桐()、肉实树()和胭脂()占优势。按照2 m的间隔对群落不同乔木的高度级进行划分(图1), 可将乔木层分为2个亚层: 第1亚层高度为10—15 m, 平均高度约为11.8 m, 冠幅大而交错, 该层分布有23种37株乔木, 以黄桐、山杜英、中华杜英和银柴为主, 其中高度达15 m的有山蒲桃()和山杜英, 各1株。第2亚层高度为5—10 m, 平均高度约为6.8 m, 分布有33种93株乔木, 以银柴、肉实树、鼎湖血桐()、乌檀()为主, 其它树种还有黄桐、香楠()等。
灌木层高1—4 m, 约为54种208株, 该层种类和数量较多, 但乔木幼树种类和数量较少, 以银柴、肉实树、黄毛五月茶()、山蒲桃为主。其它树种则多为林下耐荫树种, 如九节()、罗伞树()、大沙叶()等。
草本层种类较少, 多为蕨类植物, 其中金毛狗数量最多, 在样地局部连续成片出现, 形成群落。其它较多的蕨类植物则有江南星蕨()、单叶新月蕨()、傅氏凤尾蕨()和半边旗()等。
层间结构丰富, 大型藤本攀附现象明显, 遮蔽林层空间, 以紫玉盘()、锡叶藤()、木防己()等木质藤本为主, 其它藤本植物还有夜花藤()、粉背薯蓣(var.)、网脉崖豆藤()等。
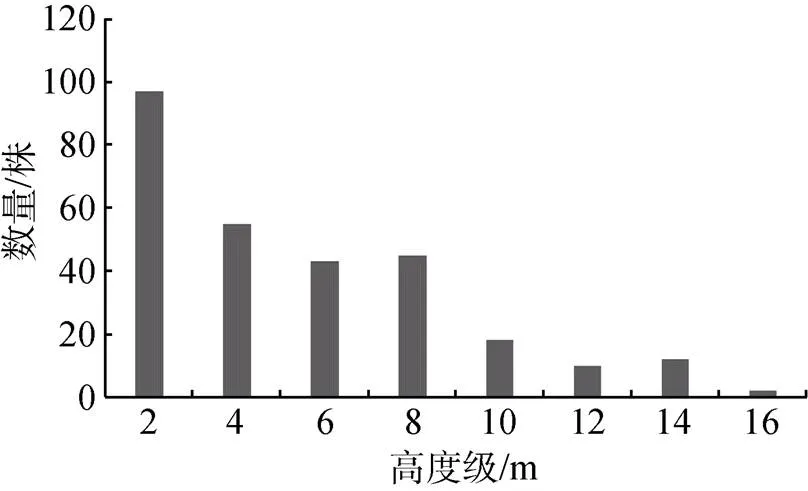
图1 七娘山风水林群落高度级频率分布图
Figure 1 The height class frequency distribution at the Fengshui woods of Qiniang Mountain
2.3 群落频度分析
依据Raunkiaer 频度定律对本群落进行频度分析, 其中A级频度比例高达83.94%, B级为12.41%, C级为2.92%, 而D级仅有0.73%, 无E级频度分布, 分布结构为A>B>C>D>E, 与Raunkiaer 标准频度定律A>B>C≥D
2.4 群落优势种和重要值分析
分层计算本群落各物种的重要值, 列出乔木层、灌木层和草本层重要值>6的树种(表3), 其中乔木有58种, 以银柴、黄桐和肉实树占优, 群落优势种明显, 但无明显建群种, 为多优势群落; 灌木54种, 以九节和大沙叶()占优, 分布有较多的肉实树和银柴幼树; 因林冠郁闭度高, 草本层较为稀疏, 仅有26种, 多以耐荫蕨类为主, 其中金毛狗数量多达58株, 冠幅大而密集, 乔木幼苗仅见黄毛五月茶。
2.5 风水林群落命名
根据宋永昌[13]对中国植被的划分原则, 该风水林的植被型为适雨常绿阔叶林(亚热带雨林), 树种主要以大戟科、肉实树科和桑科等热带性较强的科属组成, 优势种主要为黄桐、银柴、肉实树, 标志种为乌檀。群丛的命名规则参照宋永昌系统[14], 以优势种为主, 一般不超过2种, 辅以特征种组或标志种组的代表植物来命名; 将标志种置于优势种前, 用“-”连接; 如果优势种之间优势度区别不明显, 并出现种类相互交错时, 则在种名之间用“ 、”号或“, ”号联结, 前后顺序并不严格表示优势度的大小。基群丛的命名在群丛名称后面用“-”连接上灌木层和草本层的优势种。本研究中, “优势种”为样地中乔木层重要值最高的种, “标志种”选取该群落的代表性物种。结合各层树种重要值, 此群落可命名为Ass.-、--(即乌檀-银柴、黄桐-九节-金毛狗群落)
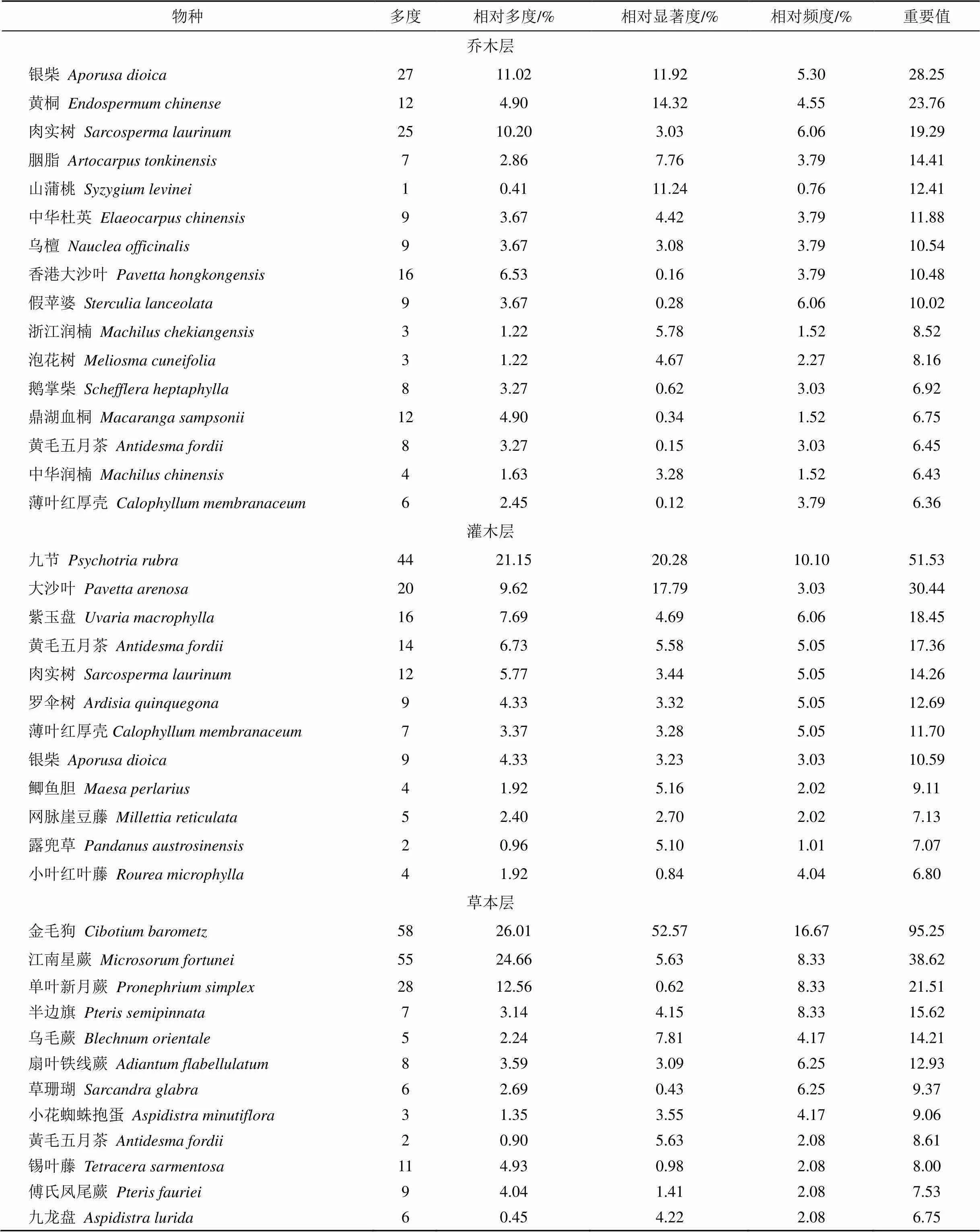
表3 七娘山风水林群落乔木层、灌木层和草本层优势种重要值
注: 表中仅列出重要值>6的树种; 其中乔木层16种, 灌木层12种, 草本层12种; 重要值<6的乔木层树种有42种, 灌木层有42种, 草本层有14种, 略。
2.6 群落的物种多样性分析
物种多样性是指种数量及其个体分配均匀度两者的综合[15], 是群落特征的一个重要方面, 对物种多样性的研究可以更好地认识群落的组成、变化和发展。对本群落的物种多样性进行分析可知(表4), Margalef 指数中乔木层的丰富度指数最高, Simpson 多样性指数在各层次上的表现规律与Shannon- Wiener 多样性指数一致, 均为乔木层>灌木层>草本层, 此外Pielou 均匀度指数上也呈现乔木层>灌木层>草本层的格局。乔木层的Simpson 指数、Shannon-Wiener 指数和Pielou 均匀度指数均高于灌木层和草本层, 不符合自然生态系统中各层次的物种多样性灌木层>乔木层>草本层的格局[16], 可能是因为乔木层优势种明显, 郁闭度大, 由此对灌木的生长产生抑制, 也导致乔木的幼苗和幼树分布较少, 另外下层树种受林冠的遮挡多分布于林窗和空旷区域, 导致均匀度较低, 由此也说明该群落处于一个较高的发展阶段。
群落的物种多样性不仅由群落本身的类型结构决定, 也受其演替阶段、微生境和人为活动的影响, 因此可作为定量数值反映群落的稳定性和动态演替[17]。与地理位置邻近的其它风水林群落进行对比(表5), 本群落的物种多样性指数低于香港仔郊公园保护和管理较好的黄桐群落, 高于大鹏半岛香蒲桃风水林群落和钝叶假蚊母树群落以及东莞臀果木-黄桐风水林群落, 但均匀度指数均高于其它4个群落。
2.7 群落优势种群的年龄结构分析
种群的年龄结构是种群内不同年龄个体数量的分布情况[19], 群落内种群个体的龄级分布不仅可以说明种群动态与发展趋势, 也可以在一定程度上反映种群的稳定性以及种群与环境之间的互动作用, 进而揭示种群在群落中的地位和作用[20]。选取本群落乔木层中重要值较高的银柴、黄桐、肉实树和珍贵资源树种乌檀进行种群的年龄结构分析(图2), 从图中可以看到, 乔木层中重要值最高的银柴种群以IV、V级立木占优, III、II、I级立木依次减少, 呈倒金字塔型结构, 但银柴作为地带性物种, 适应性强, 因此并不意味着为衰退种群; 乌檀种群类似, 以大径级个体占优, 并缺乏幼苗储备, 这可能是由于乌檀种群天然更新能力差[21], 加之群落内树种间竞争大, 由此处于衰退状态; 而黄桐种群IV、V、III级立木大致呈金字塔型, 但仍有一定的幼苗储备, 说明种群增长减缓, 趋向稳定期, 因此仍然能在一定时期内保持较大优势; 肉实树以I、II级苗木占优, 无V级立木分布, 该种群正处于发展阶段, 发展潜力大, 结合其在群落中的分布数量, 推测该种群未来可能代替银柴, 与黄桐组成多优势群落。
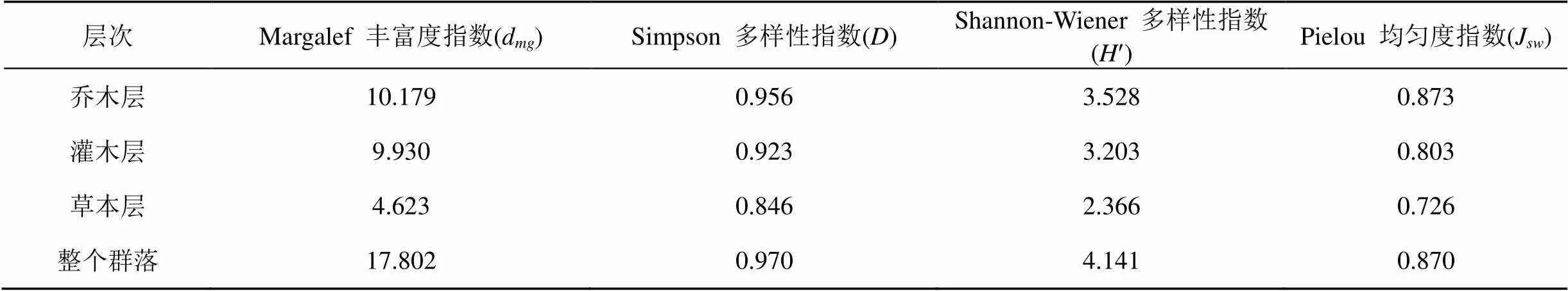
表4 七娘山风水林群落各层次的物种多样性指数
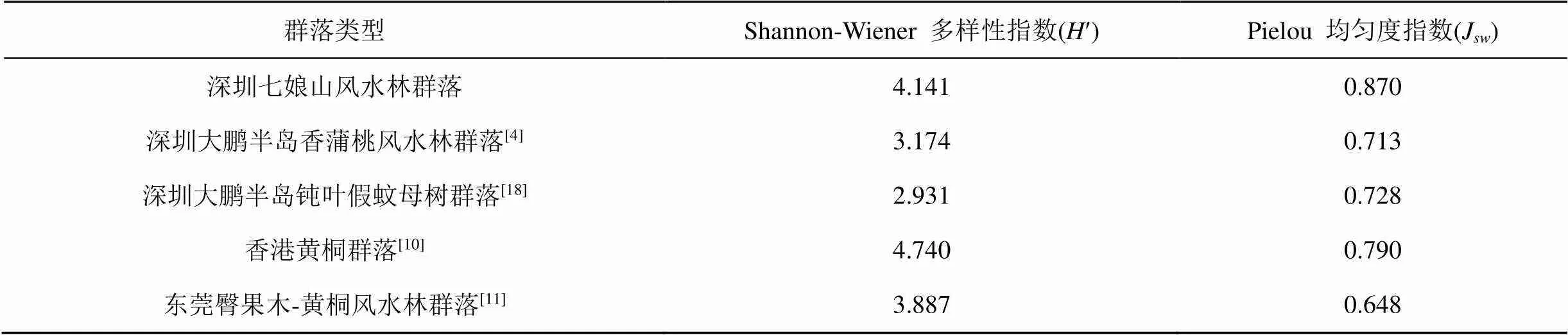
表5 不同地区群落的物种多样性指数对比
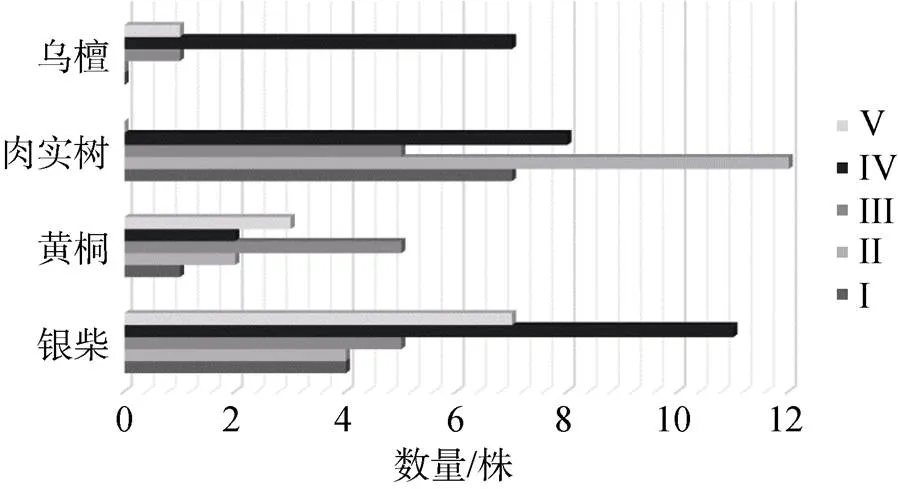
图2 七娘山风水林群落乔木层主要优势种年龄结构图
Figure 2 Age-structure of tree layer dominant species at the Fengshui woods of Qiniang Mountain
3 讨论与结论
深圳七娘山高岭古村乌檀-银柴、黄桐-九节-金毛狗风水林群落共有植物117种, 隶属55科86属, 树种主要分布于桑科、叶下珠科、茜草科和大戟科等热带、亚热带科属, 其中珍稀濒危植物有白桂木、土沉香和金毛狗, 珍贵资源树种有乌檀。群落外貌终年常绿, 垂直分层现象明显, 不同层间存在渗透和镶嵌分布现象, 层间藤本植物丰富, 较典型的南亚热带常绿阔叶林有着更为复杂的垂直结构特征。在地理成分上, 以热带亚热带成分占优, 热带性质显著。
Raunkiaer 频度级分析表明, 该群落频度级分布规律为A>B>C>D>E, 其中C、D级仅占3.65%, 分别为肉实树、假苹婆、银柴、金毛狗和九节, E级缺乏, 该分布规律与热带、亚热带常绿阔叶林频度规律一致[22]。南亚热带常绿阔叶林结构较为复杂, 种类丰富, 因此A级与E级频度呈现两极分化, 群落中偶见种多, 分布不均匀, 物种多样性倾向于集群分布[23]。结合该群落树种的重要值进行分析, 频度级为C级的银柴、肉实树和频度级为B级的黄桐都占有较高的重要值, 另外从优势种群的年龄结构也可以看到, 银柴种群处于成熟衰退型, 而黄桐种群和肉实树种群均为增长型, 种群间相互抑制扩张, 加之群落郁闭度高, 乔木种子更新困难, 于是形成了多优势种的发展状态。此外有研究表明样方数目的增加也会使得群落中A级种的数量增多, 而E级种数量降低[24]。
物种多样性比较中, 本群落的Simpson 多样性指数为0.970, Shannon-Wiener 多样性指数为4.141, 低于香港黄桐群落, 但高于大鹏半岛的香蒲桃风水林群落和钝叶假蚊母树群落以及东莞臀果木-黄桐风水林群落。Shannon-Wiener 多样性指数是衡量南亚热带森林群落物种多样性水平较好的一个指数[25], 从不同冠层来看, 七娘山风水林群落的Shannon-Wiener 多样性指数表现为乔木层>灌木层>草本层, 与处于进展演替阶段的东莞臀果木-黄桐群落结构一致, 而处于演替中期阶段大鹏半岛的香蒲桃风水林群落和钝叶假蚊母树群落其Shannon- Wiener 多样性指数则表现为灌木层>乔木层>草本层, 乔木层的物种多样性会随着演替过程而增加, 灌木层和草本层则是先增加后下降[26], 乔木层在物种多样性和均匀度上高于灌木层, 表明群落处于较为高级的发展阶段。结合群落中4个优势种群的年龄结构分析, 优势种群银柴开始处于次优状态, 黄桐和肉实树种群表现为增长型, 在未来发展中, 群落中生性树种比例将继续增加, 可能会演化为黄桐、肉实树和假苹婆等中生性树种为主的中生常绿阔叶林。
风水林曾遍布华南地区, 但其中只有较为少数能保存下来[2]。该风水林是深圳近年来发现保存较为完好的一个群落, 群落内分布着多种珍稀濒危植物和珍贵资源树种, 比周边其他风水林群落有着更高的物种丰富度和多样性。乌檀种群天然更新困难, 其分布多呈零星状, 大都分布狭窄, 为中海拔森林中少见的乔木树种[21], 因此选择乌檀作为该风水林群落的标志种。此外该群落以银柴、黄桐作为优势种, 银柴是亚热带常绿阔叶林的地带性物种, 黄桐作为南亚热带低地常绿阔叶林的代表类群, 以大戟科、桑科、肉实树科为优势科, 明显区别于以壳斗科和樟科植物为主要建群种的南亚热带常绿阔叶林类型[10], 有着热带雨林向南亚热带阔叶林的过渡性特征。风水林的保护多依靠周边村落, 而高岭古村的村民们因交通不便自上世纪90年代便搬离该地, 古村后来被纳入大鹏半岛国家地质公园范围, 由此吸引了不少驴友前去登山探险, 在调查时已受到一定的人为干扰。因此该群落作为珍贵物种的资源库, 建议应停止相关的旅游开发和城建项目, 采取相应的封山育林措施, 限制游客进入, 减少过度的人为干扰, 促进该风水林的稳定性和自然进展演替。
[1] 李仕裕, 叶育石, 王发国, 等. 广州市风水林植物组成及分布区类型分析[J]. 植物资源与环境学报, 2013, 22(1): 102–109.
[2] ZHUANG Xueying, CORLETT R. Forest and forest succession in Hong Kong, China[J]. Journal of Tropical Ecology, 1997, 13(6): 857–866.
[3] 杨期和, 潘素芳, 赖万年, 等. 粤东桥溪村风水林群落特征初探[J]. 广西植物, 2015, 35(6): 833–841.
[4] 张永夏, 陈红锋, 秦新生, 等. 深圳大鹏半岛“风水林”香蒲桃群落特征及物种多样性研究[J]. 广西植物, 2007(4): 596–603.
[5] 张荣京, 张永夏, 严岳, 等. 深圳大鹏半岛常绿季雨林和常绿阔叶林群落物种多样性分析[J]. 山地学报, 2005(4): 4495–4501.
[6] 中国生态系统研究网络科学委员会. 陆地生态系统生物观测规范[M]. 北京: 中国环境科学出版社, 2007.
[7] PRPCTOR J, LEE Y F, LANGLEY A M, et al. Ecological Studies on Gunung Silam, A Small Ultrabasic Mountain in Sabah, Malaysia. I. Environment, Forest Structure and Floristics[J]. Journal of Ecology, 1988, 76(2): 320–340.
[8] 孙儒泳. 基础生态学[M]. 北京: 高等教育出版社, 2002.
[9] 吴征镒. 中国种子植物属的分布区类型[J]. 植物资源与环境学报, 1991(S4): 1–139.
[10] 王伯荪, 陆阳, 张宏达, 等. 香港岛黄桐森林群落分析[J]. 植物生态学与地植物学学报, 1987(4): 241–251.
[11] 吕浩荣, 刘颂颂, 叶永昌, 等. 东莞凤岗镇臀果木-黄桐风水林群落特征分析[J]. 热带亚热带植物学报, 2009, 17(2): 137–145.
[12] 胡玉佳. 海南岛龙脑香森林的群落特征及其类型[J]. 生态科学, 1983(2): 16–24.
[13] 宋永昌. 对中国植被分类系统的认知和建议[J]. 植物生态学报, 2011, 35(8): 882–892.
[14] 宋永昌. 中国常绿阔叶林分类试行方案[J]. 植物生态学报, 2004(4): 435–448.
[15] KERSHAW K A. Quantitative and dynamic plant ecology [M]. London:Edward Arnold, 1985.
[16] 贺金生, 陈伟烈, 李凌浩. 中国中亚热带东部常绿阔叶林主要类型的群落多样性特征[J]. 植物生态学报, 1998(4): 16–24.
[17] 彭少麟, 王伯荪. 鼎湖山森林群落分析Ⅰ.物种多样性[J]. 生态科学, 1983(1): 11–17.
[18] 刘海军, 郭强, 张信坚, 等. 深圳大鹏半岛自然保护区钝叶假蚊母树群落特征[J]. 生态科学, 2018, 37(2): 182–190.
[19] 张育新, 马克明, 祁建, 等. 北京东灵山海拔梯度上辽东栎种群结构和空间分布[J]. 生态学报, 2009, 29(6): 2789–2796.
[20] 刘东蔚, 王海军, 陈勇, 等. 深圳羊台山黄牛木群落学特征研究[J]. 生态科学, 2014, 33(2): 379–385.
[21] 陈健妙. 珍稀野生植物乌檀的开发利用[J]. 中国野生植物资源, 2003(4): 38–39.
[22] 张信坚, 邱建勋, 胡玮珊, 等. 江西信丰细迳坑自然保护区观光木群落研究[J]. 亚热带植物科学, 2016, 45(4): 343–350.
[23] 李薇, 朱丽萍, 汪春燕, 等. 深圳市内伶仃岛山蒲桃+红鳞蒲桃-小果柿群落结构及其物种多样性特征[J]. 生态科学, 2018, 37(2): 173–181.
[24] 温远光. 大明山森林群落的频度分析[J]. 广西农业大学学报, 1998(2): 195–198.
[25] 彭少麟, 周厚诚, 陈天杏, 等. 广东森林群落的组成结构数量特征[J]. 植物生态学与地植物学学报, 1989(1): 10–17.
[26] 贺金生, 陈伟烈, 江明喜, 等. 长江三峡地区退化生态系统植物群落物种多样性特征[J]. 生态学报, 1998(4): 65–73.
Coenological characteristics ofandat the Fengshui woods of Qiniang Mountain, Shenzhen
CHEN Zhihui1, GU Chao2, GUAN Kailang1, CHEN Jingrui1, LIU Yirong1, FAN Qiang1, LIAO Wenbo1,*
1. State Key Laboratory of Biocontrol & Guangdong Provincial Key Laboratory of Plant Resources, School of Life Sciences, Sun Yat-sen University, Guangzhou 510275, China 2. Shenzhen Dapeng Peninsula National Geopark, Shenzhen 518002, China
In order to provide a scientific basis for the protection, utilization and development of the Fengshui woods of Gaoling Village in Qiniang Mountain of Shenzhen city, a 1500m2quadrat sampling method was conductedinits Fengshuiwoods to assess the species composition and community structure. In this study, 108 species, derived from 49 families and 78 genera, were seed plants; 8 species, derived from 5 families and 7 genera, were pteridophytes; and one gymnosperm species, were recorded. Thegeographical element characteristic of seed plants genuswasdominatedbytropicaldistribution, up to 91.13%. The forest communitywas evergreen, with no obviousseasonal changes and canopy fluctuations,andwas withrichinterlayerstructure. For dominant tree species were,andfollowedby,, etc; the dominant shrubs specieswereand;thedominant herbs specieswas the. The Raunkiaer’s law of frequency was calculated to be A>B>C>D>E, whereby C and D were only 3.65% and none for E. The Shannon-wiener index, Simpson index, and Pielou index showed that biodiversity richness was tree layer>shrub layer>herb layer. When compared with the adjacent well-conserved fengshui woods ofandcommunities in Dapeng Peninsula and the-community in Dongguan, Higher species diversity was observed for the-community in Qiniang Mountain. The age structure analysis of the dominant species showed that the population size ofwas declining, while the population sizes ofandwas increasing. We concluded that this Fenshui woods might evolve into a mesophytic evergreen broad-leaved forest dominated byandin the future.
fengshui woods; community characteristic; Qiniang mountain; ancient Gaoling village
10.14108/j.cnki.1008-8873.2020.06.021
Q948.15
A
1008-8873(2020)06-167-08
2019-10-10;
2019-11-26
深圳大鹏半岛国家地质公园七娘山溪流域生物多样性调查及保护规划项目(HT-99982019-0430); 深圳市自然生态系统植物调查项目(092018121848v1); 广东省高等学校本科质量工程项目(2016-2019)
共同第一作者: 陈志晖(1995—), 男, 硕士研究生, 主要从事植物分类及区系学研究, E-mail: chenzhh93@mail2.sysu.edu.cn; 谷超(1988—), 男, 硕士, 主要从事七娘山地质公园科研和管理工作, E-mail:68606813@qq.com
廖文波, 男, 博士, 教授, 主要从事植物分类学研究, E-mail: lsslwb@mail.sysu.edu.cn
陈志晖, 谷超, 关开朗, 等. 深圳七娘山黄桐和乌檀所在风水林群落特征分析[J]. 生态科学, 2020, 39(6): 167–174.
CHEN Zhihui, GU Chao, GUAN Kailang, et al. Coenological characteristics ofandat the Fengshui woods of Qiniang Mountain, Shenzhen[J]. Ecological Science, 2020, 39(6): 167–174.
猜你喜欢
建筑与预算(2022年6期)2022-07-08
农村实用技术(2020年1期)2020-12-16
海峡姐妹(2018年4期)2018-05-19
现代园艺(2018年14期)2018-01-18
中华建设(2017年3期)2017-06-08
妇女之友(2017年3期)2017-04-20
海峡姐妹(2016年2期)2016-02-27
中国药物应用与监测(2015年5期)2015-12-11
小说月刊(2014年8期)2014-04-19
学与玩(2009年2期)2009-03-09
