
山西苹果褪绿叶斑病毒的分布及遗传多样性分析
2021-01-04 12:26续海红王燕飞窦彦鑫陈伟杨凯
植物保护 2021年6期
关键词:遗传变异
续海红 王燕飞 窦彦鑫 陈伟 杨凯
摘要 :蘋果褪绿叶斑病毒Apple chlorotic leaf spot virus (ACLSV) 是侵染苹果的主要潜隐性病毒之一,在我国苹果植株上发生普遍,严重威胁我国苹果的品质与产量。本研究从山西省12个苹果主产区随机采集360份表现褪绿和斑驳等症状的苹果叶片作为研究样本,通过RT-PCR检测,360份样本中有209份样本为ACLSV阳性,对209份阳性样本的外壳蛋白(coat protein, CP)基因进行分离、测序、克隆,得到12个新的ACLSV分离物(分别命名为 Shanxi 1~Shanxi 12)。选择17个来自不同国家的分离物与12个新的ACLSV分离物在核苷酸和氨基酸层面上进行序列一致性和系统发育分析。结果显示,29个ACLSV分离物被划分为2个不同进化群体。进一步对2个不同ACLSV群体进行选择压分析和中性检验,结果表明,组Ⅰ与组Ⅱ的ACLSV群体之间存在明显的遗传差异,其中负向选择可能是ACLSV遗传变异的原因之一。本研究较全面地分析了ACLSV的发生、危害,并对山西苹果的ACLSV分离物进行了遗传结构分析,为山西苹果褪绿叶斑病毒病的防治提供了理论指导。
关键词 :山西苹果; 隐性病毒; 苹果褪绿叶斑病毒; 外壳蛋白; 遗传变异
中图分类号:
S 432.41
文献标识码: A
DOI: 10.16688/j.zwbh.2020578
Analysis on distribution and genetic diversity of Apple leaf spot virus from Shanxi province
XU Haihong1, WANG Yanfei2, DOU Yanxin1, CHEN Wei2*, YANG Kai1,3
(1.Shanxi Key Laboratory of Germplasm Improvement and Utilization in Pomology, Institute of Pomology,
Shanxi Agricultural University, Taiyuan 030031, China; 2.School of Life Science, Shanxi Normal University,
Linfen 041000, China; 3. Institute of Cotton Research, Shanxi Agricultural University, Yuncheng 044000, China)
Abstract
Apple chlorotic leaf spot virus (ACLSV) is one of the main recessive viruses infecting apples.It occurs widely on apple plants in China, posing a serious threat to the quality and yield of apples in China.In this study, 360 samples of apple leaves with symptoms such as fading and mottling were randomly collected from 12 major apple production areas in Shanxi province.Based on RT-PCR test results, 209 out of 360 samples showed ACLSV-positive.The CP genes of 12 new ACLSV isolates, named as Shanxi 1—Shanxi 12, were isolated, sequenced and cloned from these positive samples.Phylogenetic analysis was performed based on the nucleotide and amino acid sequences of the CP genes of 29 ACLSV isolates (17 were obtained online).The results showed that these isolates were divided into two distinct evolutionary populations, with a possible relation to the region and host.The genetic diversity of different ACLSV populations was further studied.Selective pressure analysis and neutrality test showed that these isolates had significant genetic differences between group Ⅰ and group Ⅱ.Negative selection might play an important role in the genetic variation of ACLSV.The knowledge on the occurrence, damage, genetic structure and evolutionary mechanism of Shanxi ACLSV is useful for the design of sustainable management strategies for controlling ACLSV.
Key words
apple; recessive viruses; Apple chlorotic leaf spot virus; coat protein; genetic diversity
苹果是世界四大水果之一,广泛分布在许多国家和地区,如中国、欧洲、日本、澳大利亚和美国。中国作为最大的苹果生产和消费国,天然的地理环境形成了很多苹果优势种植区,苹果的种植面积和产量、人均占有量和出口量等均居世界前列[12],已成为农民增收,脱贫致富的重要支柱产业[34]。近些年随着黄土高原优势产区面积增加,渤海湾产区果园更新及西南冷凉高地新型果园的崛起,苹果病毒病成为制约果园发展的首要病害,其中苹果褪绿叶斑病毒病发生最为普遍,严重影响了苹果的产量和质量以及果农的经济效益[5]。目前苹果病毒病缺乏有效的防治方法,因此苹果病毒病的系统研究对苹果产业的健康发展具有重要意义。
苹果病毒病在世界范围内广泛传播。目前已报道侵染苹果的病毒有39种,中国已鉴定的苹果病毒有17种[68]。苹果病毒分为潜隐性病毒和非潜隐性病毒,其中苹果茎沟病毒Apple stem grooving virus (ASGV)、苹果褪绿叶斑病毒Apple chlorotic leaf spot virus (ACLSV)、苹果茎痘病毒Apple stem pitting virus (ASPV)是苹果上报道的最主要的3个潜隐性病毒[911]。ACLSV是侵染苹果的主要潜隐性病毒之一,属于乙型线形病毒科Betaflexiviridae,纤毛病毒属Trichovirus[12]。ACLSV为正義单链RNA病毒,病毒粒子呈长线状或丝状,基因组大小约7 545~7 555 nt,5′端带有帽状结构, 3′端有一个Poly(A)尾,编码3个开放式阅读框(open reading frames, ORFs)[1315]。ORF1编码蛋白大小为1 877~1 887 aa,分子量约为216 kDa。具有依赖RNA的RNA聚合酶(Pol)、类木瓜蛋白酶(P-Pro)、解旋酶(Hel)和甲基转移酶(Mt)活性,在ACLSV复制过程中发挥重要功能[16]。 ORF2编码运动蛋白(movement protein, MP),大小为446~460 aa,分子量约为50 kDa。参与ACLSV在细胞间的运动 [17]。ORF3编码ACLSV的外壳蛋白(coat protein, CP)[18],大小为193 aa,分子量约为22 kDa。CP蛋白是重要的结构蛋白,序列相对保守, N端高度可变,是ACLSV株系划分的重要依据[1920]。
本研究于2015年-2016年在山西省12个苹果主产区随机采集360份表现为褪绿和斑驳等症状的新鲜苹果叶片。对ACLSV发生危害、分子变异等情况进行系统性调查和鉴定,了解ACLSV在山西苹果主产区的遗传与流行规律。此外,本研究借助生物学与生物信息学对分离获得的山西ACLSV全长CP基因进行遗传结构和分子变异分析,为山西ACLSV的防治提供参考。
1 材料与方法
1.1 植物样品采集
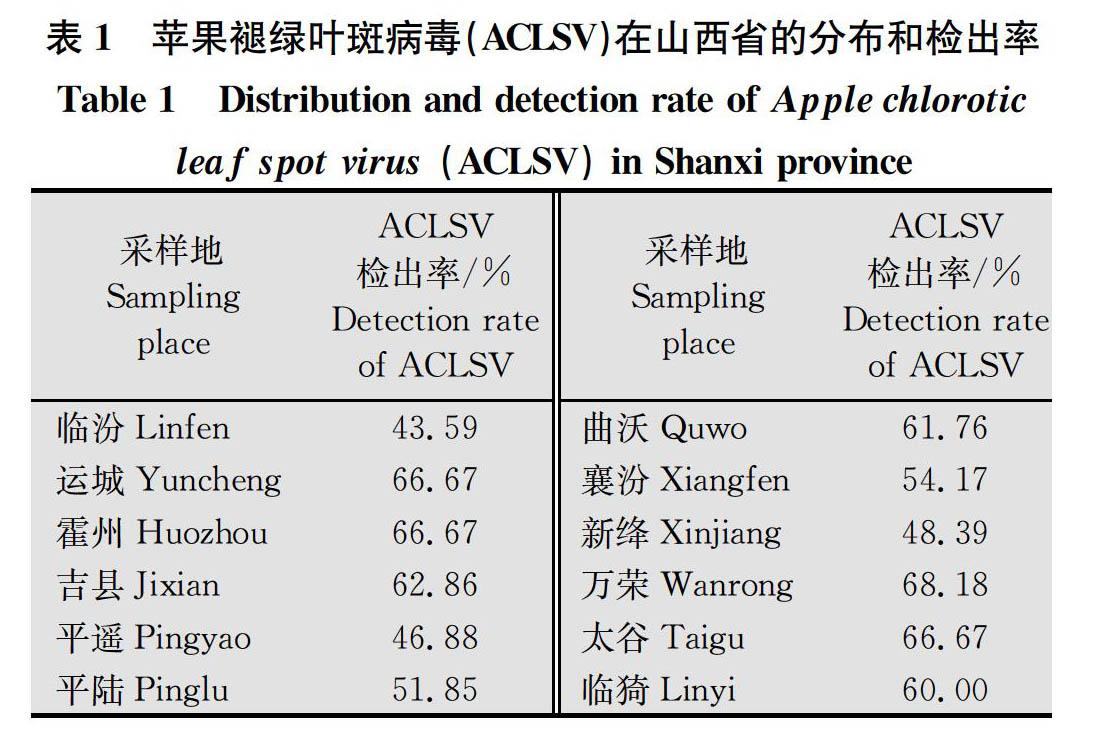
2015年和2016年的4月-6月,在山西省12个苹果主产区(临汾市尧都区、运城市区、霍州、吉县、临猗、平遥、平陆、曲沃、太谷、万荣、襄汾、新绛)开展ACLSV病毒病调查,采集360份表现为褪绿或斑驳等症状的新鲜苹果叶片,迅速放入冰盒,在24 h内送实验室分析。
1.2 ACLSV引物设计
根据GenBank数据库中的ACLSV(登录号为 KC935956)序列中6727-6746位核苷酸设计引物ACLSV-CPF(5′-GAGARTTTCAGTTTGCTMGR-3′,M=A/C;R=A/G),7519-7543位核苷酸设计引物ACLSV-CPR(5′-AGTCTACAGGCTATTTATTATAAGT-3′),用于ACLSV检测和全长CP基因克隆。目标片段长度为817 bp。
1.3 RNA提取和RT-PCR扩增与测序
使用植物总RNA提取试剂盒(Tiangen公司)提取苹果叶片样品的总RNA。使用Quant cDNA 第一链合成试剂盒(Tiangen公司)合成cDNA。以cDNA为模板进行PCR扩增,反应程序:94℃预变性3 min;94℃变性1 min,52℃退火1 min,72℃延伸2 min,35个循环; 72℃ 延伸10 min,4℃保存。反应体系为25 μL,其中,cDNA 2 μL,2×PCR Master Mix 12.5 μL,10 μmol/L正、反向引物各1 μL,ddH2O 8.5 μL。2%琼脂糖凝胶电泳检测RT-PCR扩增产物,使用琼脂糖凝胶纯化试剂盒(CW2302M,康维试剂)纯化并回收阳性扩增条带,ACLSV回收产物与PMD19-T载体16℃过夜连接,连接产物在水浴锅中42℃热激90 s转化大肠杆菌 DH-5α,LB培养基(含有50 μg/mL Amp)上筛选阳性菌落,经PCR鉴定阳性的菌液送擎科生物技术有限公司测序。每个ACLSV阳性样本至少进行3次重复测序。
1.4 山西ACLSV群体的遗传多样性分析
对本研究获得的山西ACLSV的CP基因序列在NCBI上进行比对分析,获得17条来自不同国家和寄主上的,与本研究序列相似的ACLSV CP基因序列。采用SDTv 1.2软件[20]计算核苷酸序列和氨基酸序列的一致性。使用MEGA 7.0软件[21]对本研究分离并克隆的山西ACLSV CP基因进行遗传变异分析。基于山西ACLSV分离物CP基因核苷酸序列,通过最大似然法(construct/test maximum likelihood tree)中的general time reversible模型构建系统发育树,自展值设定为1 000,低于50%的进化分支在本研究被自动舍弃。
1.5 山西ACLSV种群特征与遗传学分析
借助DnaSP 5.10.01 软件,基于CP基因的核苷酸序列,进行山西 ACLSV种群遗传与进化分析,计算参数按照软件设定参数进行[22]。
同时,计算Tajima’s D、Fu & Li’s D 和Fu & Li’s F 参数,对本研究获得的山西ACLSV分离物进行中性检验[23]。使用DnaSP 5.0计算山西ACLSV分离物CP基因非同义突变频率(Ka)和同义突变频率(Ks)的比值,进行选择压分析,若Ka/Ks=1,表示这些群体处在中性或者近似中性选择中;Ka/Ks<1,群体处在负向选择中;如果Ka/Ks>1,群体处在正向选择中。
1.6 山西ACLSV种群重组分析
基于CP基因的核苷酸序列,利用重组检测软件RDP 4.31自带的7种检测方法(MaxChi、RDP、SiScan、BootScan、Chimaera、GENECONV和3Seq)对ACLSV不同分离物重组和重组断点进行自动识别,若至少4种不同方法检测到同一重组事件,且P<10-6,则该事件被认定为阳性重组事件[2425]。
1.7 山西ACLSV种群差异检验和基因流分析
利用DnaSP 5.10.01 软件,基于CP基因的核苷酸序列,分别计算反映山西ACLSV种群差异的5个检验值(Kst*、Ks*、Snn、 Z* 和Fst)[26]。通常在没有遗传分化的情况下,Kst*、Ks*、Snn、 Z*的值接近于0。通过1 000次检验,4个检验值(Kst*、Snn 、Ks*、Z*)均有统计学意义则能反映ACLSV种群遗传差异情况[27]。Fst作为重要的基因分化系数,常被用来衡量种群间多样性程度。一般来说,未分化至完全分化群体的Fst绝对值范围为0~1。Fst的绝对值>0.33,表明基因流动不频繁;Fst的绝对值<0.33,表明基因流动频繁[28]。
2 结果与分析
2.1 山西ACLSV发生情况
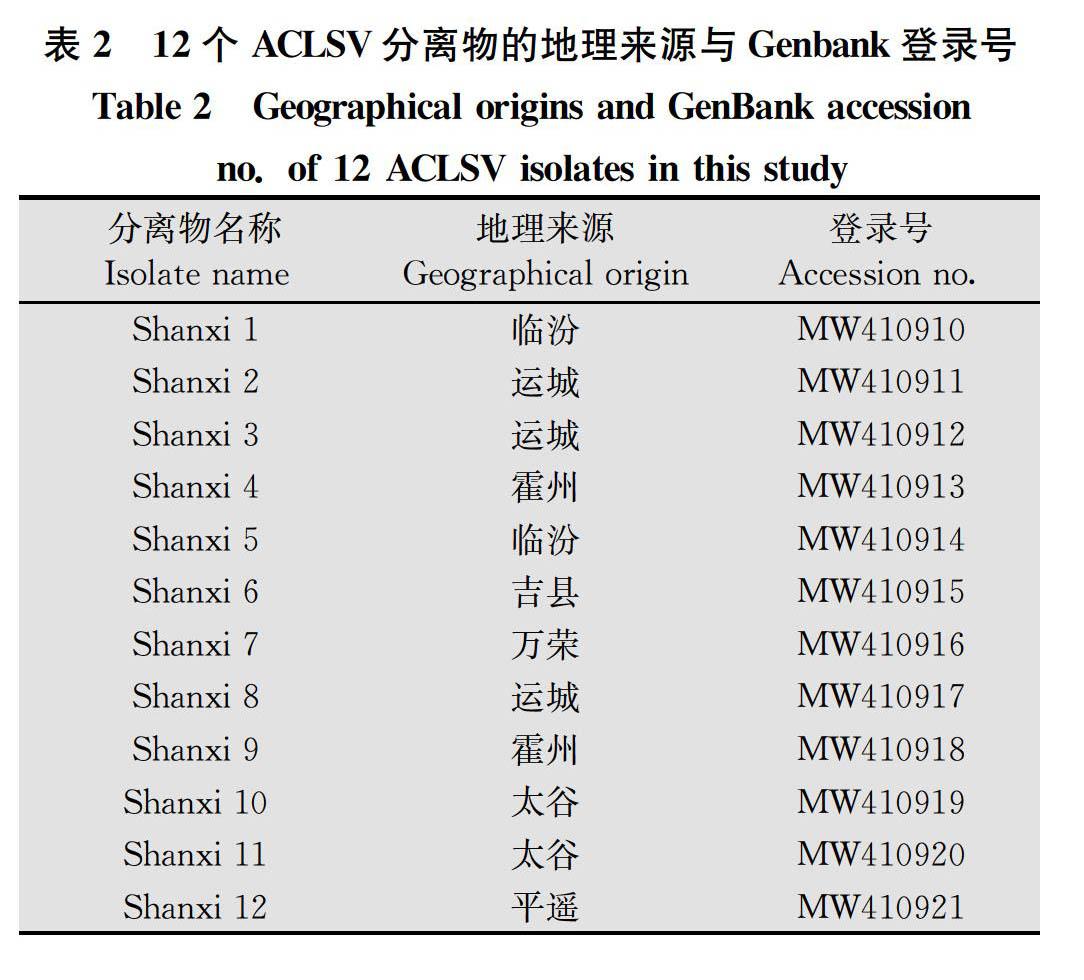
本研究在山西省12个苹果主产区随机采集了360份具有ACLSV特征的叶片样本, 通过RT-PCR检测发现其中有209份样本被ACLSV浸染,各采样点检出率为43.59%~68.18%,平均检出率为58.06%(表1)。万荣地区检出率最高,为68.18%,其余依次为运城、霍州、太谷、吉县、曲沃、临猗、襄汾、新绛、平遥和临汾。临汾的检出率最低,为43.59%。对209份样品的RT-PCR产物回收测序后,通过NCBI-BLAST分析(https:∥www.ncbi.nlm.nih.gov/),有197份样品的RT-PCR产物核苷酸序列与已公布的ACLSV 的核苷酸和氨基酸序列一致性为100%,为已知的ACLSV分离物;而其余12份样品RT-PCR产物核苷酸序列与已登录的ACLSV分离物在核苷酸和氨基酸水平均有差异,为新的ACLSV分离物(Shanxi 1~Shanxi 12)(表2)。
2.2 山西 ACLSV 群体的遗传多样性分析
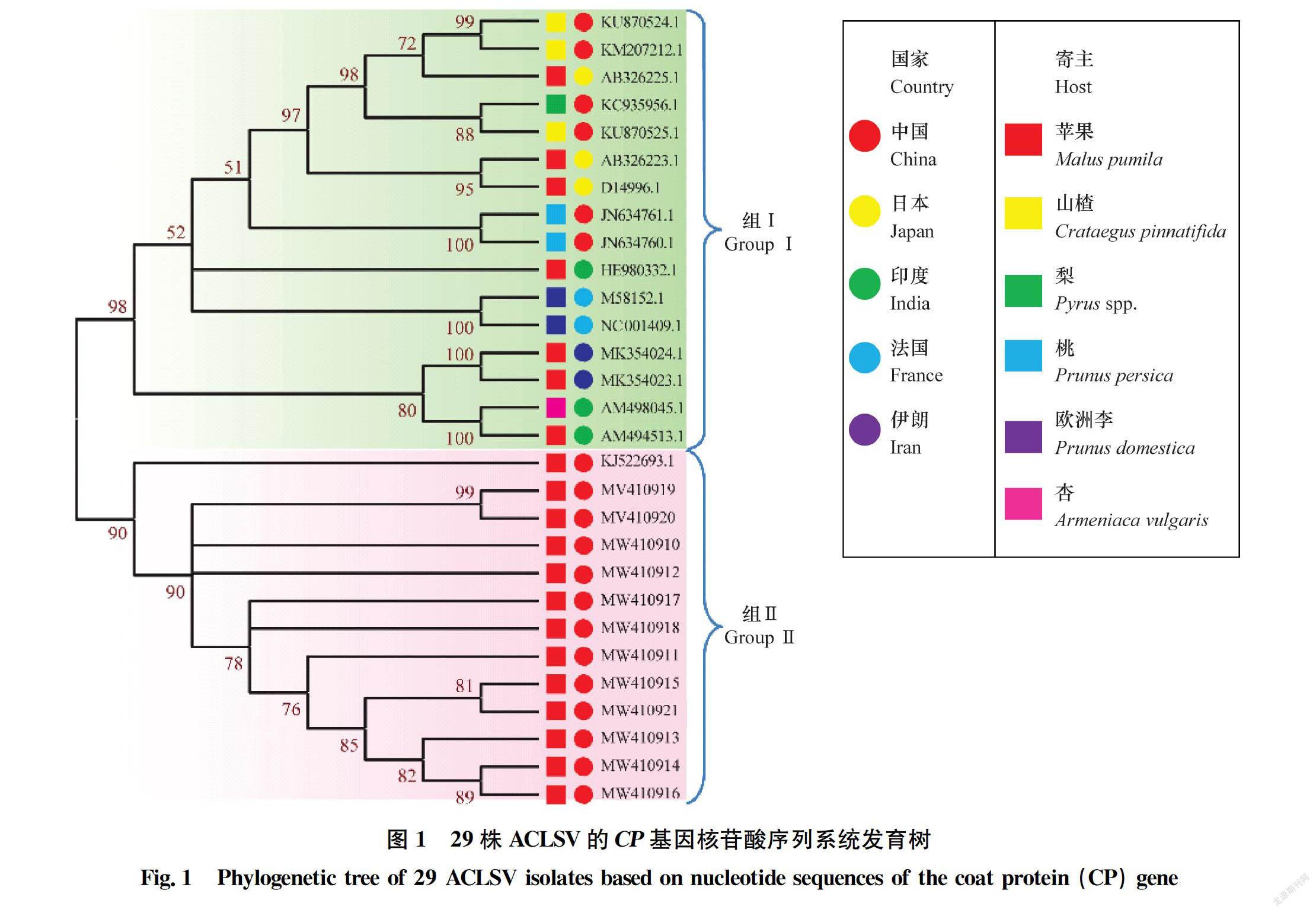
将来自NCBI数据库的17个分离物与本研究分离的12个分离物,共计29个ACLSV分离物的CP基因进行了系统发育和遗传变异分析。结果显示,29个ACLSV分离物被分为2组(图1)。其中,组Ⅰ包含16个分离物,这些分离物分别来自于5个国家的6个寄主。就国家而言,来自中国的分离物有6个,日本3个,印度3个,法国2个,伊朗2个。就寄主而言,分离自苹果的有7个,山楂3个,桃子2个,李子2个,梨1个,杏1个。组Ⅱ包含13个ACLSV分离物,均是从中国的苹果上分离获得。包括本研究分离的12个山西ACLSV分离物与来自山东的分离物(KJ522693.1)。
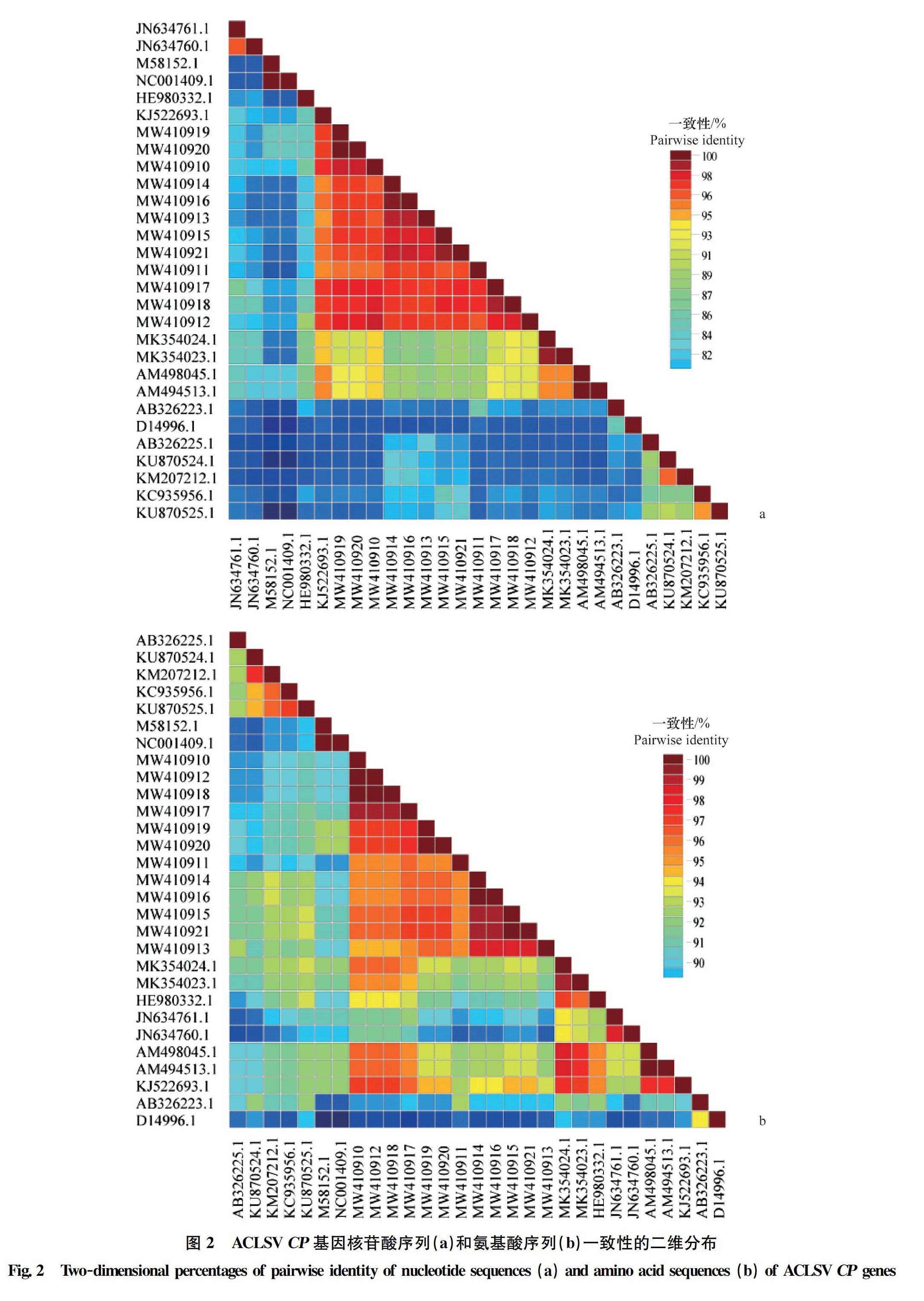
基于CP基因序列一致性分析表明,29个ACLSV分离物 CP基因核苷酸一致性为81.10%~100%,CP基因氨基酸一致性为88.60%~100%,这说明山西ACLSV中CP基因氨基酸在进化中具有较高保守性(图2)。本研究获得的山西分离物MW410910(Shanxi 1)与 KJ522693.1 的CP基因核苷酸一致性最高,为97.42%;MW410916(Shanxi 7)与D14996.1 序列的CP基因核苷酸一致性最低,为83.33%。其次,山西ACLSV分离物MW410910(Shanxi 1)、MW410912(Shanxi 3)和MW410918(Shanxi 9)与 KJ522693.1的氨基酸一致性最高,为97.93%;MW410917(Shanxi 8)与D14996.1序列的氨基酸一致性最低,为89.12%。
2.3 山西 ACLSV种群特征与变异情况
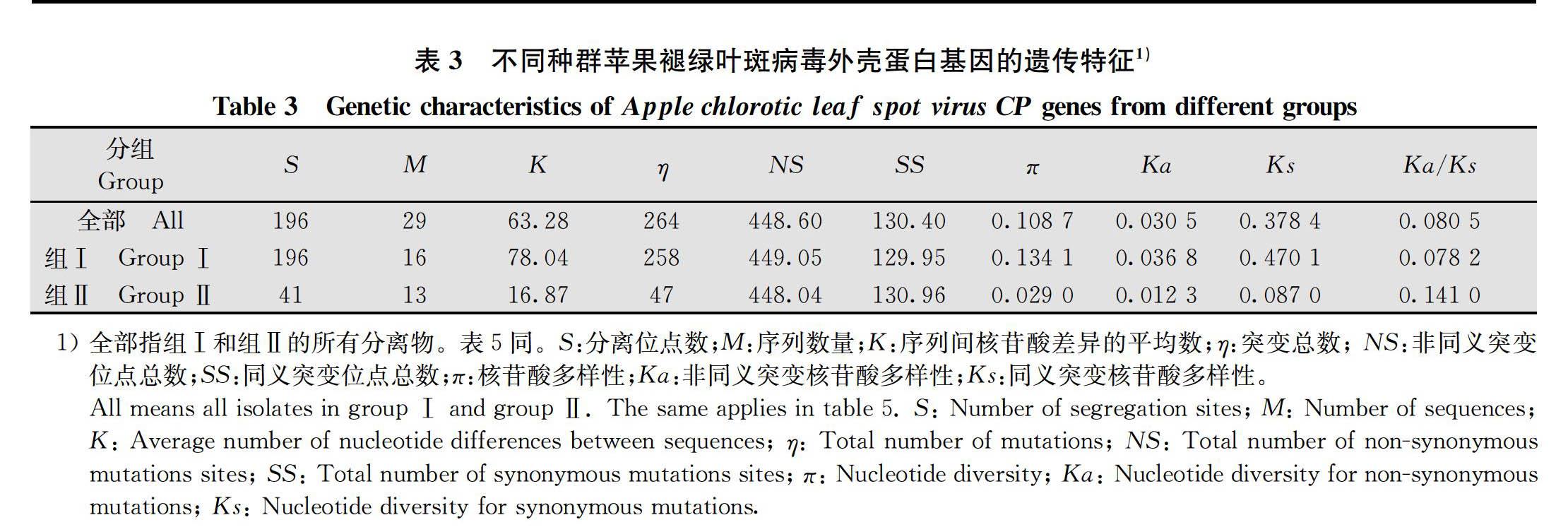
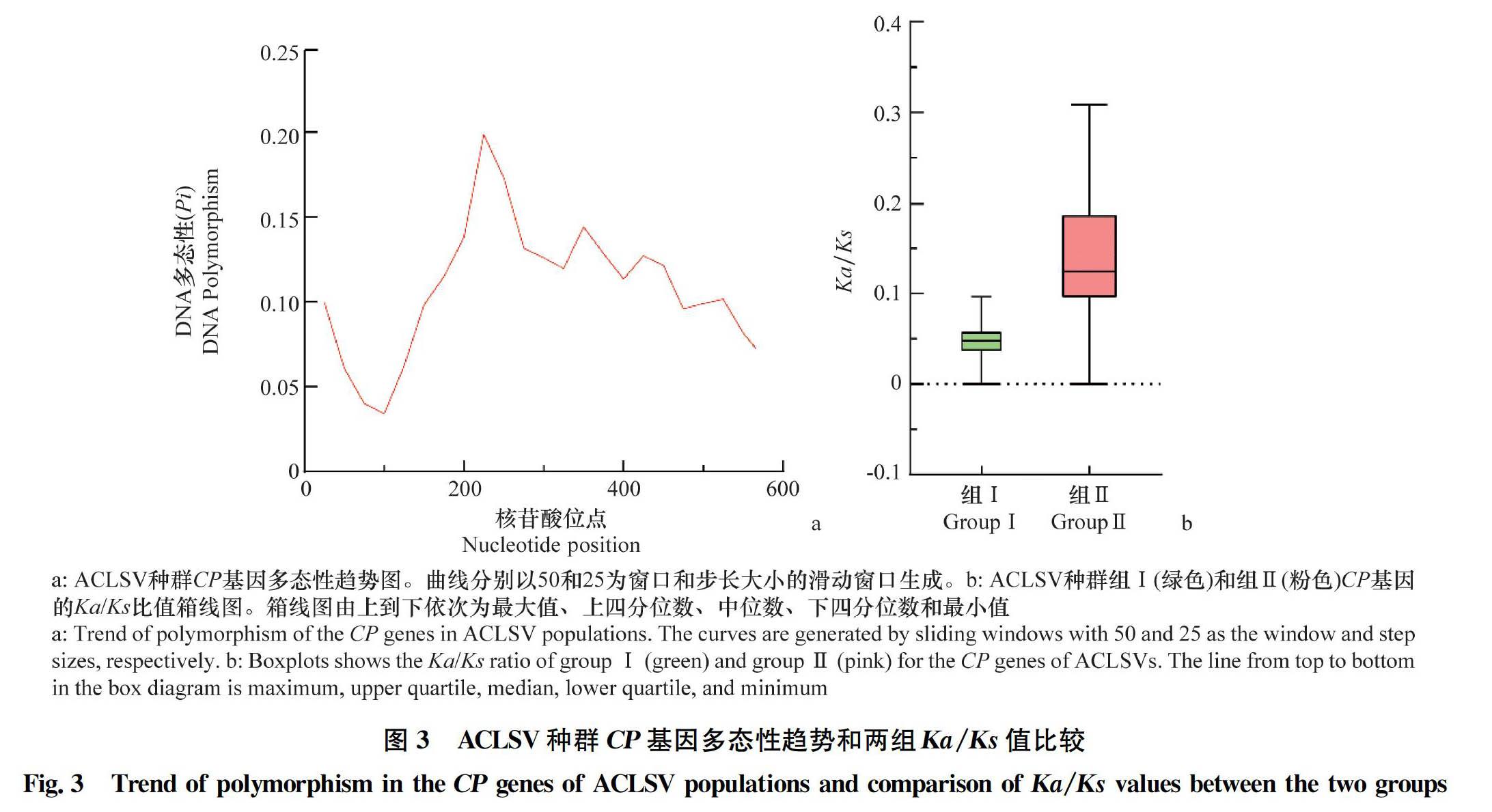
将来自于NCBI数据库的17个分离物与本研究分离的12个分离物 共29个ACLSV 分离物按照系统发育关系分为2个组,并使用DnaSP 5.10.01软件对ACLSV的种群特征进行分析。结果显示,组Ⅰ的突变总数(η=258)、分离位点数(S=196)、序列间核苷酸差异的平均数(K=78.04)和核苷酸多样性(π=0.134 1)都是最高的(表3)。随后,对ACLSV CP基因的多态性进一步分析,结果显示CP基因在1-50、201-250、326-375、401-450和501-550位点的多态性水平较高,对应Pi值分别为0.099 8、0.198 4、0.144 1、0.127 0、0.059和0.101 5(圖4a)。
2.4 山西ACLSV种群重组分析
重组是病毒遗传变异的一个重要来源。为了研究重组在ACLSV种群进化中的作用,利用重组检测软件RDP 4.31对来自本研究的12个分离物和来自GenBank的 17个分离物的CP基因进行了重组事件检测。检测结果显示,在29个ACLSV分离物中均未发现明显的重组事件。
为了进一步分析ACLSV CP基因的变异和选择压,本研究计算了29个ACLSV分离物非同义突变位点和同义突变位点的比值(Ka/Ks)(表3)。结果显示,ACLSV两个系统发育分组的非同义突变频率/同义突变频率的比值(Ka/Ks)均小于1,表明ACLSV 的CP基因进化主要受负向选择。进一步对2个进化枝的选择压差异进行分析,组Ⅰ和Ⅱ的Ka/Ks平均值分别为0.078 2和0.141 0(表3)。此外,组Ⅰ和组Ⅱ选择压的两两分析结果表明,组Ⅰ和组Ⅱ的Ka/Ks值分布存在差异(图3b)。虽然ACLSV两个组的Ka/Ks值均小于1,但是组Ⅱ的Ka/Ks值明显高于组Ⅰ,表明组Ⅰ比组Ⅱ受到更强的负向选择。
2.5 山西ACLSV的种群的差异检验和基因流分析
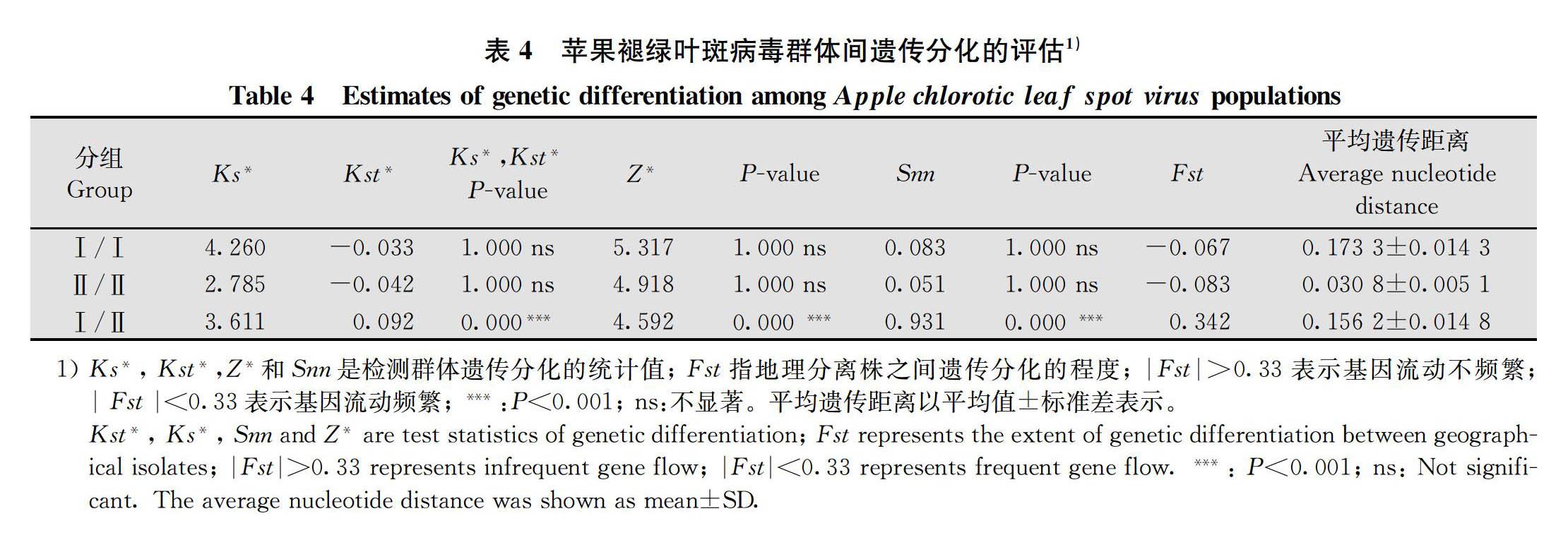
本研究评估了ACLSV种群组内与组间的遗传分化程度。根据Ks*, Kst*, Z* 和 Snn的P值,我们发现组Ⅰ与组Ⅱ在遗传分化上存在显著差异,而组Ⅰ内和组Ⅱ内分离物之间没有显著差异。同时我们可以看到在组Ⅰ内和组Ⅱ内的|Fst|均小于0.33,组Ⅰ与组Ⅱ组间的|Fst|大于0.33。说明ACLSV分离物在组内的基因流动非常频繁,组间的基因流动很少。进一步计算了组内与组间的遗传距离,结果显示,组Ⅰ内和组Ⅱ内分离物间的遗传距离分别为(0.173 3±0.014 3)和(0.030 8±0.005 1),而组Ⅰ与组Ⅱ间的遗传距离为(0.156 2±0.014 8)(表4)。
2.6 山西ACLSV中性检验和群体动态
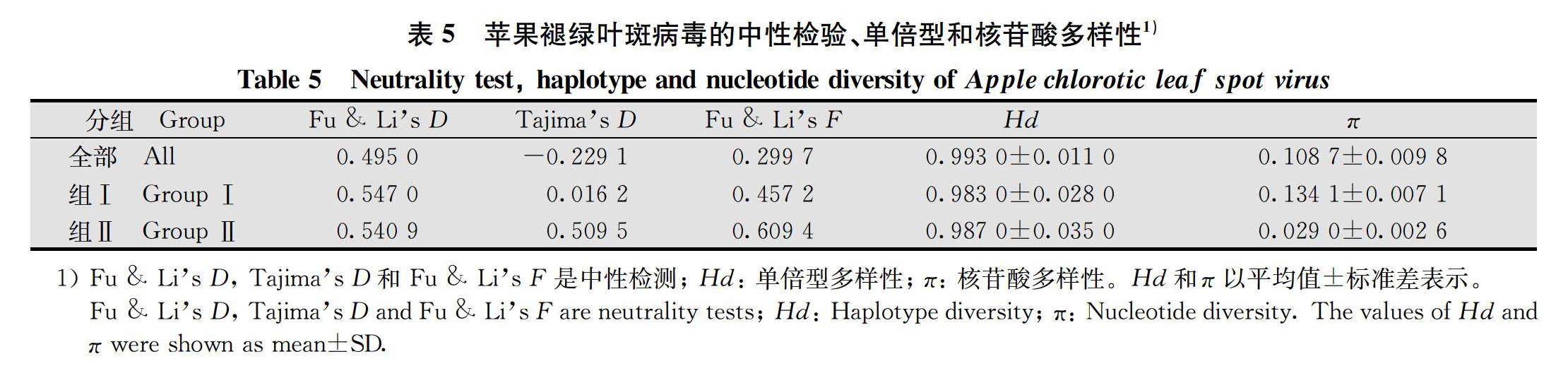
本研究基于Fu & Li’s D、Tajima’s D、Fu & Li’s F 统计检验值评估了29个ACLSV分离物CP基因的核苷酸多样性模式(表5)。研究结果显示,ACLSV组Ⅰ和组Ⅱ的Tajima’s D、Fu & Li’s D、Fu & Li’s F均为正值,但这些值均没有显著差异,这可能是由于种群瓶颈结构或/和平衡选择造成的。此外,单倍型多样性(Hd=0.987 0)高和核苷酸多样性(π=0.029 0)低均在组Ⅱ中出现。综上结果表明,山西ACLSV种群多样性可能是由于平衡选择造成的。
3 结论与讨论
苹果褪绿叶斑病毒是侵染苹果的主要潜隐性单链RNA病毒,严重威胁苹果产业的健康发展。本研究发现,采样的山西9个产区ACLSV的发病率超过50%,给果农造成巨大的经济损失。此外, ACLSV的寄主范围广,除可以侵染苹果外,还可侵染梨、桃、李、杏、樱桃等多种果树,对果树的生长、产量和质量均造成严重的影响[29]。ACLSV在不同宿主和环境中会发生突变以适应不同寄主,而高突变率加重了ACLSV危害[30]。本研究通过对山西及GenBank中登记的ACLSV分离物CP基因核苷酸序列、氨基酸序列、种群多样性和种群结构进行分析,基本明确了山西12个苹果产区的ACLSV存在变异,种群多样性较高,是危害山西苹果优产、高产的重要因素,有效预防ACLSV已经成为山西苹果产区亟待解决的问题。
山西苹果ACLSV种群是较为稳定的类群。ACLSV分离物系统发育分析和核酸序列一致性分析显示,来自中国山西苹果的分离物单独分为一组;其他的分离物(如中国山楂、中国桃子、日本苹果和印度苹果等)分为一组。这表明ACLSV分离株的遗传多样性主要是由地理和寄主因素共同决定的,該结果与2015年李科等报道的山东苹果分离物的聚类与地理或寄主并无明显关系的结果相反[18],这可能是由于本研究仅是基于CP基因分析,ACLSV遗传多样性是否与地理或寄主因素相关还需进一步验证。同时我们发现中国山西苹果的ACLSV种群与其他种群之间的遗传多样性存在明显差异,其种群内的突变数和核苷酸多样性都是最小,说明山西苹果的ACLSV种群相比其他已知的ACLSV分离物多样性较低。结合前人研究结果ACLSV在自然环境存在较大的由环境因子、地理位置、寄主等引起的负向选择压力[31]。可能是这种负选择压力影响了山西苹果ACLSV种群的CP基因变异,同时,不同ACLSV种群CP基因的Ka/Ks值表现出高度多样性,这些都证明ACLSV种群可能经历了不同的压力。本研究中来自山西苹果的ACLSV种群受到负选择压远小于组Ⅰ,说明该种群与其他ACLSV种群相比正趋于稳定。
地域和寄主的负向选择压力可能是山西苹果ACLSV种群变异的主要因素。有研究报道ACLSV种群之间存在显著的遗传差异和频繁的基因交流[6],而本研究发现,ACLSV分离株在组内的基因交流非常频繁,而在组Ⅰ与组Ⅱ之间很少出现基因交流。同时发现,组Ⅱ内的遗传距离小于组Ⅰ与组Ⅱ间的遗传距离,但有趣的是组Ⅰ内的遗传距离大于组Ⅰ与组Ⅱ间的遗传距离。这可能是由于组Ⅰ的分离物来自不同国家和寄主,而组Ⅱ的分离物均来自中国苹果。不同的宿主、环境和载体都可能对ACLSV产生选择压力,在强大的选择压力下ACLSV能够非常快速地进化,从而提高其遗传多样性。通过对山西ACLSV群体进行中性检验发现山西苹果ACLSV种群可能发生了平衡选择或种群收缩[32]。 以上结果进一步证实中国山西苹果的ACLSV种群与其他种群之间存在明显的遗传差异,同时山西苹果ACLSV种群是非常保守的。ACLSV作为单链RNA病毒,其RNA聚合酶缺乏DNA聚合酶的校对能力[33]。
参考文献
[1] 刘凤之, 王海波, 胡成志. 我国主要果树产业现状及“十四五”发展对策[J].中国果树, 2021(1): 15.
[2] 郑国富. 我国苹果出口贸易发展的特征、问题与升级战略[J].中国果树, 2021(6): 8992.
[3] 王金政, 毛志泉, 丛佩华, 等. 新中国果树科学研究70年——苹果[J]. 果树学报, 2019, 36(10): 12551263.
[4] 王树桐, 王亚南, 曹克强. 近年我国重要苹果病害发生概况及研究进展[J]. 植物保护, 2018, 44(5): 1325.
[5] 姚润东, 史文森, 孙吴润泽, 等. 中国西南主要苹果产区潜隐性病毒分子鉴定[J]. 四川大学学报(自然科学版), 2019, 56(2): 357362.
[6] CHEN Wei, JING Xiaoya, SUN Liuqing, et al. Genome cloning and genetic diversity of Apple chlorotic leaf spot virus[J]. Indian Journal of Biochemistry & Biophysics, 2019, 56: 514519.
[7] 李保华, 王彩霞, 董向丽. 我国苹果主要病害研究进展与病害防治中的问题[J]. 植物保护, 2013, 39(5): 4654.
[8] ABTAHI F, SHAMS-BAKHSH M, SAFAIE N, et al. Incidence and genetic diversity of apple chlorotic leaf spot virus in Iran [J]. Journal of Plant Pathology, 2019, 101(3): 513519.
[9] 秦子禹, 孙建设, 王娜, 等. 苹果茎痘病毒TaqMan探针实时荧光定量RT-PCR检测方法的建立[J]. 园艺学报, 2015, 42(7): 14001408.
[10]MA Xiaofang, HONG Ni, MOFFETT P, et al. Genetic diversity and evolution of Apple stem pitting virus isolates from pear in China [J]. Canadian Journal of Plant Pathology, 2016, 38(2): 218230.
[11]陈雅寒, 孙平平, 马强, 等. 东北冷寒产区苹果褪绿叶斑病毒检测及其分子多样性分析[J]. 园艺学报, 2019, 46(12): 23972405.
[12]BRAKTA A, THAKUR P D, HANDA A. First report of apple top working disease caused by viruses (Apple stem grooving virus, Apple chlorotic leaf spot virus, and Apple stem pitting virus) in apple in India [J]. Plant Disease, 2013, 97(7): 1001.
[13]CHEN Shanyi, ZHOU Ying, YE Ting, et al. Genetic variation analysis of apple chlorotic leaf spot virus coat protein reveals a new phylogenetic type and two recombinants in China[J]. Archives of Virology, 2014, 159(6): 14311438.
[14]NICKEL O, SILVA F N, FAJARDO T V M, et al. Characterization and genetic variability of coat protein genes of Apple chlorotic leaf spot virus isolates from southern Brazil[J]. Tropical Plant Pathology, 2017, 43(2): 109116.
[15]WANG M, DAI H. First report of apple chlorotic leaf spot virus in hawthorn in China[J]. Plant Disease, 2015, 99(1): 164.
[16]URBANOVICH O. Molecular variability of Apple chlorotic leaf spot virus isolated in Belarus [J]. Environmental and Experimental Biology, 2016, 14(3): 121126.
[17]KESHAVARZ T, HAJNAJARI H. Status of the Apple chlorotic leaf spot virus infection in native and imported apple tree cultivars in the national collection of Kamalshahr horticulture [J]. Journal of Plant Protection, 2019, 33(4): 387396.
[18]李科, 時洪伟, 荆陈沉, 等. ACLSV山东苹果分离物基因重组及CP序列多样性分析[J]. 中国农业科学, 2015, 48(14): 28572867.
[19]RANA T, CHANDEL V, KUMAR Y, et al. Molecular variability analyses of Apple chlorotic leaf spot virus capsid protein[J]. Journal of Biosciences, 2010, 35(4): 605615.
[20]HE Zhen, YASAKA R, LI Wenfeng, et al. Genetic structure of populations of sugarcane streak mosaic virus in China: comparison with the populations in India[J]. Virus Research, 2016, 211: 103116.
[21]张虎, 景晓雅, 孙柳清, 等. 玉米矮花叶病毒分离物基因组克隆及多样性分析[J]. 中国农业科技导报, 2019. 21(9): 3643.
[22]HOSSEINI H, MEHRVAR M, ZAKIAGHL M, et al. Comparative genetic diversity of potato virus Y populations based on coat protein gene [J]. Acta Virologica, 2017, 61(2): 161174.
[23]XIE Xiansheng, CHEN Wei, FU Qiang, et al. Molecular variability and distribution of Sugarcane Mosaic Virus in Shanxi, China [J/OL]. PLoS ONE, 2016, 11(3): e0151549. DOI: 10.1371/journal.pone.0151549.
[24]DALMON A, DESBIEZ C, COULON M, et al. Evidence for positive selection and recombination hotspots in Deformed wing virus (DWV) [J/OL]. Scientific Reports, 2017, 7: 41045. DOI: 10.1038/srep41045.
[25]WANG Yanan, WU Beilei, BORTH W B, et al. Molecular characterization and distribution of two strains of Dasheen mosaic virus on taro in Hawaii [J]. Plant Disease, 2017, 101(12): 19801989.
[26]SAMIEI A, MEHRVAR M, ZAKIAGHL M, et al. Distribution and phylogenetic analysis of the 3′UTR and coat protein gene of Iranian Beet black scorch virus[J]. Journal of Plant Diseases and Protection, 2019, 126(6): 535542.
[27]MURPHY J F, MORAWO T. Comparative evaluation of disease induced by three strains of Tobacco etch virus in Capsicum annuum L.[J]. Plant Disease, 2016, 101(1): 217223.
[28]LI Xiangdong, ZHU Tiansheng, YIN Xiao, et al. The genetic structure of Turnip mosaic virus population reveals the rapid expansion of a new emergent lineage in China [J/OL]. Virology Journal, 2017, 14(1): 165. DOI: 10.1186/s12985-017-0832-3
[29]秦子禹, 孙建设, 王娜, 等. 一种高效的苹果褪绿叶斑病毒TaqMan探针实时荧光定量RT-PCR检测方法[J]. 植物保护学报, 2015, 42(4): 7479.
[30]SONG Yansu, HONG Ni, WANG Liping, et al. Molecular and serological diversity in Apple chlorotic leaf spot virus from sand pear (Pyrus pyrifolia) in China [J]. European Journal of Plant Pathology, 2011, 130(2): 183196.
[31]GAO Fangluan, LIN Wuzhen, SHEN Jianguo, et al. Genetic diversity and molecular evolution of Arabis mosaic virus based on the CP gene sequence [J]. Archives of Virology, 2016, 161(4): 10471051.
[32]TORRICO A K, CELLI M G, CAFRUNE E E, et al. Genetic variability and recombination analysis of the coat protein gene of Strawberry mild yellow edge virus [J]. Australasian Plant Pathology, 2016, 45(4): 401409.
[33]XING Fei, ROBE B L, ZHANG Zhijia, et al. Genomic analysis, sequence diversity, and occurrence of Apple necrotic mosaic virus, a novel ilarvirus associated with mosaic disease of apple trees in China [J]. Plant Disease, 2018, 102(9): 18411847.
(責任编辑:杨明丽)
猜你喜欢
湖北农业科学(2017年22期)2017-12-25
中国医药导报(2017年30期)2017-11-16
成长·读写月刊(2017年10期)2017-10-15
江苏农业科学(2016年1期)2017-05-17
魅力中国(2016年19期)2017-04-18
电子技术与软件工程(2017年1期)2017-03-06
吉林农业(2016年12期)2017-01-06
中学生物学(2016年7期)2016-11-02
江苏农业科学(2014年10期)2014-11-22
江苏农业科学(2014年9期)2014-11-15
