
不同种源花榈木苗期生长及生理特性比较
2021-01-07 05:13金念情杨彬韦小丽肖龙海段如雁
广西植物 2021年12期
金念情 杨彬 韦小丽 肖龙海 段如雁


摘 要: 為了解不同种源花榈木在贵阳的生长特性和差异,该文通过对10个种源地花榈木进行育苗试验,测定其两年生实生苗的苗高、地径、生物量、叶片光合参数、光合色素、硝酸还原酶活性、硝态氮含量和根系活力,并进行差异性分析。结果表明:(1)10个种源花榈木净光合速率、气孔导度、胞间CO2浓度、蒸腾速率和水分利用效率差异显著(P<0.05),表明不同种源花榈木光合特性及光能利用效率具有较大差异,浙江杭州和浙江永康花榈木是具有较高光合生长潜力的种源。(2)种源间的叶绿素含量、硝酸还原酶、硝态氮、根系活力存在显著差异,福建建瓯种源的叶绿素a、叶绿素b含量和叶绿素总量最高,能够将光合原初反应过程中积蓄的光能进行高效地传递,促进碳的同化;贵州花溪种源硝酸还原酶活性最大,硝态氮含量最高,对氮元素的利用能力较强,能够促进植物蛋白质、氨基酸和叶绿素等的合成;贵州望谟种源根系活力最大,吸收养分的能力强。(3)各种源间苗高、地径和生物量的分配存在显著差异,浙江杭州种源的植株枝叶繁茂、根系发达,生长表现好,安徽黄山种源的植株矮小,生长表现较差;浙江杭州种源将生物量更多分配在根和叶,提高其根系吸收养分和叶片获取光能的能力,安徽黄山种源总体生物量积累最少,长势最差。(4)通过主成分分析法对各种源的花榈木适应性进行综合评价,结果显示浙江杭州种源>贵州黎平种源>浙江永康种源>贵州望谟种源>福建建瓯种源>贵州凯里种源>贵州石阡种源>贵州花溪种源>贵州平塘种源>安徽黄山种源。综上结果表明,浙江杭州、贵州黎平和浙江永康种源花榈木对贵阳地区立地环境具有较强的适应能力和生长潜力。
关键词: 花榈木, 种源, 生长差异, 生理, 主成分分析
中图分类号: Q945
文献标识码: A
文章编号: 1000-3142(2021)12-2051-10
收稿日期: 2020-09-22
基金项目: 国家自然科学基金(31460193);贵州省高层次创新人才培养计划项目(黔科合平台人才 [2016]5661);中央财政林业科技推广示范项目([2017]TG01 号) [Supported by the National Natural Science Foundation of China (31460193); Guizhou High Level Innovative Talents Training Program (Qiankehe Platform Talents [2016] 5661); Forestry Science and Technology Promotion Demonstration Project of Central Finance ( [2017] TG01)]。
作者简介: 金念情(1996-),硕士研究生,主要从事森林培育研究,(E-mail)JNQ19960823@163.com。
通信作者: 韦小丽,教授,博士研究生导师,主要从事森林培育研究,(E-mail)gdwxl-69@126.com。
Comparison of growth and physiological characteristics of
Ormosia henryi seedlings from different provenances
JIN Nianqing1, YANG Bin2, WEI Xiaoli1*, XIAO Longhai1, DAUN Ruyan3
( 1. College of Forestry, Guizhou University, Guiyang 550025, China; 2. Guizhou Forestry Survey and Design
Co. LTD, Guiyang 550001, China; 3. Guizhou Institute of Biology, Guiyang 550025, China )
Abstract: In order to understand the growth characteristics and differences of different Ormosia henryi provenances in Guiyang, we collected seeds from ten different provenances of O. henryi and conducted seedling experiments. We compared differences in seedling height, root collar diameter, biomass, leaf photosynthetic parameters, photosynthetic pigment, activity of nitrate reductase, content of nitrate nitrogen and the root activity of the biannual seedlings, and then analyzed the differences in ten provenances of O. henryi. The results were as follows: (1) There were significant differences in the net photosynthetic rate (Pn), stomatal conductance (Gs), intercellular carbon dioxide concentration (Ci), transpiration rate (Tr) and water use efficiency (WUE) among ten O. henryi provenances (P<0.05), indicating that different O. henryi provenances showed significant differences in photosynthetic characteristics and light use efficiency. Plants from Hangzhou and Yongkang of Zhejiang had the higher photosynthetic growth potential. (2) There were significant differences in chlorophyll content, nitrate reductase, nitrate nitrogen and root activity among provenances. The contents of chlorophyll a and chlorophyll b and total content of chlorophyll were the highest in Jian'ou of Fujian provenance. These plants could efficiently transfer the accumulated light energy during the initial reaction of photosynthesis and promote carbon assimilation. Huaxi of Guizhou provenance had the highest nitrate reductase activity, the highest nitrate nitrogen content, and strong utilization ability of nitrogen elements, which could promote the synthesis of plant protein, amino acid and chlorophyll. The root system from plants in the Wangmo of Guizhou provenance had the greatest activity and the ability to absorb nutrients. (3) There were significant differences in the distribution of seedling height, ground diameter and biomass among all provenances. The plants from Hangzhou of Zhejiang provenance had luxuriant branches and leaves, developed roots and good growth performance, while the plants from Huangshan of Anhui provenance were short and had poor growth performance. Hangzhou of Zhejiang provenance distributed more biomass in roots and leaves to increase the ability of roots to absorb nutrients and leaves to acquire light energy. Huangshan of Anhui provenance accumulated the least biomass and had the worst growth. (4) The adaptability of ten provenances of O. henryi was comprehensively evaluated by principal component analysis. The results showed that Hangzhou of Zhejiang provenance > Liping of Guizhou provenance > Yongkang of Zhejiang provenance > Wangmo of Guizhou provenance > Jian’ou of Fujian provenance > Kaili of Guizhou provenance > Shiqian of Guizhou provenance > Hauxi of Guizhou provenance > Pingtang of Guizhou provenance > Huangshan of Anhui provenance. In conclusion, Plants from Hangzhou of Zhejiang, Liping of Guizhou and Yongkang of Zhejiang O. henryi provenances had high potential for adaptation and growth potential in Guiyang.
Key words: Ormosia henryi, provenance, growth difference, physiological, principal component analysis
种源试验是研究植物种群变异与环境因子之间的相关性,是进行遗传改良和引种栽培的重要手段(黄国伟等,2017)。同一树种在不同的地理环境下,因为缺乏基因交流,会在长期对环境的适应下产生不同的性状差异(Gu,2010),且林木的育种周期较长,所以对林木苗期的试验成为引种优良种源的初筛方法。种源试验在对麻栎(Quercus acutissima)(薛美玲等,2019)、香椿(Toona sinensis)(周祥斌等,2016)、棕榈(Trachycarpus fortunei)(王婷婷等,2019)等植物的研究中取得了一定成果并运用到生产实践中。
花榈木(Ormosia henryi)又名花梨木,蝶形花科红豆树属,是国家Ⅱ级重点保护野生树种和珍稀濒危树种,在我国长江以南大部分省份均有分布,生长在海拔为100~1 300 m的地方(孟宪帅和韦小丽,2011)。花榈木的木材坚韧、纹理清晰细密、材质紧密优良、削面光滑美观,是制造高档家具和雕刻的珍贵用材树种,且种子色泽鲜艳可做工艺品及装饰品(郑万钧,1985)。目前对花榈木的研究多集中在群落特征(王小东等,2018;刘鹏等,2017;孟宪帅和韦小丽,2011)、菌根接种(段如雁等,2019)、种子休眠及破除方法(邓兆等,2011)、虫害防控(徐芳玲等,2015)、育苗基质筛选、栽培与快繁(段如雁等,2017)等方面。由于长期的生殖隔离和周围环境因子的共同作用,使同一树种的不同个体和群体间形态特征和生长存在显著差异,不同地理种源的物种在不同区域的生长表现不同(李阳等,2020;Lestari et al.,2011)。有研究发现,不同种源香椿的光合特性和叶绿素含量存在显著差异(周祥斌等,2016),而硝酸还原酶、硝态氮在不同种源红皮云杉之间差异显著,并对植物生长起重要作用(王秋玉等,2003)。根系活力体现在根系吸收、利用、转化和还原能力,是体现根系功能的综合性指标。光合能力的大小、叶绿素含量、硝酸还原酶、硝态氮的含量和根系活力是影响不同种源植物生产力大小的重要因素。因此,各地需通过种源试验来选择适合当地自然环境的地理种源。目前,对于不同种源花榈木苗期生长变异的研究还未见报道。本文以浙江、福建、安徽和贵州花榈木分布区的10个不同地理种源种子培育的两年生花榈木幼苗为研究对象,对各种源花榈木苗期生长及光合生理特性进行测定,分析生理和生长的差异性,探索花榈木苗期生物学特性与地理空间的关系,以期为选择本地区优良种源提供苗期分析,同时为花榈木种源的遗传改良工作提供基础数据。
1 研究方法
1.1 试验地概况
试验地位于贵州省贵阳市花溪区贵州大学林学院实验苗圃,属黔中山原地貌,地理位置为106°40′ E、26°25′ N。年平均降雨量1 129 mm,年相对湿度79%,年生长期271 d,绝对最高温度39.5 ℃,最低气温-9.5 ℃,年平均温度15.8 ℃,海拔1 100 m,属于亚热带高原湿润季风气候,试验地光照充足,排水良好。
1.2 试验材料
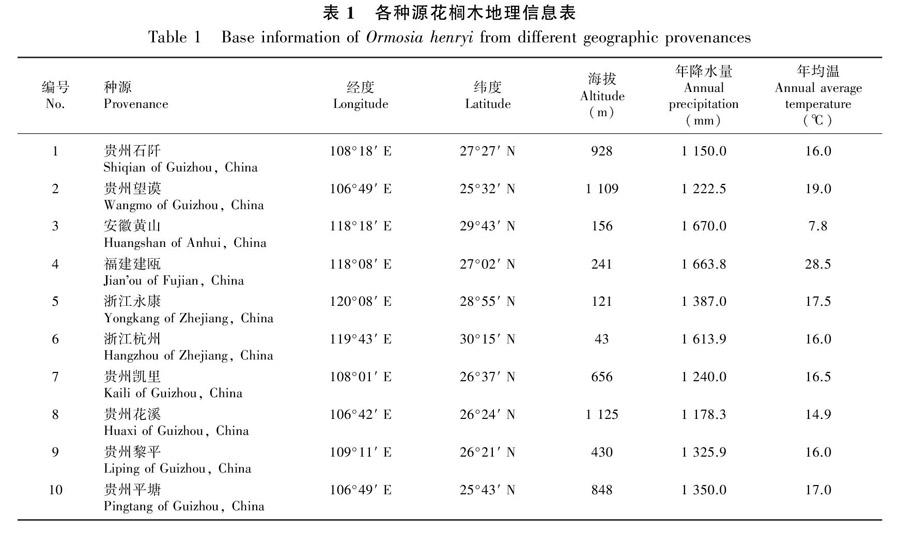
试验材料为两年生花榈木实生苗,种子分别来源于贵州、浙江、福建、安徽的10个不同地区(表1),育苗的基质相同,苗木管理方式一致。
1.3 试验材料培育
试验于2015年4月开始播种,对10个种源花榈木种子进行浸种催芽后,在试验地内用容器袋播种育苗。育苗基質为V珍珠岩∶V蛭石∶V泥炭=1∶1∶2混合基质,在每立方米基质中添加20 g多菌灵和2%过磷酸钙2 g,搅拌均匀后将种子穴播于规格为12 cm×15 cm的无纺布容器袋中。试验采用完全随机化设计,每个种源的种子播种300袋。在幼苗出土后搭盖遮荫网,并做好浇水、除草、施肥及病虫害防治等工作,苗木管理方式一致。
1.4 指标测定方法
1.4.1 生长指标的测定 2016年11月,待其生长结束后,每个种源的苗木随机选取30株挂牌编号,测定其苗高、地径。
1.4.2 生物量的测定 在2016年待苗木停止生长后对花榈木幼苗进行生物量的测定,每个种源的幼苗随机选取9株。将每株幼苗从育苗袋中取出,用清水洗去根团基质,保持根系完整,用吸水纸吸干后置于105 ℃烘箱中烘干至恒重后,称其根、茎、叶干重。
1.4.3 生理指标的测定 于2016年8月随机选择每个种源的5株健康植株作为重复,选择典型晴天9:00—11:00测定生理指标。设置光强 1 600 μmol·m-2·s-1、叶室温度 30 ℃、空气流量500 μmol·s-1、CO2浓度400 μmol·mol-1,采用LI-6400便捷式光合仪测定成熟叶片的光合指标参数,之后取样测定植株的叶绿素含量、硝酸还原酶、硝态氮和根系活力。水分利用效率计算公式为WUE=Pn/Tr。采用张治安等(2004)的方法对各项指标进行测定。采用乙醇-丙酮混合液提取法测定叶绿素含量;采用活体法测定硝酸还原酶活性;采用水杨酸法测定硝态氮含量;采用TTC法测定根系活力。
1.5 数据计算及统计方法
采用Excel 2010进行数据的录入、整理和图表制作,用SPSS 18.0统计分析软件对试验数据进行单因素方差分析(One-way ANOVA),多重比较采用最小显著差数法(LSD法)。
2 结果与分析
2.1 不同地理种源花榈木生长特性比较
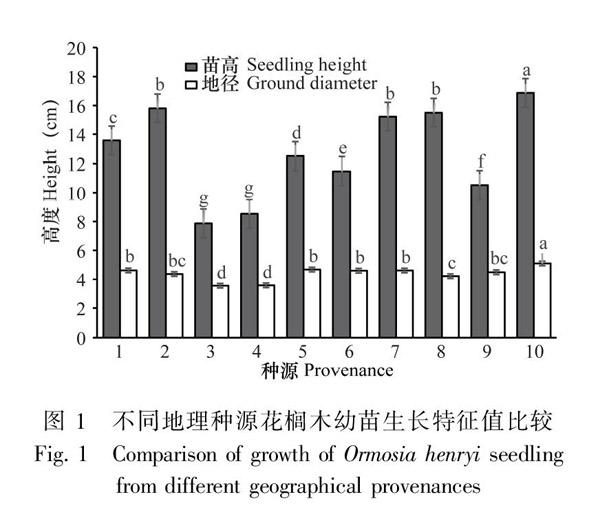
10个种源间花榈木苗高、地径表现出显著差异(P<0.05)。苗高的变异幅度为7.87~16.87 cm,最高的是贵州平塘,最矮的是安徽黄山;地径的变异幅度为3.54~5.1 mm,最粗的是贵州平塘,最细的是安徽黄山(图1)。
对不同种源花榈木苗高和地径进行方差分析,结果表明苗高和地径存在显著差异(P<0.05),不同种源花榈木对苗高和地径的生长有显著影响。总体来说,安徽黄山和福建建瓯的苗高和地径生长较慢,来自贵州平塘的生长表现最好。
2.2 不同地理种源花榈木苗期生物量比较
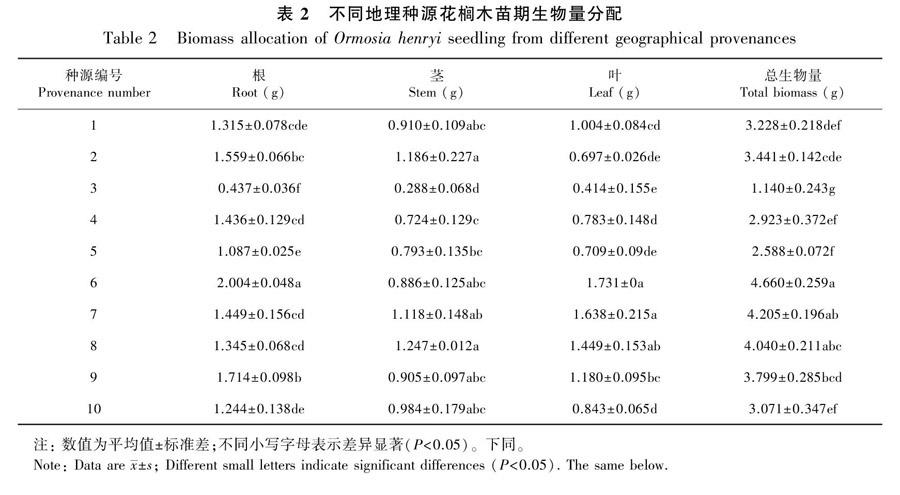
由表2可知,不同种源花榈木的根生物量的变异幅度为0.437~2.004 g,根生物量最大的是浙江杭州,最小的是安徽黄山;茎生物量的变异幅度为0.288~1.247 g,茎生物量最大的是贵州花溪,最小的是安徽黄山;叶生物量的变异幅度为0.414~1.731 g,叶生物量最大的是浙江杭州,最小的是安徽黄山; 总生物量的变异幅度为1.14~4.66 g, 总生物量最大的是浙江杭州,最小的是安徽黄山。
总体上,不同种源的花榈木各部位生物量存在较大差异,根、茎、叶和总生物量的最大值分别为最小值的4.59、4.33、4.18、4.09倍,说明不同种源的花榈木对环境的生长适应性不同,尤其是以安徽黄山和浙江永康种源的生物量积累较低,而浙江杭州和贵州花溪的生物量积累最好。
2.3 不同地理种源花榈木叶片叶绿素含量比较
由表3可知,不同种源花榈木的叶绿素a含量、叶绿素b含量、叶绿素总量和叶绿素a/叶绿素b均差异显著(P<0.05),其变异幅度分别为1.36~2.0 mg·g-1、0.51~1.29 mg·g-1、1.87~3.29 mg·g-1、1.68~2.68。其中,福建建瓯的花榈木叶绿素a含量、叶绿素b含量和叶绿素总量最高,贵州平塘的最低;叶绿素a/叶绿素b最大的是贵州平塘,最小的是福建建瓯。福建建瓯种源叶绿素a含量、叶绿素b含量和叶绿素总量分别是含量最低的贵州平塘种源的1.47、2.53和1.76倍,而叶绿素a/叶绿素b是贵州平塘的0.4倍左右,表明福建建瓯种源花榈木幼苗的耐荫性较贵州平塘和其他种源都高,不同种源对光的吸收能力不同,福建建瓯种源对红光有较强的吸收能力,光合作用最强,各种源对蓝光的吸收基本一致,没有显著差异(P>0.05)。
2.4 不同地理种源花榈木硝酸还原酶活性、硝态氮含量、根系活力比较
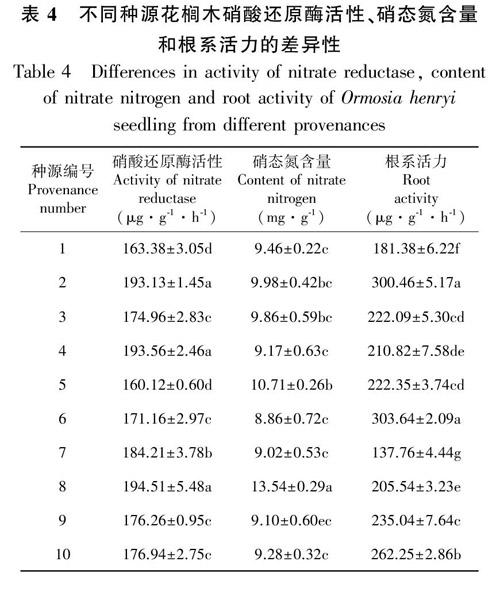
由表4可知,不同种源花榈木的硝酸还原酶活性的变异幅度为160.12~194.51 μg·g-1·h-1,最大的是贵州花溪,最小的是浙江永康;硝态氮含量的变异幅度为8.86~13.54 mg·g-1,最高的是贵州花溪,最低的是浙江杭州;根系活力的变异系数为137.76~303.64 μg·g-1·h-1,最高的是浙江杭州,最低的是贵州凯里。
各种源间花榈木的硝酸还原酶活性、硝态氮含量和根系活力均存在差异,而硝酸还原酶活性的大小和硝态氮含量的多少能反映出植物对氮元素的吸收和利用能力,根系活力则影响植物地上部分的营养和生长状况。说明贵州花溪的氮元素营养较高,有利于生长,而贵州凯里的根系活力较低,仅为根系活力最大的浙江杭州种源的0.45倍,远低于平均值,生长潜力较差。
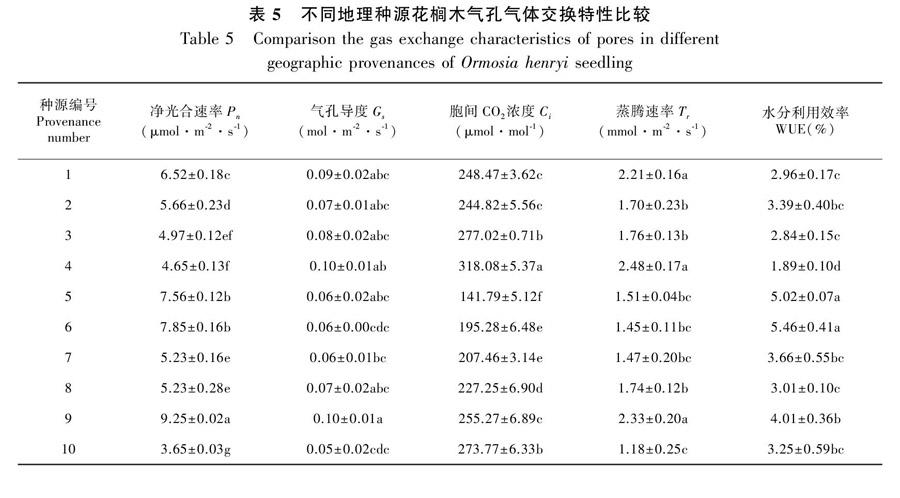
2.5 不同地理种源花榈木光合参数比较
由表5可知,10个种源花榈木叶片的气体交换参数净光合速率(Pn)、胞间CO2浓度(Ci)、蒸腾速率(Tr)及水分利用率(WUE)、气孔导度(Gs)在种源间的差异均达到显著水平(P<0.05),说明不同种源的花榈木幼苗的光合能力差异较大(P<0.05)。其中,Pn变幅在3.65~9.25 μmol·m-2·s-1之间,最大的是贵州黎平种源, 最小的是贵州平塘种源,仅为前者的0.39倍;Gs变幅在0.05~0.1 mol·m-2·s-1之间,最大的是贵州黎平和福建建瓯两个种源,是最小的贵州平塘种源的2倍;而Ci浓度变幅在141.79~318.08 μmol·mol-1之间,最小的是浙江永康种源,最大的是福建建瓯种源,是浙江永康种源的2.24倍;Tr变幅在1.18~2.48 mmol·m-2·s-1之间,最大和最小分别是福建建瓯种源和贵州平塘种源,相差达到1.3 mmol·m-2·s-1;WUE变幅在1.89%~5.46%之间,最高的是浙江杭州种源,最低的是福建建瓯种源,相差了3.57%。由此可见,各种源之间光能利用效率差异较大。总体上,贵州黎平和浙江杭州种源花榈木的光合效率较高,其净光合速率较高,且胞间二氧化碳摩尔分数较低,有较强的光合能力。
2.6 花榈木幼苗叶片气体交换参数之间的相关性分析
由表6可知,气孔导度(Gs)与胞间二氧化碳浓度(Ci)和蒸腾速率(Tr)均呈正相关关系,气孔导度(Gs)和蒸腾速率(Tr)呈极显著正相关(P<0.01),说明气孔导度越大,蒸腾速率越高。凈光合速率(Pn)与胞间二氧化碳浓度(Ci)、气孔导度(Gs)、蒸腾速率(Tr)不相关。水分利用效率(WUE)与净光合速率(Pn)呈正相关(P<0.05),与胞间CO2浓度呈极显著负相关(P<0.01),说明光合速率越大,水分利用率越高,胞间CO2浓度就越低。
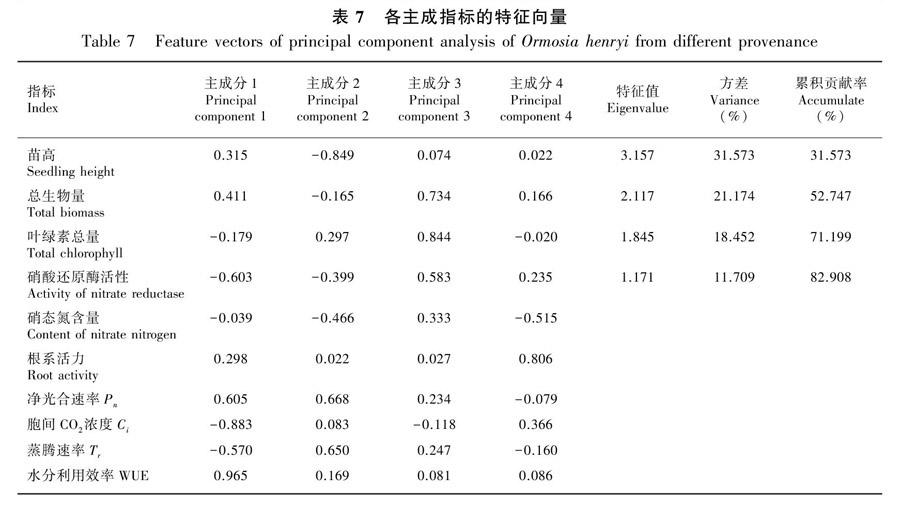
2.7 不同种源花榈木生长及生理主成分分析
通过主成分分析法,对测定的各项指标进行相关性综合评价,提取具有代表性的10个指标,分别为苗高、总生物量、叶绿素总量、硝酸还原酶活性、硝态氮含量、根系活力、净光合速率、蒸腾速率、胞间CO2浓度、水分利用效率。对不同种源花榈木的这10个指标进行主成分分析,提取特征值>1的4个主成分,其贡献率分别为31.57%、21.17%、18.45%、11.71%,累计贡献率为82.91%,可以进行分析。第一主成分中,水分利用效率的特征值达到0.965;从第二主成分来看,净光合速率和蒸腾速率特征值较大,分别为0.668和0.650;第三主成分叶绿素总量的特征值最大,为0.844;从第四主成分来看,根系活力的特征值为0.806,是第四主成分的主导因子(表7)。累积贡献率分别为31.57%、52.75%、71.20%和82.91%,累计贡献率增长较慢,说明花榈木的生长存在丰富的遗传多样性。
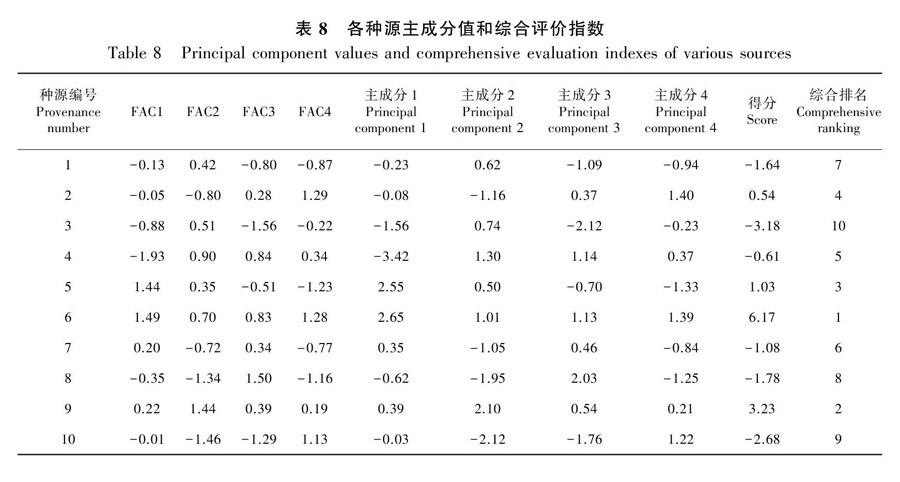
通过参考薛美玲(2019)的方法计算各种源花榈木的综合评价指数,计算得到10个种源花榈木的综合评价指数,结果见表8。具体表现为浙江杭州种源(6.17)>贵州黎平种源(3.23)>浙江永康种源(1.03)>贵州望谟种源(0.54)>福建建瓯种源(-0.61)>贵州凯里种源(-1.08)>贵州石阡种源(-1.64)>贵州花溪种源(-1.78)>贵州平塘种源(-2.68)>安徽黄山种源(-3.18),浙江杭州种源的综合性状最好,在试验地环境适应性最强,贵州黎平种源、浙江永康种源和贵州望谟种源表现优良,其余的种源表现较差。
3 讨论与结论
植物个体的差异受到其遗传资源和环境因子的共同影响。而同一树种分布在不同的地理区域内,由于光照、养分和水分的差异,会形成不同的生态习性和遗传特性,并在种子中遗传保存下来。因此,采集不同分布区内的种子在相同环境中培育出来,受遗传因素的影响,会表现出显著的生长差异(薛美玲等,2019)。叶片是植物进行光合作用的主要场所, 光合速率直接反映出光合作用的强弱,受温度、光照、水分等环境因素的影响,其气孔导度、蒸腾作用和水分利用效率会产生变化(张力文等,2012)。有研究表明,气孔作为叶片水气交换的通道,其开度直接影响叶片光合作用过程中碳的固定(Flexas et al.,2009),胞间二氧化碳浓度受气孔导度的影响较大,气孔导度越大,胞间二氧化碳浓度越高,越有利于碳的固定,但蒸腾过程中水分散失较大,水分利用效率低,不利于叶片水分的保持(薛美玲等,2019)。周祥斌等(2016)对不同种源香椿幼苗的研究认为,光合参数主要受遗传因子控制,可以作为分析香椿的遗传多样性和地理变异的主要性状。本研究对10个种源花榈木的光合参数进行分析,各光合参数间存在显著差异,其光合特征和适应能力均有所区别,这主要是由于遗传因素的影响,不同种源花榈木在相同环境下的光合响应不同,与本地种源相比,浙江杭州、永康种源的气孔开度较低,通过增加净光合速率、降低蒸腾速率来提高叶片水分利用效率,这可能是由于浙江杭州、永康海拔较低,相对湿度较大,因此该地花榈木通过减小气孔导度,降低蒸腾速率,减小叶片的水分散失,从而提高水分利用效率,这与栓皮栎黄龙林区种源通过较小的蒸腾速率、较大的水分利用效率来提高幼苗对环境的适应能力的策略相同(段宝利,2003),都具有较大的生长潜力;贵州黎平、石阡和望谟太阳辐射总量较花溪多,水分充足,花榈木光合能力较强,能夠促进苗木生物量的积累,这与谭长强等(2019)认为火力楠具有高光合速率就具有高光能利用效率,较高的净光合速率在一定程度上代表较高的生物量的观点一致。福建建瓯种源地温度较高,气孔导度较大,同时蒸腾作用较强,因此叶片水分散失较快,净光合速率低,水分利用效率低,光合储能较低;贵州平塘花榈木采种地地势较低,河谷湿度大,因此气孔导度较小,蒸腾速率慢,净光合速率较低;安徽黄山种源地温度低,净光合速率低, 对光能的吸收和转化能力较差。
叶绿素是植物进行光合作用的重要色素,起着接收和转化光能的作用,其浓度的高低在一定程度上决定着植株光合速率的快慢,从而影响植株干物质的积累,叶绿素的积累受环境因素影响极为显著(王小东等,2018)。本研究结果表明,浙江杭州种源的叶绿素总量较花溪种源的高,叶片光合作用积蓄的能量能够通过色素分子传递的效率更高,将活跃的化学能转化为稳定的碳水化合物的能力更强;福建建瓯种源的叶绿素a含量和叶绿素总量最高,能够加强光能的传递,但由于原初反应过程的效率不高,因此积蓄的能量较少,导致碳收益较低,这一结果与姚甲宝等(2018)的研究结果相似。硝酸还原酶能够将植物体内的氮素进行同化,转化为植物能够进行吸收利用的硝态氮,为植物生长中物质的合成提供氨基酸和蛋白质,促进植物的生长(孙敏红等,2013),根系则是植物吸收土壤中各种元素的组织,其活力的大小直接决定了植物获取营养的能力(王秋玉等,2003)。冯丽贞和刘爱琴(1998)对杉木的研究结果表明,不同种源杉木的生长速率与硝酸还原酶活性呈正相关,硝酸还原酶活性在一定程度上可以代表杉木的生长速率,本研究结果也表现出相同规律,花溪种源的硝酸还原酶活性和硝态氮含量均最高,且茎生物量最多。夏莘等(2018)认为,浙江松阳种源的鹅掌楸在生物量分配时,将更多的资源分配到根和叶,能够扩大根吸收面积和叶片光合面积,以获取更多养分和能量,有利于植株生长,本研究中浙江杭州种源也是采取这种策略,其根系活力是贵州花溪种源的1.48倍,总生物量是花溪种源的1.15倍,说明浙江杭州种源对营养的吸收和转化能力高于花溪种源,对环境的适应能力更强。苗高和地径作为苗木生长状况最直观的指标,对优良种源的早期选择有重要意义。孙银祥等(1999)对不同种源的樟树研究结果认为,樟树不同种源间苗期生物量具有显著差异,苗高、地径、地上部分生物量之间存在显著的表型差异和遗传影响,地径与地上部分生物量紧密联系。本研究结果表明,不同种源间苗高、地径、生物量等生长性状存在显著差异,浙江杭州种源表现最好,黄山表现最差,本地种源优势不明显,导致这种差异的主要原因可能是在长期的地理隔离中,各种源受环境因素的影响,形成一定的基因型差异,更好地适应其分布区域地理环境,而在相同环境生长时,由于这种差异促使外地种源的生长表现甚至优于本地种源。植物在不同生境中长期的适应使其形成了不同光照获取策略和生活习性。
综上所述,为更科学合理地筛选出适合本地生长的种源,采用主成分分析法,将10个种源花榈木的生长情况进行综合判定,结果为浙江杭州种源>贵州黎平种源>浙江永康种源>贵州望谟种源>福建建瓯种源>贵州凯里种源>贵州石阡种源>贵州花溪种源>贵州平塘种源>安徽黄山种源,综合得出结论,浙江杭州种源在本地的生长表现较好,是引种的首选种源,贵州黎平、浙江永康和贵州望谟种源的花榈木表现也较本地种源优良,而安徽黄山种源生长最差,不适宜在本地进行引种栽培。
参考文献:
DENG Z, WEI XL, MENG XS, et al., 2011. A preliminary study on dormancy and germination of Ormosia henryi seeds [J]. Guizhou Agric Sci, 39(5): 69-72. [邓兆, 韦小丽, 孟宪帅, 等, 2011. 花榈木种子休眠和萌发的初步研究 [J]. 贵州农业科学, 39(5): 69-72.]
DUAN BL, 2003. Study on the water physio-ecological characteristics and the adaptation to water stress for four provenances of Quercus variabilis [D]. Yangling: Northwest A & F University. [段宝利, 2003. 不同种源栓皮栎水分生理生态特性及其对干旱胁迫适应 [D]. 杨凌: 西北农林科技大学.]
DUAN RY, 2019. Diversity of Rhizobium in Ormosia henryi prain and screening of excellent strains [D]. Guiyang: Guizhou University. [段如雁, 2019. 花榈木根瘤菌多样性及优良菌株筛选 [D]. 贵阳: 贵州大学]
DUAN RY, WEI XL, ZHANG ZD, et al., 2017. Study on formula fertilization for container seedlings of rare species Ormosia henryi [J]. J For Environ, 37(2): 225-230. [段如雁, 韦小丽, 张之栋, 等, 2017. 珍贵树种花榈木容器苗配方施肥试验 [J]. 森林与环境学报, 37(2): 225-230.]
FENG LZ, LIU AQ, 1998. Study on nitrate reeducates activity comparison of different provenance Chinese fir [J]. J Fujian Coll For, 18(4): 3-5. [馮丽贞, 刘爱琴, 1998. 不同种源杉木硝酸还原酶活力的比较研究 [J]. 福建林学院学报, 18(4): 3-5.]
FLEXAS J, BARN M, BOTA J, et al., 2009. Photosynthesis limitations during water stress acclimation and recovery in the drought-adapted Vitis hybrid Richter-110 (V. berlandieri × V. rupestris) [J]. J Exp Bot, 60(8): 2361-2377.
GU L, PALLARDY SG, TU K, et al., 2010. Reliable estimation of biochemical parameters from C3 leaf photosynthesis-intercellular carbon dioxide response curves [J]. Plant Cell Environ, 33(11): 1852-1874.
HUANG GW, PENG C, CHEN HL, et al., 2017. Comparison of seedling growth and photosynthetic characteristics of different provenances of Toona ciliata Roem. [J]. J NW For Univ, 32(2):123-129. [黄国伟, 彭婵, 陈慧玲, 等, 2017. 不同种源红椿苗期生长及光合特性比较 [J]. 西北林学院学报, 32(2): 123-129.]
LESTARI R, EBERT G, HUYSKENS-KEIL S, 2011. The effect of light and water supply on growth, net CO2 assimilation rate and mineral content of salak (Salacca zalacca (gaertn.) voss) seedlings [J]. Int J Biol, 3(3): 94.
LIU P, HE WC, HUANG XC, et al., 2017. Research status and conservation strategy of Ormosia henryi [J]. S Chin For Sci, 45(3): 45-48. [刘鹏, 何万存, 黄小春, 等, 2017. 花榈木研究现状及保护对策 [J]. 南方林业科学, 45(3): 45-48.]
LI Y, WEI JP, MA HY, 2020. Variations of phenotypic characteristics in Leymus chinensis among different provenances [J]. Acta Ecol Sin, 40(4): 1175-1183. [李阳, 魏继平, 马红媛, 2020. 不同种源羊草表型差异性 [J]. 生态学报, 40(4): 1175-1183.]
MENG XS, WEI XL, 2011. Life table and surival analysis on Ormosia wild endangered populations [J]. Seed, 30(7): 66-68. [孟宪帅, 韦小丽, 2011. 濒危植物花榈木野生种群生命表及生存分析 [J]. 种子, 30(7): 66-68.]
SUN MH, XIE SX, LU XP, et al., 2013. Effects of NaWO4 treatment on growth and nitrate nitrogen content in Poncirus trifoliate seedlings [J]. Nonwood For Res, 31(4): 115-120. [孙敏红, 谢深喜, 卢晓鹏, 等, 2013. 钨酸钠处理对枳幼苗生长及植株硝态氮含量的影响 [J]. 经济林研究, 31(4): 115-120.]
SUN YX, YAO XH, REN HD, 1999. Growth variance and character analysis of Cinnamomum camphora seedling from different provenances [J]. J Zhejiang For Coll, 16(3): 234. [孙银祥, 姚小华, 任华东, 1999. 樟树种源苗期差异及性状相关 [J]. 浙江林学院学报, 16(3): 234.]
TAN ZQ, CHEN Y, LIU X, et al., 2019. Comparison of growth and photosynthetic traits of Michelia macclurei from six provenances in eastern areas of Guangxi [J]. Guangxi For Sci, 48(2): 147-151. [谭长强, 陈依, 刘秀, 等, 2019. 桂东地区6个种源火力楠生长及光合特征比较 [J]. 广西林业科学, 48(2): 147-151.]
WANG QY, LI FJ, YAN SP, et al. , 2003. The influence of nitrate reductase and soluble protein on the provenance growth of Picea koraiensis [J]. J NE For Univ, 31(5): 59-61. [王秋玉, 李凤娟, 闫绍鹏, 等, 2003. 硝酸还原酶和可溶性蛋白对红皮云杉种源生长的影响 [J]. 东北林业大学学报, 31(5): 59-61.]
WANG TT, WEI XL, ZHANG L, 2019. Comparison and evaluation of growth characteristics in Trachycarpus fortunei seedlings from different provenances in Guizhou [J]. Non-Wood For Res, 37(3): 146-152. [王婷婷, 韦小丽, 张兰, 2019. 贵州不同种源棕榈苗期生长性状比较及評价 [J]. 经济林研究, 37(3): 146-152.]
WANG XD, LIU P, LIU MJ, et al., 2018. Biology and ecology research status of Ormosia species in China [J]. Plant Sci J, 36(3): 440-451. [王小东, 刘鹏, 刘美娟, 等, 2018. 中国红豆属植物生物与生态学特征研究现状 [J]. 植物科学学报, 36(3): 440-451.]
XIA X, PAN WT, LI YQ, et al., 2018. An analysis on the growth characteristics andvariation regulation of different provenances of Liriodendron seedling [J]. Acta Agric Univ Jiangxi, 40(1): 118-126. [夏莘, 潘文婷, 厉月桥, 等, 2018. 鹅掌楸不同种源苗期生长特征及其变异规律分析 [J]. 江西农业大学学报, 40(1): 118-126.]
XU FL, WEI XL, GU DH, et al, 2015. The main pest species, harm and control methods of Ormosia henryi in Guizhou [J]. For Sci Technol, (3): 41-43. [徐芳玲, 韦小丽, 古定豪, 等, 2015. 贵州花榈木主要害虫种类、危害及防控方法 [J]. 林业科技通讯, (3): 41-43.]
XUE ML, FANG YM, DAI CZ, et al., 2019. Differences in leaf characteristics and water use efficiency of Quercus acutissima seedlings from different provenances [J]. North Hortic, (13): 101-109. [薛美玲, 方炎明, 戴长志, 等, 2019. 不同种源麻栎幼苗叶片性状与水分利用效率的差异 [J]. 北方园艺, (13): 101-109.]
YAO JB, CHU XL, ZHOU ZC, et al., 2018. Response of seedlings of three Schima superba provenances to different light environments when mixed planting with Cunninghamia lanceolata [J]. For Res, 31(6): 144-153. [姚甲宝, 楚秀丽, 周志春, 等, 2018. 3个与杉木混植木荷种源幼苗对光照响应的差异 [J]. 林业科学研究, 31(6): 144-153.]
ZHANG LW, ZHONG GC, ZHANG L, et al., 2012. A study on photosynthesis and photo-response characteristics of three Salvia species [J]. Acta Pratac Sin, 21(2): 70-76. [张力文, 钟国成, 张利, 等, 2012. 3种鼠尾草属植物光合作用-光响应特性研究 [J]. 草业学报, 21(2): 70-76.]
ZHANG ZA, ZHANG MS, WEI RH,2004. Experimental guidance of plant physiology [M]. Beijing: China Agricultural Science and Technology Press: 43-45. [張治安, 张美善, 蔚荣海, 2004. 植物生理学实验指导 [M]. 北京: 中国农业科技出版社: 43-45.]
ZHENG WJ, 1985. Tree records of China [M]. Vol. 2. Beijing: China Forestry Publishing House. [郑万钧, 1985. 中国树木志 [M]. 2册. 北京: 中国林业出版社.]
ZHOU XB, ZHOU W, DENG LT, et al., 2016. Comparison of photosynthetic characteristics among 14 Toona sinensis provenances [J]. J S Chin Agric Univ, 37(4): 51-56. [周祥斌, 周玮, 邓丽婷, 等, 2016. 14个香椿种源光合特性的比较研究 [J]. 华南农业大学学报, 37(4): 51-56.]
(责任编辑 何永艳)
猜你喜欢
中国药学药品知识仓库(2022年9期)2022-05-23
江苏农业学报(2019年1期)2019-09-10
湖北农业科学(2018年8期)2018-06-08
Coco薇(2017年5期)2017-06-05
大学教育(2016年11期)2016-11-16
中小企业管理与科技·上旬刊(2016年10期)2016-11-15
考试周刊(2016年84期)2016-11-11
吉林农业(2016年7期)2016-05-14
环球时报(2015-05-13)2015-05-13
江苏农业科学(2014年10期)2014-11-22
