
竹叶兰花器官发育过程及生理特性研究
2021-02-22 07:31吴洁秋朱根发王凤兰杨凤玺
热带作物学报 2021年1期
关键词:抗氧化酶
吴洁秋 朱根发 王凤兰 杨凤玺
摘 要:本研究通过解剖形态学观察,并检测主要代谢物质、抗氧化酶活性以及内源激素含量的变化,阐述竹叶兰花器官发育特征及生理特性,为新花卉作物的开发利用提供理论依据。结果表明:(1)广州地区户外栽培条件下,竹叶兰全年可开花,为总状花序,单朵次第开花,整枝花期188 d;(2)根据花器官发育特征分为5个时期:花芽分化期、萼片伸长期、合蕊柱发育期、花瓣着色期及花朵绽放期,单花发育仅需32 d;(3)进入生殖生长后,可溶性糖、淀粉和可溶性蛋白含量均显著上升,可溶性糖含量最高,其次为可溶性蛋白;超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性呈平稳上升趋势;(4)生长素(IAA)、赤霉素(GA3)含量在营养生长期持续上升,在花芽发育期呈先下降后上升趋势;与之相反,脱落酸(ABA)含量在花芽分化期显著上升,在花芽发育后期下降;推测竹叶兰体内低水平的IAA、GA3和高水平的ABA可能对花芽分化起重要调控作用。
关键词:竹叶兰;花器官发育;营养代谢;抗氧化酶;内源激素
中图分类号:S682.31 文献标识码:A
Abstract: In this study, we observed the anatomical and morphological characteristics of the flower organs and detected the changes of metabolite content, antioxidant enzyme activity, and the content of endogenous hormones (IAA, GA3, and ABA) to explore the physiological and biochemical characteristics of continuous flowering of Arundina graminifolia (D. Don) Hochr., which could provide theoretical basis for the development and utilization of new flower crops. When cultivated under natural conditions in Guangzhou, A. graminifolia could flower throughout the year; with single flower blooms in turn in a long raceme and the flowering period of the whole inflorescence could reach 188 days. According to the characteristics, the floral organ development could be divided into five periods: flower bud differentiation, sepal growth, pillar development, flower dyeing and flower opening. This process took about 32 days. After entering the reproductive growth, the soluble sugar, starch, and soluble protein contents increased significantly, with the highest content of soluble sugar, followed by soluble protein. SOD, POD and CAT activities showed a steady upward trend. The content of IAA and GA3 increased continuously in vegetative growth period, and decreased first and then increased in flower bud development period. In contrast, ABA content increased significantly at the flower bud differentiation stage and decreased at the later stage of flower development. It was speculated that low level of IAA, GA3 and high level of ABA might play an important role in regulating flower bud differentiation.
Keywords: Arundina graminifolia; flower organ development characteristics; nutrient metabolism; antioxidant enzyme; endogenous hormone
DOI: 10.3969/j.issn.1000-2561.2021.01.020
兰科植物(Orchidaceae)又称兰花,是被子植物中的最大科,全世界约有738属28 000种[1-2]。《中國植物志》(1999年版)记载的中国兰花约有171属1247个原生种,分布于热带、亚热带及温带地区,按生境的不同分为地生兰、附生兰和腐生兰,少数为攀援藤本。兰花是被子植物中进化水平最高的类群之一[3],长期以来作为生态进化研究的理想材料,同时也是全球重要的商品花卉。开花特性一直是兰科花卉研究的热点,但不同兰花种类的生态习性和开花特性存在显著差异,大多需要适宜的环境条件才能正常开花,造成商品兰花成本增加且难以实现周年生产和应节、应季开花。如蝴蝶兰属于典型的耐阴植物,强光下停止生长甚至休眠,在适宜光照下,花芽分化的最佳温度为白天25 ℃,夜间20 ℃,超过28 ℃时花芽难以形成[4-5]。传统国兰如墨兰[6]、春兰[7]等在25~28 ℃下花芽分化完成,但需要20 ℃/ 10 ℃低温条件促进花器官发育,否则会造成花芽败育;此外,研究发现16 h长日照对春兰生殖发育起促进作用,但对寒兰花期调控则表明8 h短日照条件下花期提前的幅度更大[8-9];短日照同时也可促进石斛兰[10]、海南钻喙兰[11]花期提前。
竹叶兰[Arundina graminifolia (D. Don) Hochr.]是兰科(Orchidaceae)竹叶兰属(Arundina)地生兰,为多年生常绿草本植物[3],因叶似竹叶,丛生茎直立、分节,故得名“竹叶兰”,别称有长杆兰、草姜、山荸荠、幽涧兰等[12],主产于东南亚地区,在我国浙江、江西、广东、海南、云南等华南、西南地区均有分布[13]。傣族人民称其为“文尚海”[14],是傣族“雅解药”的主要药材成分,具有清热解毒、散瘀止痛、除湿利尿的功效[15]。竹叶兰不仅是珍贵的药用植物,也是具有很高观赏价值和广阔市场前景的兰科花卉。竹叶兰花朵酷似卡特兰,花型大而美丽,花色淡雅,尤其常年可开花,适应性强,在新加坡、夏威夷、马来西亚等地深受欢迎,正被大力发展成为一种新花卉作物,广泛用于园林绿化和盆栽。
然而在我国,竹叶兰被用于传统解毒药,目前尚未引入观赏领域;且随着竹叶兰的大量开采,野生资源日趋减少,被列为国家Ⅱ级保护植物[16]。近年来大多学者对竹叶兰的研究主要集中在化学成分[17-22]、药理活性[23-26]等生物活性物质的鉴定,少数学者阐述了其生长习性和栽培技術[12-13]以及繁殖方法[27],对其开花生理特性和园林应用的研究鲜有报道。了解和掌握竹叶兰生长发育和开花习性,是充分发挥竹叶兰观赏价值和经济效益的关键。
本文通过研究竹叶兰开花进程、营养物质代谢、抗氧化酶活性及内源激素赤霉素(GA3)、生长素(IAA)和脱落酸(ABA)的动态变化规律,初步探究其开花生物学特征,旨在为竹叶兰开花调控、栽培管理以及作为新花卉作物的产业开发奠定基础。
1 材料与方法
1.1 材料
供试验的竹叶兰原生地为广东省中山市,经驯化栽培和人工扩繁后,2017年栽植于广东省农业科学院环境园艺研究所试验田(23?9? N,13?21? 10? E),每丛3~4株,丛距为10 cm×10 cm,实施精细化田间管理。目前,植株生长良好,全年开花。本研究自2018年9月3日起至2019年7月31日,在广东省农业科学院环境园艺研究所试验田和广东省园林花卉种质创新综合利用重点实验室进行。
1.2 方法
1.2.1 形态观察测定 (1)竹叶兰生长指标测定。营养生长期监测移栽后平均株高9.63 cm幼苗(20株)的生长形态指标,从2018年9月至2019年7月开展3次重复试验,用直尺测量株高(茎基部至茎尖生长锥),并计录叶片数,每7 d监测1次,取平均值,监测周期为11周。生殖生长期选取大小长势一致且生长健壮,顶端叶芽刚转化为花芽(用手轻捏顶端,呈微鼓状,剥开叶鞘有萌发的花芽)的成苗20株。用游标卡尺分别测量植株的茎粗(从基部起第3节茎)、花梗直径、花横径(两侧瓣尖端的距离)和纵径(中萼片尖端至唇瓣最外缘的距离,花横、纵径以花序上第3朵花为标准)。用直尺分别测量株高(茎基部至茎尖生长锥/花梗基部)、花梗节间长度。株高、叶片数每7 d测量1次,取平均值;叶片数、花梗节数、花朵数、单花观赏期和单花寿命(以花序上第3朵花为标准)以及花序寿命,每3 d观察1次,取平均值。
(2)花发育形态建成过程的显微观察。从顶端分生组织进入成花转变期开始观察,每次采摘3~5个大小一致的花芽(或花分生组织),在Leica 205FA体视显微镜下解剖观察、拍照并记录。样本较大时,使用单反数码相机(Nikon-D90)拍照记录。每3 d观察1次,3次重复。
1.2.2 生理生化特性 选取生长良好、无病虫害的竹叶兰幼苗(苗龄4周,株高8~10 cm)、快速生长健壮苗(苗龄8周,株高20~22 cm)茎尖第1~2片嫩叶和生殖生长阶段5个不同发育期的花器官,共7个不同发育阶段的植物材料。取样时间为2018年11月22日、12月22日和2019年1月22日,设3次生物学重复实验,取平均值。采用752 N紫外分光光度计测定可溶性蛋白浓度、可溶性糖、淀粉含量和过氧化物酶(POD)活性,采用BIO-RAD IMark酶标仪测定超氧化物歧化酶(SOD)活性,试剂盒购自南京建成生物工程研究所;过氧化氢酶(CAT)活性采用紫外分光光度法测定。内源激素(生长素IAA、脱落酸ABA和赤霉素GA3)含量由上海酶联生物有限公司,采用酶联免疫法检测。
1.3 数据处理
采用Excel 2007软件进行数据统计并制图,采用SPSS 19.0软件进行显著性分析。
2 结果与分析
2.1 竹叶兰营养生长特性
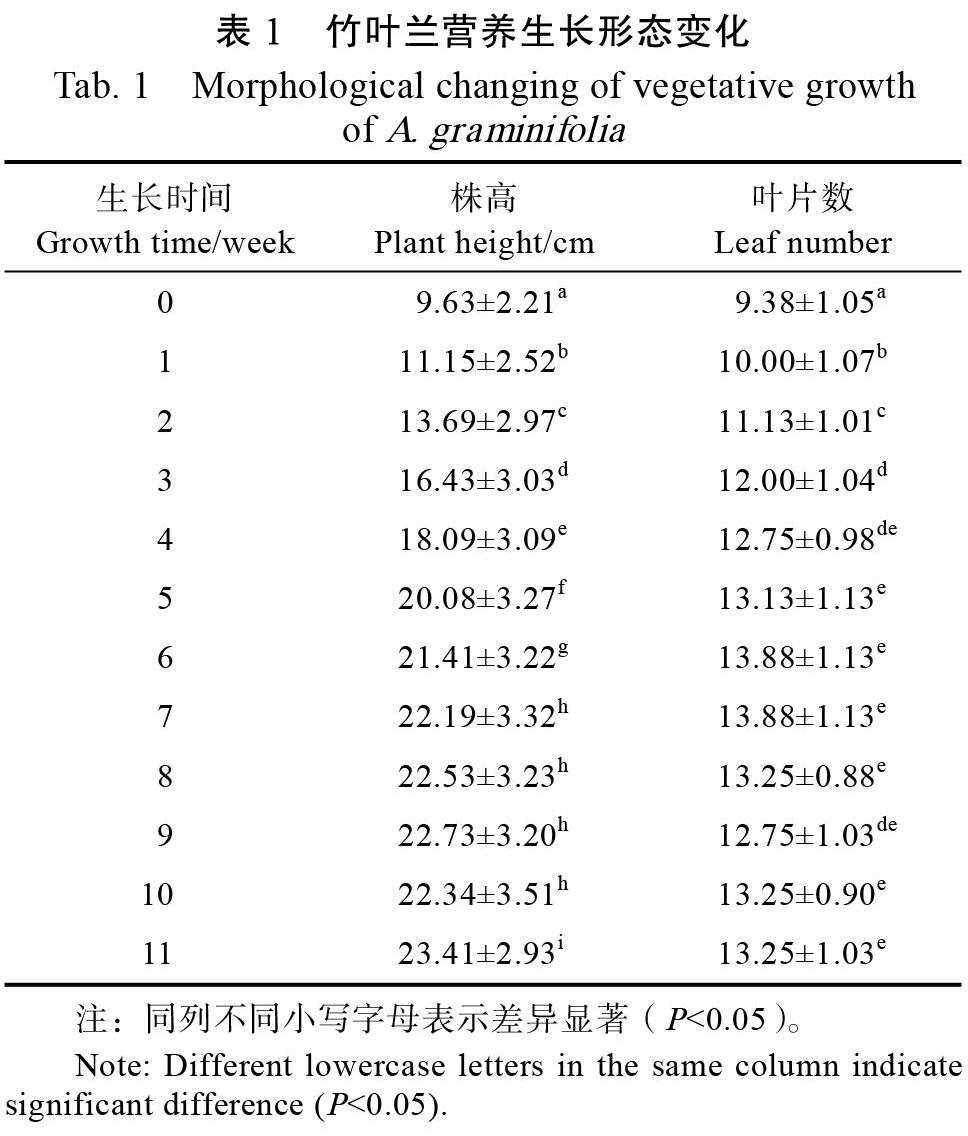
竹叶兰是兰科多年生草本植物,株高可达80 cm[3],叶似禾本科植物的叶子,叶鞘抱茎,茎有节,似竹子,几乎全年开花。通过监测竹叶兰营养期生长指标,由表1可知,竹叶兰平均生长速率达1.25 cm/周。叶片数在前期增长较快,茎尖生长锥几乎每周新生一叶,后期部分植株随着新叶抽出,基部叶片枯萎凋落,因此叶片数变化不大。
结果显示,竹叶兰营养生长阶段总体生长速率较快,前期株高和叶片数表现为快速增长,后期增长较缓慢。基部叶片凋落,可能是随着植株生长,体积增大,株距缩小,使得基部光照不足,而且广州市夏季气候炎热,进而导致干旱等环境胁迫所致。此外,竹叶兰在适宜条件下,株高长至27~34 cm时营养生长缓慢或停止,开始逐渐转变为生殖生长。
2.2 竹叶兰生殖生长特性
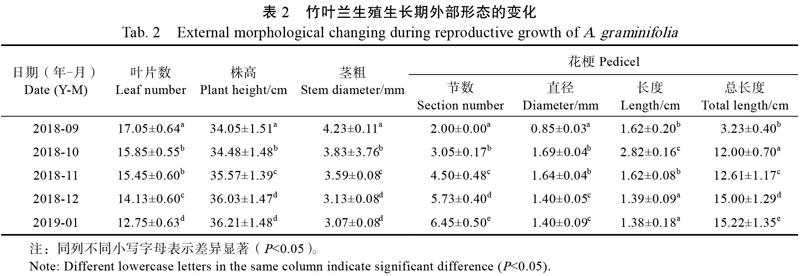
2.2.1 外部形态特征 观察发现,在广州地区自然条件下,竹叶兰常年开花,9月至次年1月开花最旺盛。对该时期形态指标的测量结果见表2,2018年9月植株花芽开始分化,株高为34 cm/株,次年1月整枝花期结束时株高约36.21 cm/株,月均株高增0.54 cm/株。茎基部老叶脱落,叶片数呈下降趋势,9月花芽分化时17.05片/株,到次年1月减至12.75片/株。
对竹叶兰花序整体结构观察结果显示,竹叶兰为总状花序,平均着花6.12朵/株。单花平均寿命为32.29 d,花朵次第开放,整枝花序观赏期可达188.18 d。花朵橫、纵径分别为39.12 mm和34.50 mm左右,单朵盛花期约4.41 d。自花梗基部起第1、2节节间长度大于第3、4节以后的节间长度,第2节最长,约2.82 cm,是最后1节节间长度(1.38 cm)的2倍(表2)。
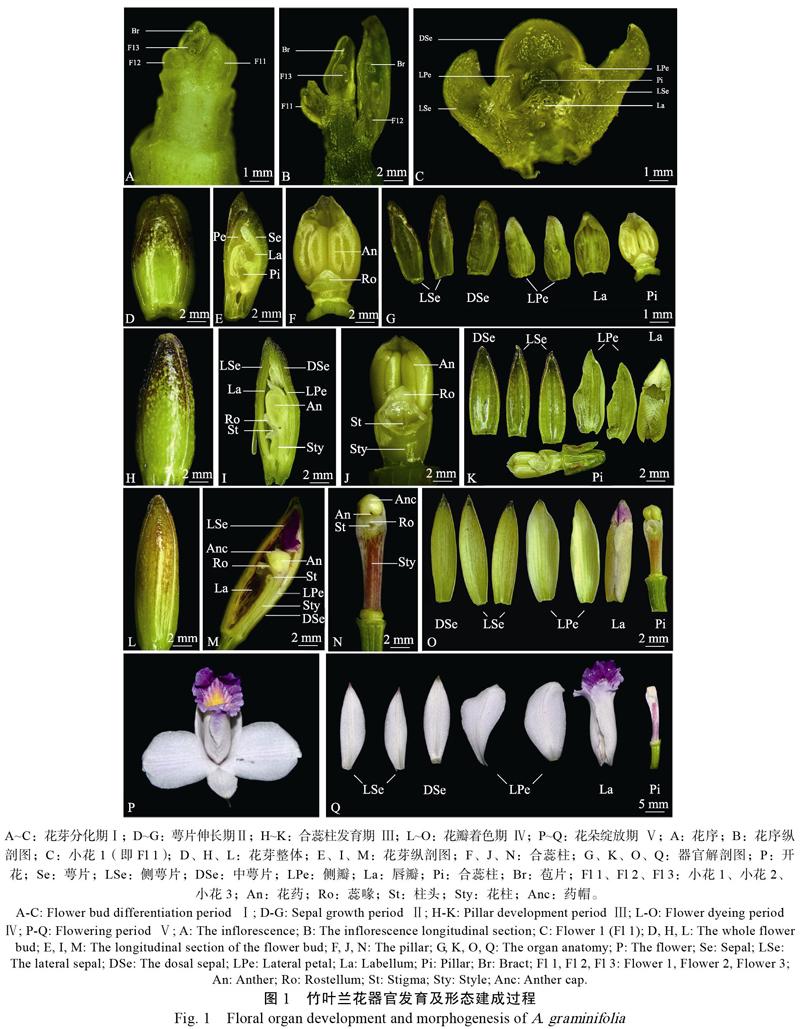
2.2.2 花器官发育 解剖形态学观察发现,竹叶兰小花主要由萼片(中萼片1个和侧萼片2个)、侧花瓣2个、唇瓣和合蕊柱7部分组成。其开花进程根据花器官发育的特征,可分为以下5个阶段:
花芽分化期(Ⅰ):竹叶兰顶端分生组织转变为花序分生组织,生长锥发生一系列变化,形成肉眼可见凸起,即花芽。此时花芽直径约2 mm,剥开苞片可见正在分化的花序(图1A)。从小花形态及着生位置观察结果显示,小花Fl 1发育最早,各器官初步分化完成,7个基本器官清晰可辨(图1C);其次为Fl 2,呈圆球状凸起,未形成主要花器官;Fl 3发育最晚,苞片与花原基相互黏合(图1A,图1B)。
萼片伸长期(Ⅱ):小花Fl 1继续分化历时约7 d,萼片快速伸长,肉质肥厚,形似“勺”状,表皮上端褐色,约占整个花芽的1/2表面积,下端绿色(图1D);中萼片面积最大,左右萼片狭长(图1G)。此时期,花瓣浅绿色,瓣纹清晰;花药膨大、黄绿色;合蕊柱未伸长(图1E,图1F,图1G)。
合蕊柱发育期(Ⅲ):花蕾生长2周后,合蕊柱显著伸长,蕊喙和柱头分化发育明显,呈透明状,花药收缩变小(图1I,图1J)。萼片长而窄,肉质变薄;唇瓣表现特异化,周缘皱褶,仍表现为浅绿色(图1H,图1K)。
花瓣着色期(Ⅳ):合蕊柱发育1周以后,花瓣开始显色。唇瓣褶片由黄绿色变为粉/紫色,小面积呈黄色,以下则为白色,表面带粉/紫色瓣纹,内侧附红褐色条纹;左右侧瓣变白,但瓣纹仍为浅绿色(图1M,图1O)。此时,萼片表皮褐色变浅,扩散至整个花蕾2/3的表面积,内侧由绿变白,开始瓣化(图1L,图1O)。花柱凹面一侧变为暗红色,白色条纹稀疏分布(图1N)。
花朵绽放期(Ⅴ):花蕾经过3~4 d的着色完善后,花朵完全展开(图1P),唇瓣、侧瓣和萼片明显外翻(图1Q)。萼片瓣化,面积小于花瓣,合蕊柱紧靠唇瓣开裂一侧,颜色鲜亮,整体花型较大,美丽高雅。
小花Fl 1花器官发育成熟,开放周期维持4~6 d,随后萎蔫脱落或虫媒授粉结果。小花Fl 2花器官发育稍晚于小花Fl 1,紧随小花Fl 1凋谢后绽放。整枝花序循环往复上述发育过程,直至完全开花结束,花梗枯萎。
2.3 竹叶兰可溶性糖、淀粉及可溶性蛋白含量
竹叶兰幼苗期和快速生长期叶片中可溶性糖、淀粉和可溶性蛋白含量变化不大;转入花芽分化期后迅速上升且呈显著差异(P<0.05),其中,可溶性糖含量比幼苗期和快速生长期分别提高38.26%和36.34%;淀粉含量上升幅度与可溶性糖相似,分别比幼苗期和快速生长期高38.75%和36.62%。可溶性蛋白在花芽分化期的上升幅度最为显著,分别比幼苗期和快速生长期提升55.96%和48.47%。整体看来,竹叶兰主要营养物质含量大小依次为可溶性糖>可溶蛋白>淀粉(图2)。
可见,可溶性糖、淀粉和可溶性蛋白质是竹叶兰生长发育过程中的重要基础物质,营养物质的积累,尤其是充足的可溶性糖和蛋白质,有利于竹叶兰花芽分化。
2.4 竹叶兰SOD、POD及CAT活性
竹叶兰SOD、POD和CAT活性总体均呈平稳上升趋势,相较于幼苗期和快速生长期,花芽分化期抗氧化酶活性上升幅度较明显,其中,SOD活性分别增强了25.69%和15.8%,POD活性分别提高31.48%和13.12%。CAT活性在快速生长期稍有下降,转入花芽分化期后快速上升,较快速生长期提升了31.48%。3个时期抗氧化酶活性从大到小排列依次为SOD>POD>CAT(图3)。
由此推测,自然环境条件下,竹叶兰不同生长期体内保持一定水平的SOD、POD和CAT活性,以适应复杂的外界环境胁迫,保证植株生长发育的正常进行;同时,通过调节酶活性,影响竹叶兰从营养生长到生殖生长的转变。
2.5 竹叶兰IAA、GA3和ABA含量及相互间的平衡
2.5.1 竹叶兰IAA、GA3和ABA含量 通过测定竹叶兰不同生长发育时期生长素(IAA)、赤霉素(GA3)和脱落酸(ABA)含量变化表明,IAA和GA3含量变化趋势基本一致,营养生长期持续上升,花芽发育期先降后升。其中,快速生长期IAA和GA3含量上升达到峰值,分别为16.13 μmol/L和91.59 pmol/L,在花芽分化期快速下降,分别降至8.47 μmol/L和40.13 pmol/L,低于快速生长期47.49%和56.22%(图4A,图4B)。
与之相反,ABA含量在植株快速生长期达最低值约为0.1 μmol/L,是幼苗期的23.08%,并在进入花芽分化期后迅速上升到0.2 μmol/L,相比快速生长期提高了1倍,在花发育第2阶段(萼片伸长期)升至最大值,约0.26 μmol/L,随后逐渐降低,开花期间维持在大约0.17 μmol/L水平(图4C)。
可见,低水平IAA、GA3有利于竹叶兰生殖生长,高水平IAA、GA3促进植株营养生长。与之相反,高水平ABA对生殖发育产生一定积极影响。
2.5.2 ABA/GA3、ABA/IAA及IAA/GA3的比值 植物激素间的协调作用对植物生长发育产生促进或抑制效应。对竹叶兰生长发育不同时期IAA、GA3和ABA的比例关系分析显示(图5),ABA/GA3和ABA/IAA比值变化波动一致,呈先降后升再降趋势。幼苗期至快速生长期ABA/GA3和ABA/IAA比值逐渐降低,花芽分化期迅速上升,萼片伸展出现峰值,随后又下降。IAA/GA3比值变化波动不大,营养生长阶段略低于生殖生长阶段,花发育前期比值较高,后期有所下降。
综上所述,竹叶兰全年开花,适应性强,花大而美丽,是良好的园林观赏兰科花卉;其开花生理生化特性不是单一机制控制的,而是在适宜环境条件下,由植株体内营养物质、抗氧化酶和内源激素等基础物质相互协作、共同调控。有关竹叶兰开花机制有待后续进一步深入研究。
参考文献
Givnish T J, Spalink D, Ames M, et al. Orchid phylogenomics and multiple drivers of their extraordinary diversification[J]. Proceedings of the Royal Society B: Biological Sciences, 2015, 282(1814): 1-10.
Chase M W, Cameron K M, Freudenstein J V, et al. An updated classification of Orchidaceae[J]. Botanical Journal of the Linnean Society, 2015, 177(2): 151-174.
中国科学院中国植物志编辑委员会. 中国植物志[M]. 北京: 科学出版社, 1999:334.
Sakanishi Y, Imanishi H, Ishida G. Effect of temperature on growth and flowering of Phalaenopsis amabilis[J]. Bulletin of the University of Osaka Prefecture B, 1980, 32: 1-9.
刘晓荣. 蝴蝶兰花芽分化及花期调控研究[D]. 杨凌: 西北农林科技大学, 2006.
李淑娴. 墨兰成花机理及花期调控技术研究[D]. 福州: 福建农林大学, 2016.
黄雪梅. 春兰花期调控技术及其生理特性研究[D]. 桂林: 广西师范大学, 2013.
龚 湉. 寒兰成花机理及花期调控研究[D]. 福州: 福建农林大学, 2015.
翁青史. 寒蘭(Gymbidium kanran)花期调控技术及花期生理响应研究[D]. 福州: 福建农林大学, 2019.
李 程, 裴忠孝, 甘林叶, 等. 光周期对春石斛开花及多胺含量的影响[J]. 植物生理学报, 2014, 50(8): 1167-1170.
Phengphachanh B, Naphrom D, Bundithya W, et al. Effects of day-length and gibberellic acid (GA3) on flowering and endogenous hormone levels in Rhynchostylis gigantea (Lindl.) Ridl[J]. Journal of Agricultural Science, 2012, 4(4): 217.
唐德英, 王云娇, 李荣英, 等. 野生竹叶兰引种栽培初报[J]. 中药材, 2005(4): 263-264.
陈之林, 曾宋君, 温铁龙, 等. 竹叶兰的无菌播种和试管成苗[J]. 植物生理学通讯, 2006(1): 66.
王 健. 竹叶兰——珍贵的药用花卉植物[J]. 中国野生植物, 1988(3): 33-34.
李文军, 朱成兰, 唐自明. 傣药竹叶兰的生药学研究[J]. 云南中医中药杂志, 2000(6): 32-33.
冯欣欣, 余金昌, 袁志永, 等. 国家Ⅱ级保护植物竹叶兰的迁地保护与野外回归研究[J]. 中国园艺文摘, 2017, 33(3): 12-14.
李玉鹏, 瓮瑞旋, 陶永生, 等. 云南傣药竹叶兰化学成分的研究[J]. 昆明医科大学学报, 2016, 37(11): 5-7.
刘美凤, 吕浩然, 丁 怡. 竹叶兰中联苄类化学成分和抗肿瘤活性研究[J]. 中国中药杂志, 2012, 37(1): 66-70.
彭 霞, 何红平, 卯明霞, 等. 竹叶兰的化学成分研究[J]. 云南中医学院学报, 2008, 31(3): 32-33.
刘美凤. 傣药竹叶兰的化学成分研究Ⅰ[C/OL]2006第六届药学会学术年会会议论文集, 2006: 5119-5122 [2019-12-15].https://kns.cnki.net/kns/ViewPage/viewsave.aspx?t=1609809362750.
刘美凤. 傣药竹叶兰化学成分研究与抗抑郁新药YL102的药学研究[D]. 北京: 清华大学, 2004.
Auberon F, Olatunji O J, Waffo-Teguo P, et al. New glucosyloxybenzyl 2R-benzylmalate derivatives from the undergrounds parts of Arundina graminifolia (Orchidaceae)[J]. Fitoterapia, 2019, 135: 33-43.
闫雪孟, 汤冰雪, 刘美凤. 竹叶兰提取物抗氧化与抗菌活性研究[J]. 时珍国医国药, 2017, 28(12): 2862-2864.
宋新月, 汤冰雪, 邱君志, 等. 竹叶兰内生真菌的分离鉴定及其抗氧化活性研究[J]. 现代食品科技, 2018, 34(2): 82-88.
陈毅坚, 石 雪, 屈 睿, 等. 竹叶兰不同部位抗氧化活性比较研究[J]. 中药材, 2013, 36(11): 1845-1849.
Liu Q Q, Sun F Y, Deng Y L, et al. HPLC-ESI-MSn identification and NMR characterization of glucosyloxybenzyl 2R-benzylmalate deriva-tives from Arundina graminifoliaand their anti-liver fibrotic effects in vitro[J]. Molecules, 2019, 24(3): 525.
金建鹏, 朱根发, 杨凤玺. 竹叶兰原球茎的快速诱导与增殖[C/OL].中国观赏园艺研究进展2018, 2018: 475-479 [2020-02-20].https://kns.cnki.net/kns/ViewPage/viewsave.aspx?t=1608539903419.
韦 莉. 蝴蝶兰花芽分化的解剖学观察及成花机理研究[D]. 南京: 南京林业大学, 2010.
郑宝强, 邓茜玫, 李 奎, 等. 不同温度处理对石斛兰花芽分化和发育的影响[J]. 林业科学研究, 2017, 30(3): 460-464.
彭 芳, 田 敏, 王彩霞, 等. 文心兰花芽分化的形态解剖特征及营养物质的动态变化[J]. 浙江农林大学学报, 2012, 29(01): 7-11.
何碧珠, 朱 萍, 王星剑. 不同生长调节剂处理对墨兰花期调控的影响[J]. 北方园艺, 2016(21): 80-84.
蔡中芳. 四季花龙眼花芽分化期营养动态变化的研究[D]. 兰州: 甘肃农业大学, 2010.
林桂玉, 黄在范, 张翠华, 等. 菊花花芽分化期超微弱发光及生理代谢的变化[J]. 园艺学报, 2008, 35(12): 1819-1824.
周厚高, 宁云芬, 张施君, 等. 新铁炮百合生长发育过程的一些生理生化变化[J]. 广西植物, 2003(4): 357-361.
樊卫国, 刘国琴, 安华明, 等. 刺梨花芽分化期芽中内源激素和碳、氮营养的含量动态[J]. 果树学报, 2003(1): 40-43.
梁 芳, 郑成淑, 张翠华, 等. 菊花花芽分化过程中芽和叶片碳水化合物含量的变化[J]. 山东农业科学, 2008(1): 40-42.
赵秋芳, 陈娅萍, 顾文亮, 等. 香草兰花芽分化期蛋白质及碳水化合物变化研究[J]. 热带作物学报, 2015, 36(6): 1053-1058.
庞夫花, 赵密珍, 王 钰, 等. ‘宁玉草莓花芽分化及其生化物质的变化[J]. 果树学报, 2014, 31(6): 1117-1122.
王桐霖, 吕梦雯, 徐金光, 等. 芍药鳞芽年发育进程及生理机制的研究[J]. 植物生理学报, 2019, 55(8): 1178-1190.
张志良, 瞿伟菁, 李小芳. 植物生理学实验指导[M]. 4版, 北京: 高等教育出版社, 2009: 218.
姚 凯, 牛晓娟, 徐 僡, 等. 锶对菠菜幼苗生长、光合和抗氧化酶活性的影响[J]. 核农学报, 2019, 33(6): 1225-1231.
张莹婷, 杨秀莲, 何 岭, 等. 2种石蒜花芽分化与碳水化合物、抗氧化物酶及内源激素变化的关系[J]. 安徽农业大学学报, 2019, 46(2): 342-349.
艾星梅, 何睿宇, 徐永艳, 等. 马铃薯开花与未开花品种的生理生化差异性研究[J]. 江西农业大学学报. 2017, 39(2): 230-236.
李秉真, 李 雄, 孫庆林, 等. 苹果梨花芽分化期几种酶活性的变化[J]. 园艺学报, 2001(2): 159-160.
林榕燕, 陈艺荃, 林 兵, 等. 杂交兰‘黄金小神童花芽分化过程形态与生理变化[J]. 福建农业学报, 2019, 34(2): 170-175.
曹尚银, 张俊昌, 魏立华. 苹果花芽孕育过程中内源激素的变化[J]. 果树科学, 2000(4): 244-248.
Costantini E, Landi L, Silvestroni O, et al. Auxin synthesis-encoding transgene enhances grape fecundity[J]. Plant Physiology, 2007, 143(4): 1689-1694.
王海波, 赵君全, 王孝娣, 等. 新梢内源激素变化对设施葡萄花芽孕育的影响[J]. 中国农业科学, 2014, 47(23): 4695-4705.
Lee T T. Role of phenolic inhibitors in peroxidase-mediated degradation[J]. Plant Physiology, 1977, 59(3): 372-375.
秦建彬, 魏翠华, 余祖云, 等. 大花蕙兰花芽分化与激素关系的研究[J]. 中国农学通报, 2011, 27(31): 109-112.
张 宁, 黄曜曜, 敖 妍, 等. 文冠果花芽分化过程及内源激素动态变化[J]. 南京林业大学学报(自然科学版), 2019, 43(4): 33-42.
庄辉发, 顾文亮, 王 辉, 等. 香草兰花芽分化期叶片内源激素含量的变化[J]. 热带农业科学, 2018, 38(3): 15-19.
冯 枫, 杨际双. 切花秋菊‘神马花芽分化与内源激素的关系[J]. 中国农业科学, 2011, 44(3): 552-561.
牛辉陵, 张洪武, 边 媛, 等. 枣花分化发育过程及其内源激素动态研究[J]. 园艺学报, 2015, 42(4): 655-664.
曾 骧. 果树生理学[M]. 北京: 北京农业大学出版社, 1992: 250-251.
周云龙. 植物生物学[M]. 3版, 北京: 高等教育出版社, 2011:260-261.
曹尚银, 张俊昌, 魏立华. 苹果花芽孕育过程中内源激素的变化关[J]. 果树科学, 2000, 17(4): 244-248.
责任编辑:谢龙莲
猜你喜欢
中国医药导报(2016年33期)2017-03-06
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
安徽农学通报(2016年24期)2017-01-12
江苏农业科学(2015年11期)2016-01-27
江苏农业科学(2015年11期)2016-01-27
河北渔业(2015年10期)2015-10-15
江苏农业科学(2015年8期)2015-09-10
湖南师范大学学报·自然科学版(2015年3期)2015-07-17
