
贵州地方中国樱桃资源的ISSR分析
2021-02-26 08:16刘厚宇陈松梅吴亚维
种子 2021年1期
宋 莎,刘厚宇,陈松梅,赵 凯,吴亚维
(1.贵州省农业科学院果树科学研究所,贵阳 550006;2.贵州大学农业生物工程研究院,贵阳 550025;3.贵州大学林学院,贵阳 550025;4.贵州省清镇市农业农村局,贵阳 550004
贵州地方樱桃资源十分丰富,但对资源保护、研究利用方面的报道相对较少。李金强等[1]对贵州樱桃资源生态分布、主要特征和利用现状进行初步调查。宋常美等[2]利用ISSR技术对贵州樱桃资源进行遗传多样性分析,发现贵州地方樱桃资源的多样性丰富,其中毕节、遵义及黔南等地资源具有丰富的多样性,尤其毕节地区拥有大量品质优良的资源,可作为育种的亲本。欧茂华[3]对贵州樱桃属植物进行调查,发现共有16个种,其中典型樱桃亚属12个种和矮生樱桃亚属4个种。然而近年来,少数人采取砍枝、砍树的掠夺式采收方式,一些优良株系逐渐被毁掉,导致本地资源破坏严重[1]。为更好地利用贵州地方樱桃资源,贵州省果树科学研究所自2015年起收集省内主要地区的中国樱桃种质资源,并建立了贵州樱桃资源圃进行资源保存和评价[4]。本研究利用ISSR (Inter-simple Sequence Repeats Polymorphism)分子标记技术[5-6]对一些野生资源和该樱桃资源圃中近40种樱桃进行遗传多样性检测,为这些资源的保存及有利基因挖掘和新品种选育提供基础资料和理论参考。
1 材料与方法
1.1 材 料
供试材料为中国樱桃资源的幼叶,共计40份(表1),其中5份资源为野生樱桃(包括32号和33号,35号云南樱桃,38号微毛樱桃和40号采自雷公山的野生樱桃),其余材料采自贵州省农科院果树科学研究所落叶果树种质资源圃。3月中旬,取幼叶于自封袋,冰盒保存迅速带回实验室,清水洗净后硅胶吸干水分,液氮处理置于-80 ℃℃冰箱保存。
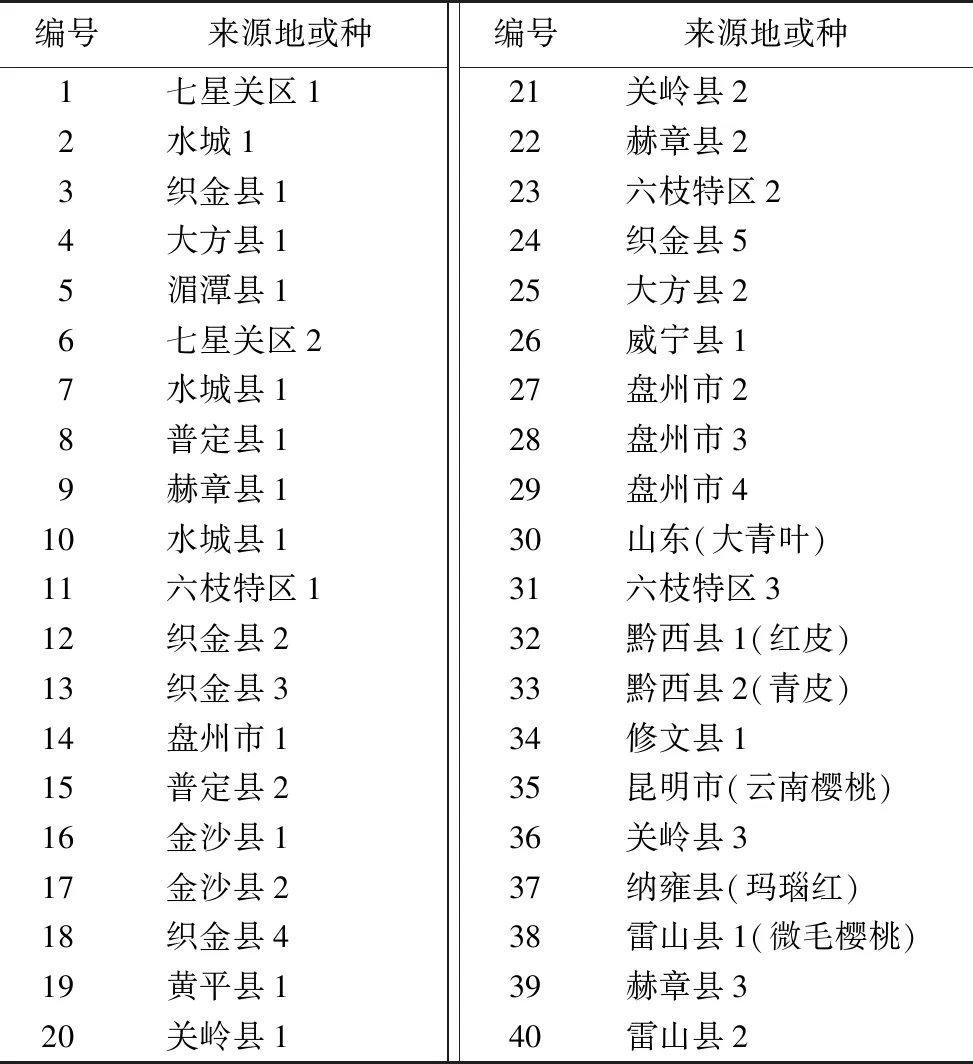
表1 供试樱桃材料Table 1 Test materials of P.pseudocerasus
1.2 ISSR引物
本试验所用ISSR引物(见表2),参照宋常美等[2]、田田等[7]的方法。
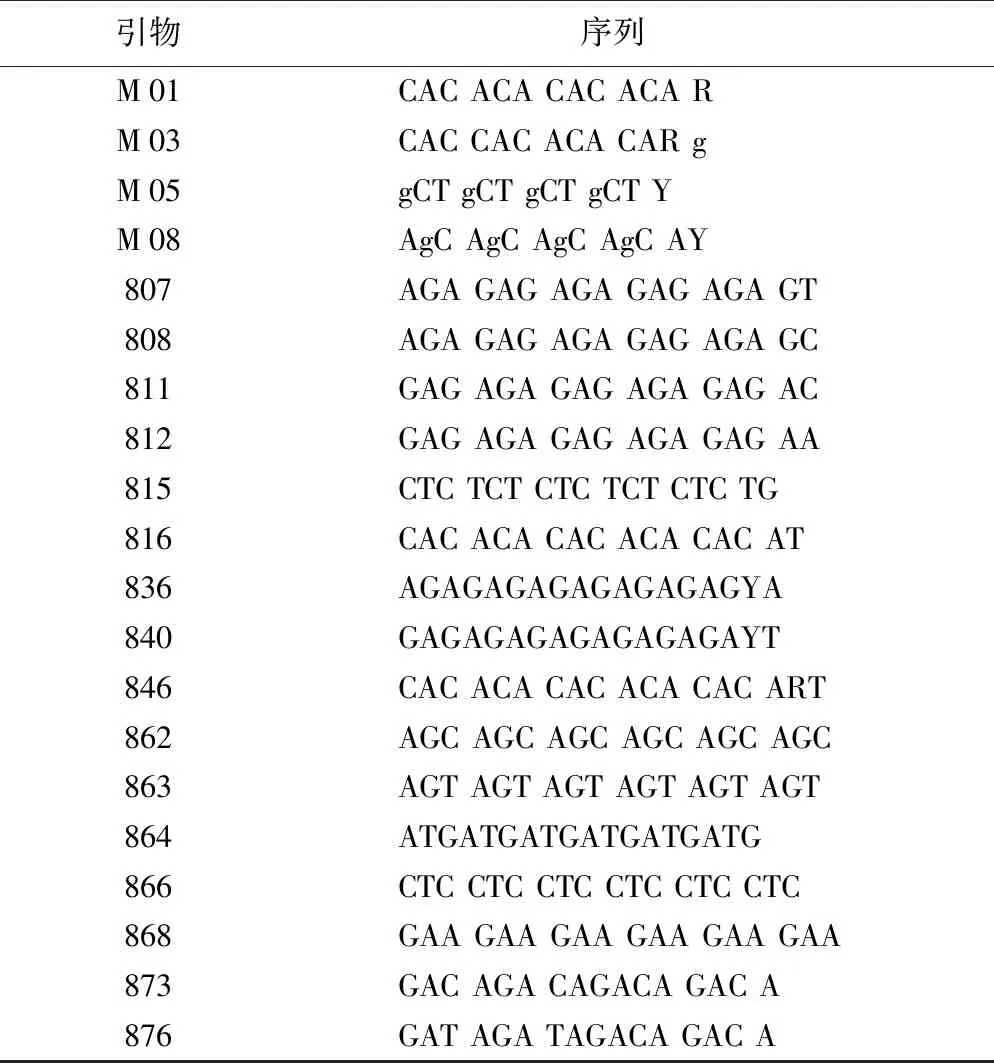
表2 用于ISSR扩增的随机引物及其序列Table 2 Random primers and sequences for ISSR amplification
1.3 DNA提取、PCR扩增及检测
采用北京天根生物科技公司的植物DNA提取试剂盒(DP 302)。使用核酸微量测定仪及琼脂糖凝胶电泳检测基因组DNA浓度及质量。PCR扩增程序为 94 ℃预变性3 min,94 ℃变性1 min,Tm(视引物而定)1 min,72 ℃延伸1 min,38个循环,72 ℃延伸7 min,4 ℃终止反应。利用8%非变性聚丙烯酰胺凝胶检测PCR产物,电泳为150 V,电泳时间根据不同浓度设定,范围在1.2~2.5 h之间。凝胶取出后放入含有5%冰醋酸和10%乙醇的固定液容器中,置摇床上 100 r·min-1固定 15 min。0.1%的硝酸银置摇床上 100 r·min-1银染 15 min,迅速用dd H2O快速清洗2次。加入显色液(-20 ℃预冷10 min)(包含1.5% NaOH+1.1%甲醛)加入轻摇至条带清晰,用dd H2O冲洗胶表面的显色液,拍照分析。
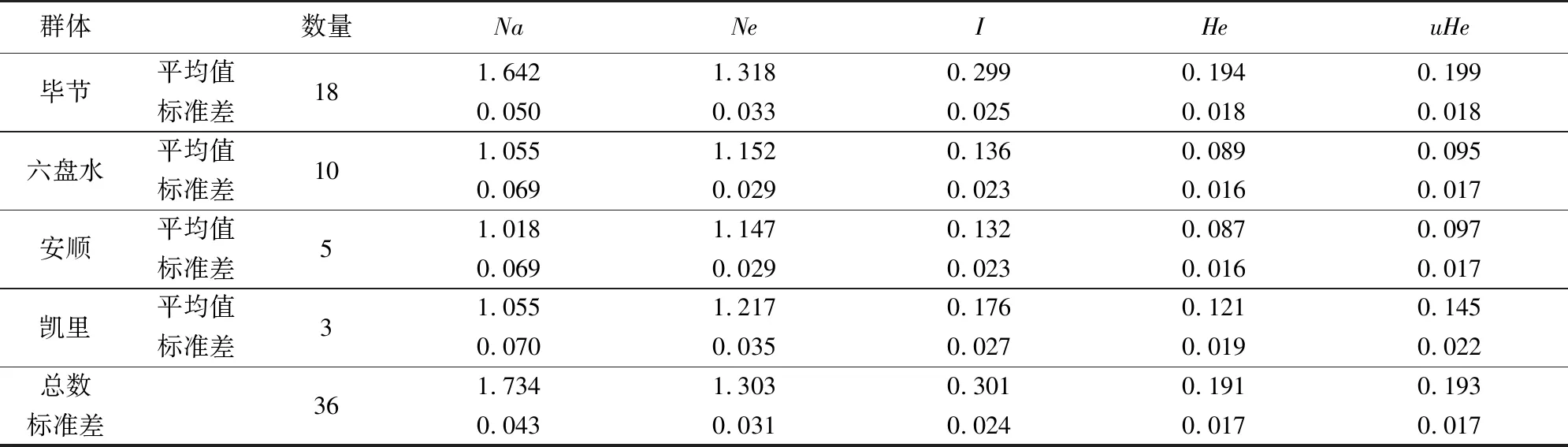
表4 樱桃源产地遗传多样性Table 4 Genetic diversity of origin of P.pseudocerasus
1.4 数据统计与分析
电泳后,统计每个ISSR位点上重复性好且清晰的条带进行记录,以1和0分别记录等位基因的有无。采用POPGENE 1.32软件,服从Hardy-Weinberger平衡,分析计算平均观察等位基因数(Na)、有效等位基因数(Ne)、Nei’s遗传多样性指数(H)、Shannon’s信息指数(I)。利用NTSYS.pc统计分析软件计算遗传相似系数[8]。根据基础值或者遗传距离GD(1-GS)按非加权数据分析法(Unweighted pair-group using arithmetic average,UPGMA)进行遗传相似性聚家族。
2 结果与分析
2.1 ISSR引物的遗传多样性研究
筛选出比较好的12条ISSR引物进行遗传多样性分析,结果(表3)显示,引物位点数在5(引物815)~14(引物807)之间,总位点数是120,平均位点数是10.00。引物多态性位点数在3(引物871)~11(引物807)之间,总数是85,多态性比率在50%~90%之间,平均多态性比率是69.01%。观测等位基因数在1.50~2.00之间,平均1.75。有效等位基因数1.11~1.69之间,平均值为1.30。Nei’s遗传多样性在0.08~0.27之间,平均为0.19。香浓指数在0.14~0.52之间,平均值0.30。表明ISSR位点遗传多样性较为丰富,但不同ISSR引物在揭示遗传多样性方面存在差异,其中,引物866具有最高的多态比率、观测等位基因数、有效等位基因数、Nei’s遗传多样性、香浓指数,是最有效的引物。
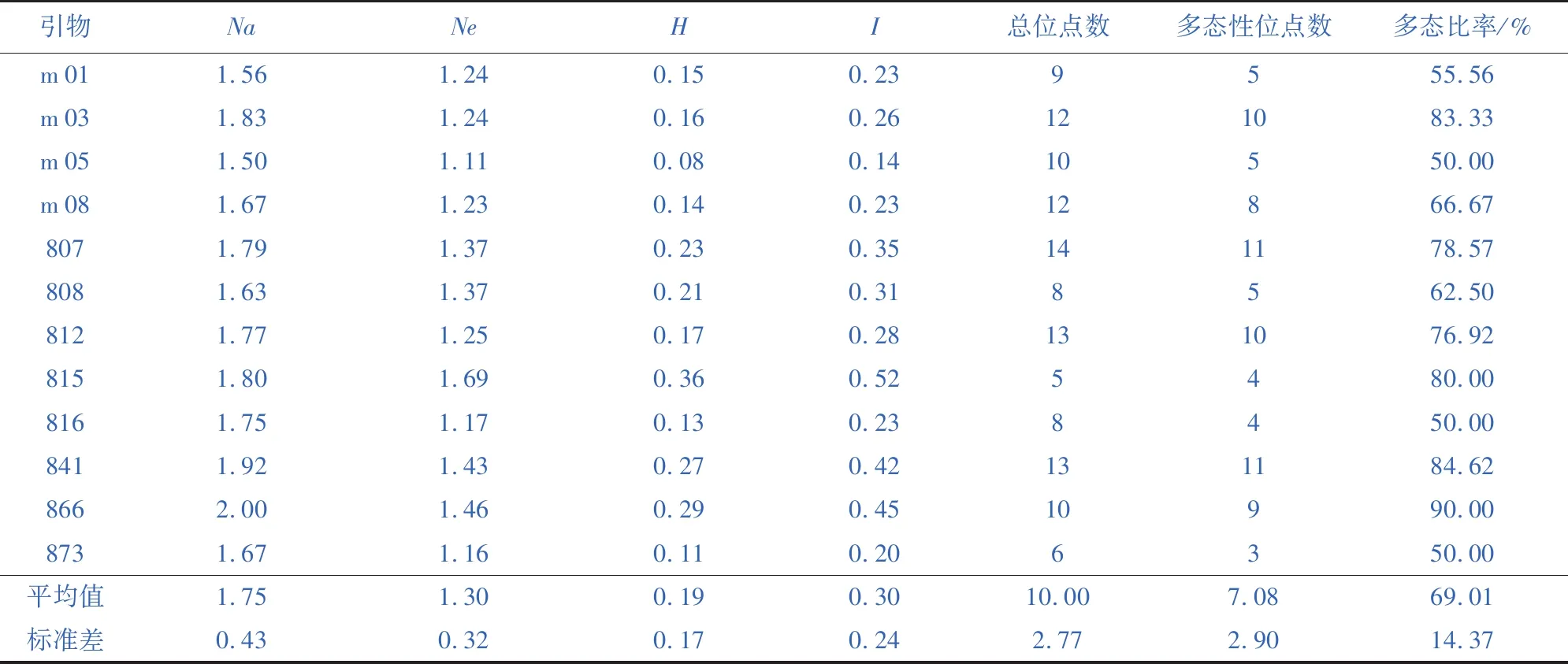
表3 ISSR引物的遗传多样性Table 3 Genetic diversity of ISSR primers
2.2 ISSR检测樱桃遗传多样性分析
以不同采集地区来分析,毕节市群体遗传多样性He最高,达0.194,其次是凯里市(0.121),安顺市和六盘水市分别为0.87和0.89。香浓指数毕节市最高(0.299),其次是凯里市(0.176),安顺市和六盘水市较低。这些数据表明,毕节市有最高的遗传多样性,这与宋常美等[2]的研究结果一致。
2.3 ISSR对樱桃主坐标分析
对野生资源与栽培资源做主坐标分析,结果如图1所示,野生资源和栽培资源遗传距离比较远,尤其是来自黔西县的“红皮”与“青皮”(表1编号32、33;图1中编号38、39)樱桃与其他樱桃遗传距离非常远,而另外3种野生樱桃也能与其余的中国樱桃栽培种区别开。
2.4 群体间的遗传变异
地区群体间的遗传变异系数为8%,群体内为92%,而野生群体与栽培群体间遗传变异系数为35%,群体内遗传变异系数为65%。野生栽培群体Gst值0.350(p>0.001),基因流为0.840。地区种群间Gst值为0.080(p>0.012)。野生与栽培群体间发生的遗传变异大于地区间。Gst值较大,说明杂合子匮乏,纯合子较多。
2.5 樱桃ISSR聚类分析
ISSR分析结果显示(图3),供试材料遗传相似性在0.64~0.98之间,以相似性系数0.81为阈值,可以把野生种和栽培种区分开,野生樱桃可以聚为3群组,黔西“红皮”与“青皮”依然聚在一家族,而且遗传相似性高达0.98,与其他所有樱桃亲缘关系都很远。云南樱桃单独聚为一家族,而雷公山的两种樱桃与玛瑙红樱桃聚为一家族。相比起来,栽培种内的遗传相似性高于野生种内。在以遗传相似性系数0.87为阈值时,栽培种可以聚为3个亚组,第一群组中,包含七星关区1、水城县2、金沙县2、六枝特区2、威宁县1、修文县1、关岭县3、普定县1、水城县3、盘州市2、盘州市1、盘州市4、关岭县1、六枝特区3、普定县2、六枝特区1、织金县3、织金县2、织金县4、织金县 5、盘州市3、赫章县3、大方县2、大青叶(山东)等24份材料。第二亚组有来自织金县1、赫章县2、金沙县1、七星关区2、赫章县1、黄平县1以及湄潭县1等7种樱桃。第三亚组水城1、关岭县2、大方县1等3种樱桃。野生樱桃群组中,比较意外的是,玛瑙红樱桃聚在与之相近的是雷公山的两种樱桃聚在一支(亚组4),而云南樱桃与之遗传距离较远(亚组5),黔西县的两种野生樱桃(亚组6)与所有樱桃遗传距离最远,每一条引物都能将其与其他樱桃区别开来。
一些地理距离近的樱桃遗传距离也近,如盘县发耳镇的两份材料(编号14、29)聚在一起,织金县小纳雍镇2份材料与织金县八步镇的材料(编号12、18、24)聚在一起,雷公山2份材料(编号37、38)聚在一起,黔西县2份材料(编号32、33)聚在一起。同样来自关岭县关索镇的两份材料,其中一份(编号20)与盘县发耳镇两份材料(编号14、29)遗传距离最近,说明可能存在人工引种,或者自然杂交。
3 讨 论
植物遗传多样性反映了植物遗传背景复杂程度及物种存在的历史远近[9]。以栽培资源和野生资源而论,尽管栽培种数量远远高于野生资源,但是野生资源还是显示出高度的遗传多样性和变异度,这与Chen Tao等[11]、Zhang Jing等[12]以及王浩等[13]的研究结果一致,说明野生资源更为古老。与野生资源比,栽培种植资源较低的遗传多一些,可能与人们对良种推广的繁殖的偏向性遭受人工选择压力以及多采用无性繁殖有关[14]。中国樱桃栽培种植资源在长期的驯化过程中经历了严重的奠基者效应以及在近期有效动态变化中产生较强的瓶颈效应,使群体中低频率等位基因严重丢失,而高频率等位基因在群体中有效固定,从而导致总群体遗传多样性丢失[13]。
前期工作中,对部分野生资源应用有所研究,其中来自黔西县的32号、33号樱桃,因春梢嫩枝颜色为红色和绿色,俗称为红皮樱桃与青皮樱桃,聚类和主坐标分析均表明,与其他栽培种和野生种樱桃遗传距离最远。注意红皮樱桃与青皮樱桃的一些特殊性状,如两种樱桃种子可以不经过数月的低温沙藏打破休眠而萌发,也可长时间置于常温干燥条件下而不丧失种子活性,这在其他樱桃中未见报道。这种特殊的萌发性状可能印证了和其他樱桃亲缘关系远,确切的起源还需要进一步从植物形态分类,以及更丰富的樱桃资源进行分子标记来探索。在民间栽培实践中,发现以其作为砧木,能促进大樱桃开花结果。2种樱桃形态相近,仅靠一年生嫩枝颜色差异鉴别,分子标记技术也证明红皮与青皮之间相似性系数为0.98,推测红皮樱桃与青皮樱桃是芽变关系。本课题组正在开展对红皮樱桃与欧洲甜樱桃砧穗互作分子机制的研究。另外,其种子萌发特性对于研究樱桃种子休眠机理提供思路。
玛瑙红樱桃是优质的樱桃品种,关于其起源问题,陈祖瑶等[15]根据记载认为是酸樱桃变异,宋常美等[16]利用分子标记技术推测酸樱桃的实生变异,与酸樱桃相似系数为0.57,其中一个亲本可能是欧洲甜樱桃。本研究发现,玛瑙红樱桃(编号37)与其他中国樱桃栽培种遗传距离较远,聚到野生樱桃这一类中,特别是与来自雷公山的两种樱桃遗传距离最近,推测玛瑙红樱桃可能有野生种的血统,酸樱桃可能属于赫章的樱桃野生种。
根据宋莎等[4]后期对资源圃樱桃性状观察对照本研究结果,同一地理来源的樱桃种质,尽管有部分叶片性状、叶柄颜色等有一定趋同,但是并未严格聚在一类,这种原因产生的结果可能是:
1) 同一地区的采集样本可能会因所处环境因子的差异导致遗传基因的变异;
2) 引物标记位点较少,使得决定性状的关键基因连锁程度较低。本研究结果可以结合这些资源的性状特征进行杂交育种,为进一步利用这些资源提供参考。
野生资源具有较高的遗传多样性,是丰富的基因库,课题组曾发现微毛樱桃具有较高的根癌病抗性,而红皮樱桃作为砧木具有促进甜樱桃成花的作用,有必要进一步发掘野生资源的价值,本研究也为此提供一些理论基础。
猜你喜欢
智慧健康(2021年17期)2021-07-30
晚晴(2021年2期)2021-05-18
晚晴(2019年5期)2019-07-08
新课程·下旬(2018年9期)2018-11-14
红蜻蜓(2017年6期)2017-10-30
湖北畜牧兽医(2015年8期)2015-10-13
青少年科技博览(中学版)(2015年10期)2015-01-11
中国民族民间医药·下半月(2014年2期)2014-09-26
中学生物学(2008年6期)2008-08-29
