
谷关假单胞菌脂肪酶基因PgLip1的克隆和表达
2021-03-12 05:18张冰玉苏小运
中国农业科技导报 2021年3期
张冰玉, 苏小运
(中国农业科学院饲料研究所,农业农村部饲料生物技术重点开放实验室,北京 100081)
脂肪酶(lipase, EC3.1.1.3)是指催化三酯酰甘油水解的酶的总称,是一类特殊的酯键水解酶[1]。其天然底物是长链脂肪酸酯,既可以在两相系统(油、水界面)中起作用,也可以在水相中起作用。脂肪酶作为生物催化剂可催化由不同底物出发的水解和合成反应,且反应条件温和、副产物少。因此,脂肪酶的应用十分广泛,如生物柴油[2-3]、食品行业[4-5]、医药卫生[6-7]、饲料[8]、化学化工[9-11]、环境保护[12]等领域,已成为在生物技术和有机合成方面应用最广泛的一类酶。
脂肪酶来源十分广泛,其中动物、植物、微生物都能生产脂肪酶。微生物来源的脂肪酶种类多,与动植物来源脂肪酶相比,有的微生物脂肪酶作用温度范围更广、稳定性更好、活性更高,适合大规模工业化生产,因此在工业领域更具有应用价值。根据作用温度,脂肪酶可分为高温、中温和低温脂肪酶[13]。与中高温脂肪酶相比,低温脂肪酶的作用温度相对较低,具有低温高催化活性和对热敏感等特性,在食品、水产饲料、洗涤、脂类加工、有机合成以及低温环境修复等方面有着广泛的应用前景[14]。
虽然已有一些低温脂肪酶被发现、克隆和表达,但其数目仍然较少,且性质较为单一。目前,假单胞菌是低温脂肪酶最主要的来源[13],假单胞菌来源的脂肪酶具有反应活性高、稳定性好、反应类型多、有机溶剂耐受性好、应用广泛等特点,特别是在光学活性化合物的拆分与合成时具有较高的立体选择性,因此具有巨大的挖掘潜力[15-17]。谷关假单胞菌(Pseudomonasguguanensis)是2013年新发现的一种假单胞菌。据报道,它可利用聚氧乙烯(20)山梨醇酐单月桂酸酯(吐温-20)和聚氧乙烯脱水山梨醇单油酸酯(吐温-80)作为单一的碳源生长,因此其基因组可编码某种酯酶;该菌还能够耐受高盐离子浓度(7%)生长且具有丰富的酶系[18],因此它所编码的酶可能有一定的抗逆性。此外,Devi等[19]从关谷假单胞菌培养筛选出一种生物乳化剂,能乳化石油和柴油等,该菌在生物技术应用上得到越来越多的关注。综合以上信息,由于谷关假单胞菌也是假单胞菌属的成员之一,也可能编码脂肪酶。基于此,本研究尝试利用已知的门多萨假单胞菌(Pseudomonasmendocina)脂肪酶蛋白序列,本研究通过对谷关假单胞菌的基因组进行同源基因搜索获得了新的脂肪酶基因,丰富了低温脂肪酶基因资源。进而通过将该脂肪酶在大肠杆菌中重组表达并进行酶学性质的生化表征研究,发现获得的脂肪酶在低温下具有良好活性,表面活性剂对其活性有激活作用,且该酶在部分有机溶剂中高浓度处理时较低浓度下具有更高的酶活,本研究结果可为该脂肪酶在工业中的应用提供有用信息。
1 材料与方法
1.1 仪器和试剂
1.1.1试验材料 大肠杆菌(Escherichiacoli)Trans T1和Transetta(DE3)菌株均购于北京全式金生物科技有限公司,分别用于基因克隆、质粒扩增以及蛋白的重组表达。
1.1.2主要试剂 限制性内切酶购自TaKaRa公司;T4 DNA连接酶购自New England Biolabs公司;DNA提取试剂盒、质粒小量提取试剂盒和DNA胶回收试剂盒购自OMEGA公司;蛋白定量试剂盒购自北京全式金公司;蛋白Marker购于北京GeneStar生物公司;p-nitrophenyl acetate(pNPA)、p-nitrophenyl butyrate(pNPB)、p-nitrophenyl caprat(pNPC)、p-nitrophenyl laurate(pNPL)、p-nitrophenyl myristate(pNPM)、p-nitrophenyl palmitate(pNPP)和dimethyl sulfoxide(DMSO)购自Sigma-Aldrich公司;其他未列出试剂均为分析纯。
1.1.3主要仪器 T100TM普通PCR仪和凝胶成像系统,购自美国Bio-Rad公司;Synergy H1酶标仪购自美国Thermo公司;himac CR-GII高速冷冻离心机购自日本HIMAC公司。
1.2 试验方法
1.2.1脂肪酶PgLip1基因的筛选、克隆和载体构建 使用RAST在线服务器(http://rast.nmpdr.org/)对谷关假单胞菌的基因组序列(NZ_FNJJ01000004.1)进行自动注释。因来源于门多萨假单胞菌的脂肪酶性质已经得到较好的表征且已在生产中得到应用[20],将其氨基酸序列在RAST服务器中对谷关假单胞菌的基因组序列进行BlastP分析,得到氨基酸序列一致性为92.6%,将该基因命名为Pseudomonasguguanensislipase1(PgLip1)。采用SignalP(http://www.cbs.dtu.dk/services/SignalP/)预测所编码脂肪酶的信号肽。PgLip1的理论蛋白质分子量以及等电点采用ExPASy-ProParam(https://web.expasy.org/protparam)来预测。
将PgLip1基因的密码子根据大肠杆菌的偏好性进行序列优化,送金斯瑞公司合成基因,使用NdeI和XbaI限制性内切酶切割所获得的基因,与pET-28a(+)质粒进行连接,将连接产物转化到大肠杆菌Trans T1感受态细胞中,涂布于含有50 μg·mL-1卡那霉素的LB平板上,37 ℃培养过夜,挑取克隆后并进行PCR鉴定。将所获得的阳性克隆在含50 μg·mL-1卡那霉素的LB培养基中培养,提取质粒并送测序,筛选阳性质粒pET-PgLip1。
1.2.2PgLip1酶的表达和纯化 将测序验证正确的重组质粒pET-PgLip1转入大肠杆菌表达宿主细胞 Transetta(DE3)中,并将菌液涂布在同时含有50 μg·mL-1卡那霉素和25 μg·mL-1氯霉素的LB平板上,37 ℃过夜培养,待菌落长出后挑取单克隆接种于50 mL LB液体培养基中培养。37 ℃、220 r·min-1振荡培养12 h,按2% 比例转接于300 mL LB培养基以扩大培养。37 ℃、220 r·min-1振荡培养2 h至OD600为0.5时,加入300 μL浓度1 mol·L-1的诱导剂异丙基-β-D-硫代半乳糖苷(IPTG),将培养温度降低至16 ℃,继续振荡培养16 h。诱导后的菌液10 000 r·min-1离心5 min,收集沉淀,将其重悬于pH 8.5的50 mmol·L-1Tris-HCl缓冲液中,使用超声破碎仪进行细胞破碎。10 000 r·min-1离心5 min,收集上清液。将上清液缓慢流加至预先平衡的镍柱,用含20~400 mmol·L-1梯度咪唑的50 mmol·L-1Tris-HCl缓冲液清洗非特异结合蛋白,最后用含500 mmol·L-1咪唑的50 mmol·L-1Tris-HCl缓冲液洗脱目的蛋白。将洗脱蛋白用SDS-PAGE凝胶电泳检验,并对不含咪唑的50 mmol·L-1Tris-HCl缓冲液进行透析,获得纯化并脱盐的重组脂肪酶。
1.2.3PgLip1的活性测定 使用牛血清蛋白为标准蛋白,采用Bardford法[21]准确定量PgLip1的蛋白浓度。脂肪酶的活性测定采用比色法,底物采用pNPM,脂肪酶水解pNPM释放出pNP,通过测定410 nm的吸光度以判定pNP的释放量。每组试验设定3个重复。在37 ℃ pH 8.0的条件下以每分钟释放出1 μmolpNP所需的酶量为一个酶活力单位(U)。
1.2.4PgLip1的酶学性质研究 ①最适底物。将含有不同链长的对硝基苯酚酯类底物pNPA、pNPB、pNPC、pNPL、pNPM和pNPP(脂肪链分别含有2、4、8、12、14和16个碳原子)均配置成3 mg·mL-1的底物浓度,与PgLip1在37 ℃、pH 8.5的50 mmol·L-1Tris-HCl缓冲液中孵育15 min,以不加脂肪酶的反应作为阴性对照,通过pNP的释放,判定该酶的最适底物。
②最适pH和pH稳定性。配制具有不同pH的缓冲液(pH 2.0~3.0:甘氨酸-盐酸;pH 3.0~8.0:柠檬酸-磷酸氢二钠;pH 8.0~9.0:Tris-HCl;pH 9.0~12:甘氨酸-氢氧化钠),设置pH 2、3、4、5、6、7、8、8.5、9、10、11、12共12个处理,浓度均为50 mmol·L-1。将0.4 μmol·L-1的PgLip1和0.084 mmol·L-1的pNPM底物,在上述不同pH的缓冲液中于37 ℃孵育15 min,测定催化底物生成pNP的能力。以酶活最高的pH作为最适pH并将此时的酶活设为100%;将其他pH条件下的酶活除以此酶活作为相对酶活。将0.4 μmol·L-1的PgLip1酶液在不同pH下37 ℃处理1 h,在最适条件下以同样底物测定剩余酶活。以未孵育处理的酶作为对照并将其酶活设为100%,测定并计算酶在不同pH下的残余相对活力。
③最适温度和热稳定性。设置10、20、30、40、50 ℃共5个温度,将0.4 μmol·L-1的PgLip1在最适pH(pH 8.5)、不同温度和0.084 mmol·L-1的pNPM底物孵育反应15 min,测定pNP的释放,从而判定最适温度。将酶活最高的反应温度下的酶活力设定为100%,将其他温度下酶活除以此酶活获得相对酶活。
将0.4 μmol·L-1的酶液分别在不同温度(40~60 ℃)下保温,在不同时间点取样,在最适温度和最适pH条件下同样以pNPM底物测定残余酶活。以未进行热处理的脂肪酶活作为100%,将处理后的残余酶活除以此酶活获得不同温度、不同时间的相对酶活,用以检测酶的热稳定性。
1.2.5金属离子对PgLip1活性的影响 选择Ca2+、Cu2+、Mg2+、Fe3+、Mn2+、Zn2+、Pb2+、Ag+、Cr3+、K+、Ni+和Co2+12种金属离子,将0.4 μmol·L-1的PgLip1分别与5 mmol·L-1的不同金属离子在pH 8.5、37 ℃孵育30 min,在最适pH和最适温度下将酶和0.084 mmol·L-1pNPM底物孵育测定酶的相对活力。以未加金属离子的反应作为对照,将该酶活设定为100%,计算不同金属离子处理的的相对酶活。
1.2.6表面活性剂、变性剂以及EDTA对PgLip1活性影响 选择1%的Triton X-100、吐温-20和吐温-80三种表面活性剂,1%β-巯基乙醇和0.1% SDS两种变性剂以及5 mmol·L-1EDTA,将0.4 μmol·L-1的PgLip1分别加入到以上不同溶液中,37 ℃孵育1 h,在最适pH 和最适温度条件下以0.084 mmol·L-1的pNPM作为底物测定酶活。以未用上述任何试剂处理的酶液作为对照,将该反应的酶活力设定为100%,计算判定表面活性剂、变性剂和EDTA对脂肪酶的影响。
1.2.7有机试剂对PgLip1活性影响 选择丙酮、甲醇、乙醇、异戊醇、乙腈、异丙醇和乙酸乙酯共7种有机溶剂,将0.4 μmol·L-1的PgLip1酶分别加入10%、30%、50%终浓度的上述有机溶剂,在30 ℃下孵育1 h,在最适pH和最佳温度条件下以0.084 mmol·L-1的pNPM作为底物测定脂肪酶的酶活。以未加有机试剂的反应作为对照,将该反应的酶活力设定为100%,计算不同有机溶剂处理的相对酶活。
1.3 数据统计与分析
根据试验数据平均值,采用SPSS 19.0软件中的One-way ANOVA过程进行单因素方差分析,LSD和Duncan氏法进行各组间多重比较。
2 结果与分析
2.1 脂肪酶PgLip1基因的克隆和表达纯化
对PgLip1基因分析发现,该基因全长840 bp,预测编码一个脂肪酶PgLip1,长度为279个氨基酸,与来源于门多萨假单胞菌的脂肪酶有92.6% 的相似性。SignalP分析发现,其N端22 个氨基酸为信号肽序列。去除信号肽的成熟脂肪酶PgLip1由257个氨基酸残基组成,理论蛋白质分子质量27.4 kDa,理论等电点pI为9.0。重组PgLip1脂肪酶的SDS-PAGE电泳结果(图1)显示,其表观分子量为27.4 kDa,与预期蛋白分子量大小基本一致。
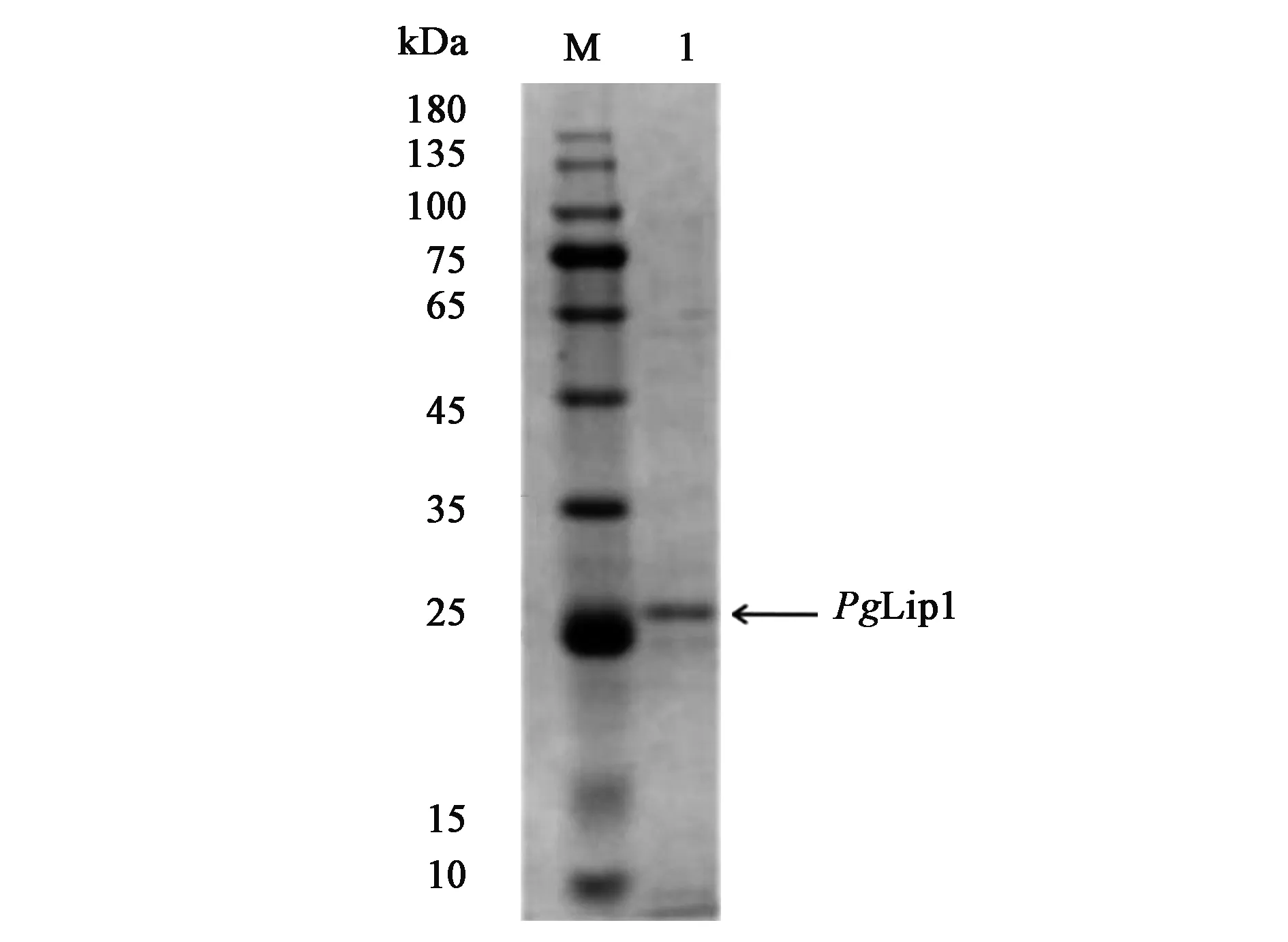
注:M—DNA分子量标准;1—纯化PgLip1蛋白。
2.2 PgLip1的酶学性质
2.2.1PgLip1的底物特异性 由图2可知,谷关假单胞菌来源的重组脂肪酶PgLip1对于具有不同长度脂肪链的pNP底物均具有一定活性,能催化本研究选择的含2~16个不同长度脂肪链的对硝基苯酚酯降解。该酶对pNPB的活性最高,比活达到27.5 U·mg-1,而对pNPA、pNPC、pNPL、pNPM和pNPP的比活分别为25.3、13.5、4.4、3.1和1.6 U·mg-1。对pNPB底物的PgLip1脂肪酶相对活力设定为100%,对碳链长度为2(pNPA)和8(pNPC)的pNP底物的酶活依次缓慢下降,其相对酶活分别为91.8%和49.1%;对碳链长度分别为12(pNPL)、14(pNPM)和16(pNPP)的pNP底物酶活则快速下降,其相对酶活分别为15.8%、11.1%和5.8%。
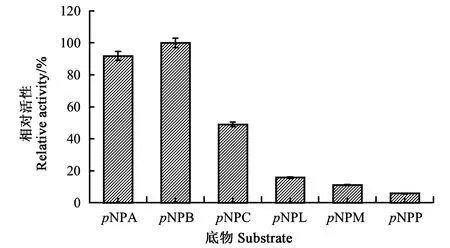
图2 PgLip1对pNP底物的相对酶活
2.2.2PgLip1的稳定性 以pNPM为底物时,pH对PgLip1脂肪酶活性的影响结果见图3,可见,在50 mmol·L-1pH为8.5的Tris-HCl缓冲液中,PgLip1的脂肪酶活性最高,其比活达到3.1 U·mg-1。当pH为8.0时,该酶的酶活为71.6%,而在pH为7.0时则迅速下降为4.0%。当pH为9.0时,该酶有84.4%的活性,而在pH为10.0仅剩余3.8%的活性。在pH稳定性方面,当pH为3.0~12时,PgLip1酶的相对活性至少保持在60%以上,说明该脂肪酶具有较宽的pH耐受性。但在pH 3.0以下,该酶的酶活几乎完全丧失。
由图3可以看出,以pNPM为底物时脂肪酶PgLip1在10 ℃时的活性最高,因此其催化水解的最适温度为10 ℃,此时酶活为4.2 U·mg-1,相对酶活为100%;但酶在0 ℃仍然具有60.5%的酶活,说明该酶是一个低温脂肪酶,能在低温下较好地降解底物。当孵育温度高于10 ℃时,该脂肪酶的酶活性开始逐渐下降。在20、30和40 ℃,其相对酶活分别为55.2%、42.1%和3.4%。将PgLip1脂肪酶在不同温度分别孵育处理,发现该酶在40 ℃处理30 min仍然能保持93.0%相对酶活,继续孵育至45 min其残余酶活为72.1%。在50 ℃处理5 min后,残余酶活为61.1%,继续延长时间残余酶活基本不变。60 ℃处理,酶活则迅速下降,30 min后酶活已基本完全丧失(7.1%)。
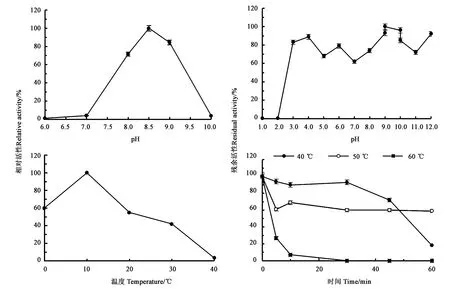
图3 PgLip1的生化性质分析
2.2.3金属离子对PgLip1活性的影响 以pNPM为底物时金属离子对PgLip1的影响结果见表1,可见,在含Pb2+、Co2+、Ca2+、Cu2+和Na+的溶液中,PgLip1相对比较稳定,其活性分别为对照处理(酶活为2.9 U·mg-1,相对酶活100%)的102.8%、91.4%、97.5%、81.5%和78.7%。相反,Mg2+、Fe3+、K+、Cr3+、Zn2+、Ni+和Mn2+对脂肪酶PgLip1有抑制作用,在这些离子溶液中该酶的相对酶活分别为20.0%、48.7%、28.8%、37.5%、8.3%、10.9%和20.6%。
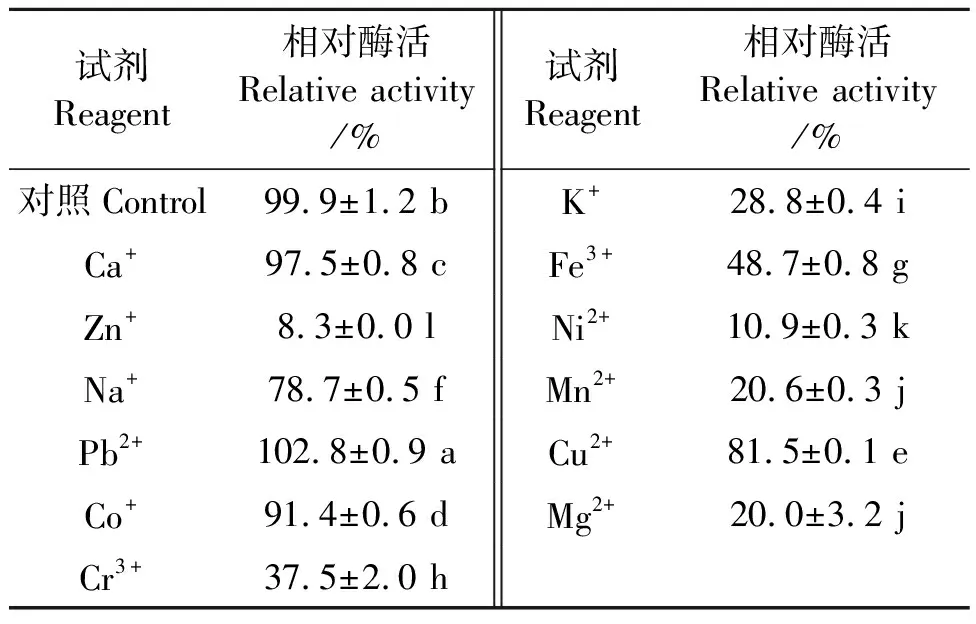
表1 金属离子对PgLip1活性的影响
2.2.4表面活性剂及EDTA等变性剂对PgLip1活性的影响 以pNPM为底物时表面活性剂及EDTA等变性剂对PgLip1的影响结果见表2,可知,表面活性剂吐温-20、吐温-80和Triton X-100对PgLip1都有一定的促进作用。在终浓度1%的表面活性剂吐温-20、吐温-80和Triton X-100的溶液中,PgLip1的酶活分别为对照处理(酶活为3.3 U·mg-1,相对酶活为100%)的316.7%、339.7%和589.0%。而当溶液中含有5 mmol·L-1的金属离子螯合剂EDTA、变性剂SDS和β-巯基乙醇时,PgLip1的相对酶活分别为184.6%、20.2%和102.4%。
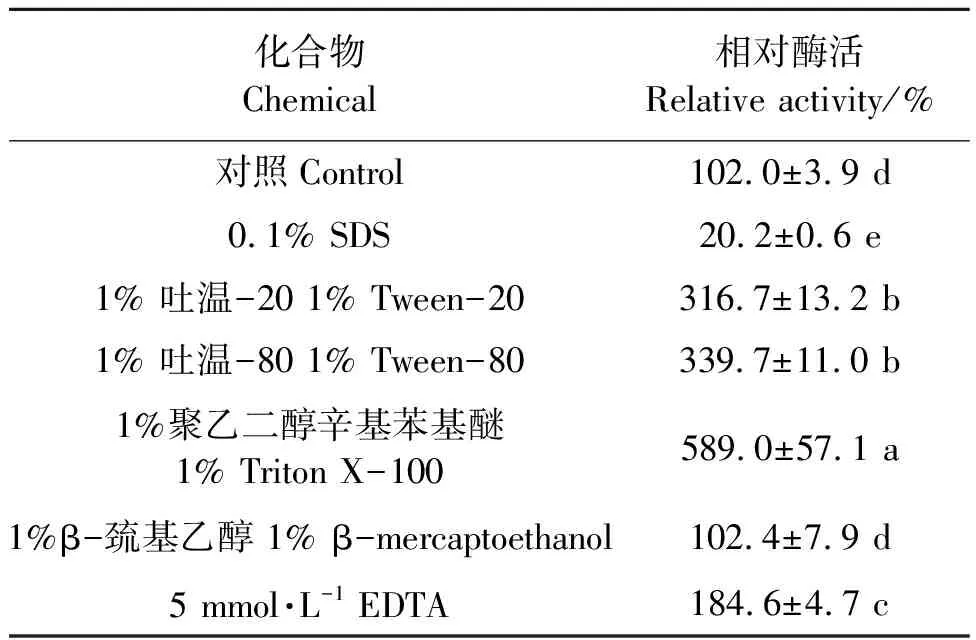
表2 表面活性剂、EDTA等变性剂对PgLip1活性的影响
2.2.5有机试剂对Pglip1活性的影响PgLip1在不同浓度不同有机试剂中的酶活结果见表3。未加有机溶剂的对照酶活为4.7 U·mg-1,相对酶活为100%。可以看出,PgLip1对50%的异丙醇和异戊醇有良好的稳定性。值得注意的是,在这两种有机试剂中,其剩余酶活分别随着有机试剂浓度的上升而增加,如在10%的异丙醇中酶活只有7.6%,但在30%的异丙醇中则为175.3%,而当异丙醇的浓度为50%时,酶活增加至240.3%。同样,在甲醇和乙醇中,虽然PgLip1的酶活均小于对照处理,但该酶的酶活也随着两种醇浓度的增加而上升。在丙酮、乙腈和乙酸乙酯中,PgLip1的酶活均有下降,且酶活并没有随着有机溶剂浓度的增加而上升。
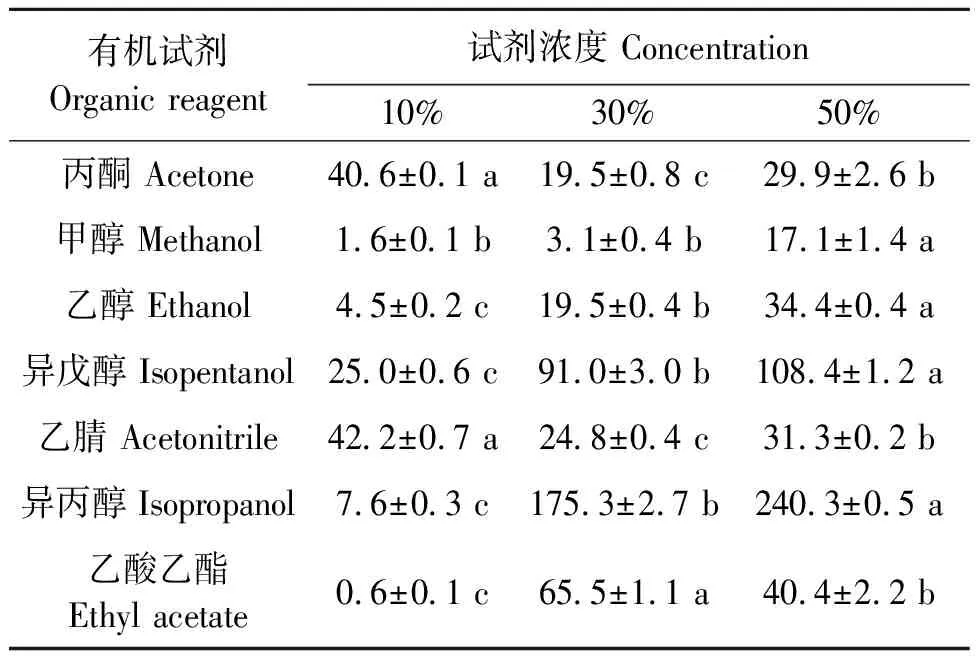
表3 不同浓度不同有机试剂对PgLip1活性的影响
3 讨论
本研究从谷关假单胞菌中克隆和重组表达了脂肪酶PgLip1,其最适培养温度为10 ℃。在0 ℃ 时,相对酶活为60%,当温度大于10 ℃ 时酶活性急剧下降。刘滔滔等[22]报道的碱性脂肪酶的酶活在35 ℃急剧下降,45 ℃基本丧失;孙苗苗等[23]报道的假单胞菌脂肪酶也具有热不稳定的性质,40 ℃酶活力快速失活,60 ℃酶活力基本丧失。Zhang等[24]筛选到一株Pseudomonassp.7323低温脂肪酶,其最适作用温度为30 ℃,在25 ℃以上显示出热不稳定性。PgLip1脂肪酶在40 ℃处理45 min能保持80%~100% 酶活力,50 ℃处理酶活力逐渐下降,60 ℃处理30 min酶活力丧失。可见,PgLip1脂肪酶具有热敏感特性。当温度较高时酶活性急剧下降,是低温酶的一个重要特征;而与中高温脂肪酶相比,低温脂肪酶有较低的活化能,在低温条件下具有高酶活[25],能维持低温条件下的高催化效率。PgLip脂肪酶具有更低的最适温度,同时在40、50 ℃仍具有相对较为耐热的特点。这一特点可能与P.guguanensis是从温泉中分离得到的有关[18],所以具有一定的耐热性。
据报道芽孢杆菌来源的脂肪酶在pH 6.0条件下处理60 min剩余50%酶活力[26]。曲威等[27]报道铜绿假单胞菌S8脂肪酶的最适pH为7.0,在pH 6.0~8.0时相对稳定,pH 4.0时剩余酶活力为50%。余琼[28]报道假单胞菌(Pseudomonassp.)YT34-8脂肪酶在pH 5.0~10.0稳定,但在pH 4.0和11.0条件下,剩余酶活力不足60%。本研究报道的PgLip1的最适pH为8.5,当pH低于8.0或高于9.0时,酶活性明显降低。在pH 3.0~12.0时,酶相对活性仍然保持在60%以上;在pH 9.0~10.0 时,酶活力较为稳定;而当pH低于3.0的极酸性环境下酶活才出现稳定性的明显下降。因此,稳定的pH耐受性使PgLip1脂肪酶具有更好的应用价值、更广的应用范围。
PgLip1脂肪酶对含2、4和8个碳原子脂肪链的对硝基苯酚酯底物具有较高的催化活性,催化短链酯的活性高于含12、14和16个碳原子长链脂肪酯的活性。虽然部分脂肪酶优先水解14碳以上的长链脂肪酸,但是低温脂肪酶对含有3~10碳链甘油脂的催化效率更高,这可能与低温脂肪酶的催化特性有关[29-31]。有文献报道圆弧青霉(Penicilliumcyclopium)脂肪酶、PseudomonasfluorescensPf0-1来源脂肪酶对短链(8碳以下)对硝基苯酚脂具有较高的特异性[32],而也有报道[22]一株产碱假单胞菌碱性脂肪酶对8碳pNP底物具有较高活性,脂肪酶对短链酯的催化能力反映了脂肪酶丰富的生物多样性。
金属是影响酶活性的重要因素,对不同的酶,金属和重金属离子可能分别具有促进或抑制脂肪酶活力的效果[33]。本研究中,Cu2+、Co2+、Na+、Ca2+和Pb2+金属离子处理后,PgLip1保持相对稳定性,酶活达到79%以上。在金属离子中的稳定性,使PgLip1在工业领域可以具有良好的应用前景[34]。
脂肪酶PgLip1在浓度高达50% 的异丙醇和异戊醇有机溶剂中具有良好的稳定性,其剩余酶活分别为240.3%和108.4%。据报道JXJ-54脂肪酶在三氯甲烷和乙醇中,酶活力随着有机溶剂体积分数的增大而增强,而在甲醇、乙醇、丙酮、异丙醇四种有机试剂中,酶的活性随着有机试剂浓度的增加而减小,在50%的乙醇和丙酮中,剩余酶活仅有5.3%和15.3%[35]。从有机试剂对脂肪酶PgLip1活性的影响结果来看,不同浓度的有机试剂对PgLip1脂肪酶活性的影响不同,其原因可能与有机溶剂的lgP值有关[35]。脂肪酶的结构通常是整体亲水,但其具有疏水的核心和亲水的表面,其活性必须由具有一定极性的溶剂来维持,而有机溶剂则可能参与维持了脂肪酶核心的稳定性。当溶剂极性过低时,酶的结构最终会被破坏,导致活性降低甚至失活[36]。脂肪酶PgLip1在有机溶剂中呈现的稳定性,使其在基于非水相的催化反应领域中具有较大的应用潜力,比如食品、油脂、新型材料、精细化工、医药、化妆品和生物能源等[37]。
此外,PgLip1在吐温-20、吐温-80和Triton X-100等表面活性剂中都被激活,在Triton X-100中激活幅度甚至达到5.9倍。但相同浓度的表面活性剂,Triton X-100激活效果显著优于吐温-80和吐温-20。刘滔滔等[22]报道的假单胞菌脂肪酶受到吐温-20、吐温-80和TritionX-100强烈的抑制,而Liu等[32]报道PseudomonasfluorescensPf0-1来源的脂肪酶在TritonX-100、吐温-20、吐温-40和吐温-80中的相对酶活达到110%、170%、160%、150%。因此,PgLip1受表面活性剂的激活程度优于这些报道的脂肪酶。PgLip1受表面活性剂激活的原理在于表面活性剂具有双亲性,一端为亲水基团,另一端则为疏水基团,它可以帮助维持酶的稳定性。Carrasco-López等[38]指出,非离子型表面活性剂在低浓度时可以与脂肪酶的疏水活性中心产生相互作用,帮助脂肪酶稳定在开放的构象,有利于底物进入活性中心以及产物的释放,因此脂肪酶的活力得到提高。但对于阴离子表面活性剂SDS,其为常见的蛋白质变性剂,其分子上的硫酸基团与蛋白质表面的带正电氨基酸可以紧密结合,而分子上的烷基则可以与蛋白质的疏水部分紧密结合,从而导致蛋白质发生变性[39]。
综上所述,本研究从谷关假单胞菌中克隆得到一个新的低温脂肪酶基因PgLip1,该酶具有嗜低温特性、较好的耐温性、宽泛的pH耐受性、对有机溶剂的耐受性以及受表面活性剂激活的特殊性质,这丰富了人们对脂肪酶生物多样性的认识;同时,PgLip1还可能在饲料行业如水产饲料以及工业应用如洗涤剂、有机合成等领域存在着潜在的应用价值。
猜你喜欢
中华胰腺病杂志(2022年4期)2022-08-23
当代水产(2022年3期)2022-04-26
河南工业大学学报(自然科学版)(2021年6期)2022-01-26
感染、炎症、修复(2021年1期)2021-07-28
教育周报·教育论坛(2020年3期)2020-10-21
求知导刊(2019年15期)2019-08-30
华东师范大学学报(自然科学版)(2019年3期)2019-06-24
科技资讯(2018年16期)2018-10-26
科技信息·下旬刊(2018年8期)2018-10-21
广西科技大学学报(2018年2期)2018-09-10
