
基于酯酶同工酶的暗褐网柄牛肝菌遗传多样性分析*
2021-04-09 07:55何明霞方艺伟杨天伟许欣景王文兵张春霞
中国食用菌 2021年2期
高 锋,何明霞,刘 静,方艺伟,杨天伟,许欣景,王文兵,王 云,张春霞
(云南省热带作物科学研究所,云南 景洪 666100)
暗褐网柄牛肝菌(Phlebopus portentosus) 是牛肝菌目(Boletales) 小牛肝菌科(Boletinellaceae) 脉柄牛肝菌属(Phlebopus) 的一种珍稀食用菌,俗称黑牛肝菌,国内主要分布于云南、广西、海南和四川;国外主要分布于泰国和斯里兰卡[1]。同时,暗褐网柄牛肝菌是西双版纳市场上深受人们喜爱的一种有营养,经济价值高,且味道鲜美的珍稀食用菌[2]。
野生暗褐网柄牛肝菌由于受到掠夺性地采收,市场供应量逐年递减。2003年以来系统地进行了暗褐网柄牛肝菌的栽培技术研究[2-4],目前其人工栽培专利已成功转让给企业进行工厂化周年栽培[5-7];仿生栽培技术也相继获得成功[8-9],并已向部分农户推广。无论是工厂化栽培还是仿生栽培,目前所用的优良菌株主要是通过组织分离野生及仿生栽培的子实体,再经过出菇试验筛选获得[10-11]。由于优良菌株传代一定次数后会发生退化[19],因此科学有效地保护暗褐网柄牛肝菌野生资源对该产业的健康可持续发展至关重要。
同工酶是催化相同反应而结构和理化性质不同的一类酶[12],同工酶分析是从蛋白质水平上研究生物群体遗传分化的重要手段之一[12-14]。对酯酶同工酶酶谱的相似性系数进行聚类分析已广泛用于研究食用菌菌株的亲缘关系和遗传分化[15-16],也用于选择杂交亲本、预测杂交优势[17]和鉴定杂交菌株[18]的研究。为了系统地评估暗褐网柄牛肝菌的遗传多样性,从云南、四川两省的9个采样地共收集31个暗褐网柄牛肝菌子实体并通过组织分离获得了其菌株,分别利用固体培养和液体培养2种方式获得其菌丝体,并对其进行酯酶同工酶分析,通过酶谱聚类分析研究分离菌株的遗传多样性。
1 材料与方法
1.1 子实体采集
野生子实体采集信息见表1,来源于同一采集地子实体的组织分离菌株为一个群体。
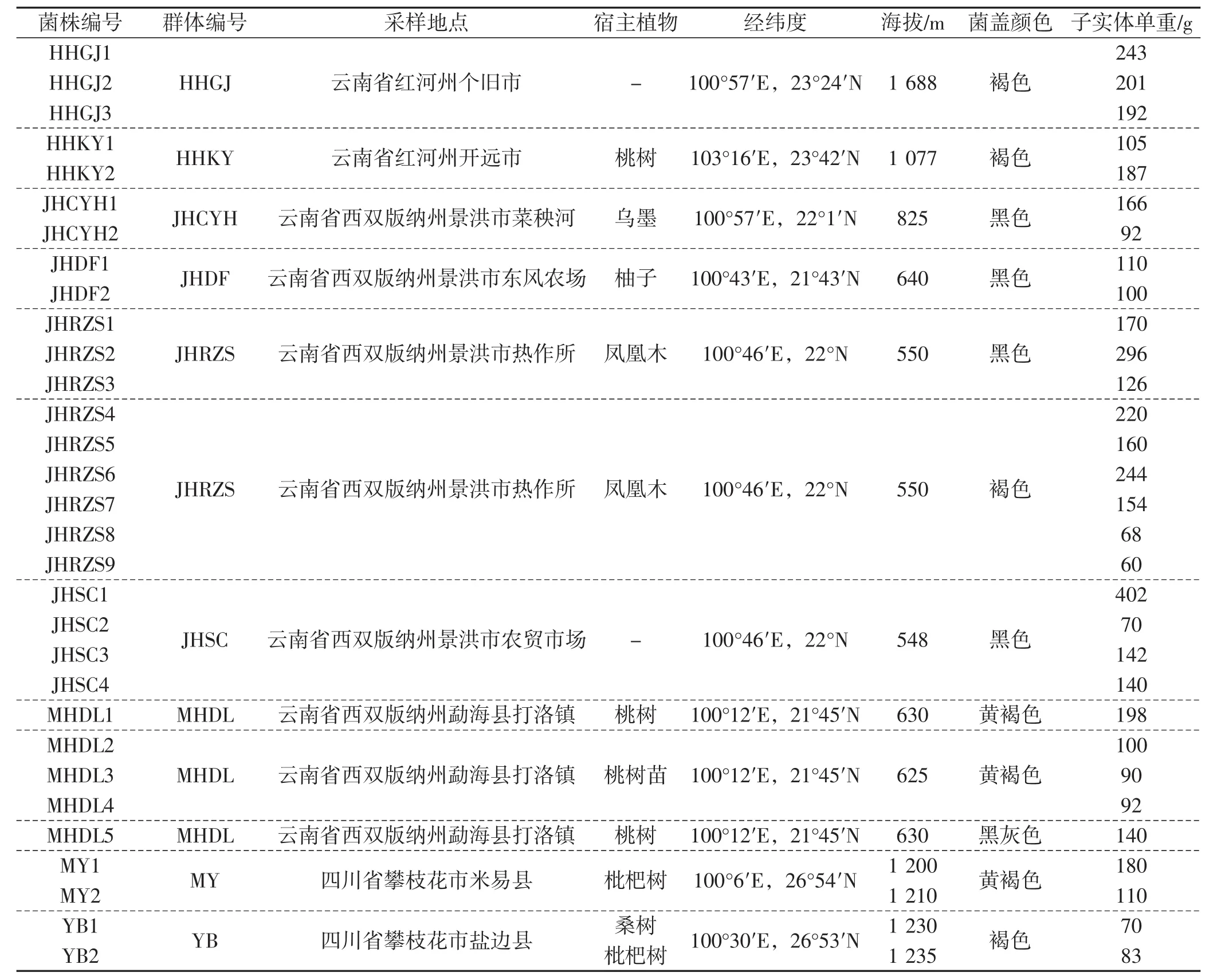
表1 子实体采集信息Tab.1 Information of sampled Phlebopus portentosus isolates
1.2 培养基
M1液体培养基:去皮马铃薯(煮汁) 200 g、葡萄糖 20 g、酵母膏 2 g、MgSO4·7H2O 1 g、KH2PO41 g,蒸馏水定容至1 L[19]。
M1固体培养基:去皮马铃薯(煮汁) 200 g、葡萄糖 20 g、酵母膏 2 g、MgSO4·7H2O 1 g、KH2PO41 g、琼脂粉15 g,蒸馏水定容至1 L[19]。
1.3 试剂
Blue Plus®Protein Marker(1.4×104~1.0×105),北京全式金生物技术有限公司;Precast-Glgel 10%预制胶、改良型Bradford蛋白浓度测定试剂盒及其他分析纯试剂,上海生工生物工程股份有限公司。
1.4 仪器
BIC-250人工气候箱,上海博迅实业有限公司;SK-100B恒温摇床,上海苏坤实业有限公司;DYCZ-24DN双垂直电泳仪,北京市六一仪器厂。
1.5 菌株分离和保藏
选取新鲜、实心、菌管尚未开放的暗褐网柄牛肝菌子实体,切去菌柄底部的泥沙,在超净工作台上,挑取菌柄与菌盖交界部位约0.5 cm×0.5 cm组织块,置于试管中M1固体培养基斜面上,在28℃~30℃条件下避光培养30 d。切取约50 mm2菌丝块,置于装有5 m无菌蒸馏水的15 mL塑料管中,15℃避光保存于云南省热带作物科学研究所。
1.6 酯酶同工酶电泳
1.6.1 菌丝体培养
固体培养菌丝体:挑取菌丝块(直径1 cm)接种于M1固体培养基平板上,于30℃避光培养7 d,在菌丝尖端切取1个菌丝块(直径1 cm) 接种于另一个平板中央,30℃避光培养18 d。
液体培养菌丝体:挑取菌丝块(直径1 cm) 接种于M1固体培养基平板上,于30℃避光培养7 d,无菌操作用手术刀将菌丝尖端挑取的10个菌丝块(直径1 cm) 切碎,转至装有400 mL M1液体培养基的500 mL三角瓶中,30℃、120 r·min-1振荡培养7 d。
1.6.2 酯酶同工酶电泳
样品制备、电泳及染色分析参照高锋等[19-20]的方法。根据蛋白质Marker的相对分子质量及各条带的相对迁移率,利用Microsoft Excel 2019拟合出标准曲线,用AlphaView SA3.4.0软件计算各菌株酯酶同工酶的相对迁移率及相对分子质量。
1.7 聚类分析
为了分析菌株间的亲缘关系,参照Sneath等[21]的方法计算相似性系数(S),公式为:

式中:a为A菌株条带数;b为B菌株条带数;c为A菌株、B菌株共有条带数。
先用SPSS 20.0软件中的Jaccard相似性计算出菌株间的酯酶同工酶酶谱相似性系数(S)矩阵,再将该矩阵换算为距离(D=1-S) 矩阵,最后用MEGA X软件的类平均连锁法(UPGMA) 对各菌株酯酶同工酶酶谱进行聚类分析[22]。鉴于同一菌株2种培养方式的酯酶同工酶酶谱存在较大差异,单一培养方式有部分酯酶同工酶未诱导出,因此,将2种培养方式的酶谱合并后进行聚类分析[22]。
2 结果与分析
2.1 酯酶同工酶电泳结果
部分液体及固体培养的菌丝体酯酶同工酶电泳图见图1、图2。暗褐网柄牛肝菌酯酶同工酶条带统计见表2。
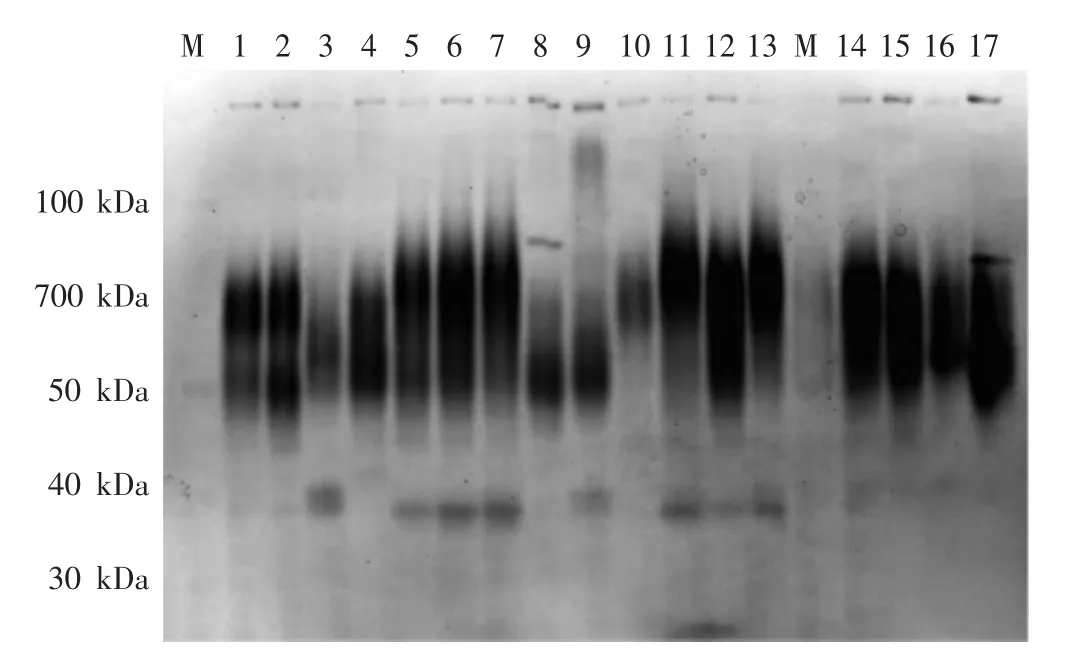
图1 部分液体培养菌丝体酯酶同工酶电泳图Fig.1 Electrophoretic zymogram of esterase isozymes of mycelia cultured in liquid medium
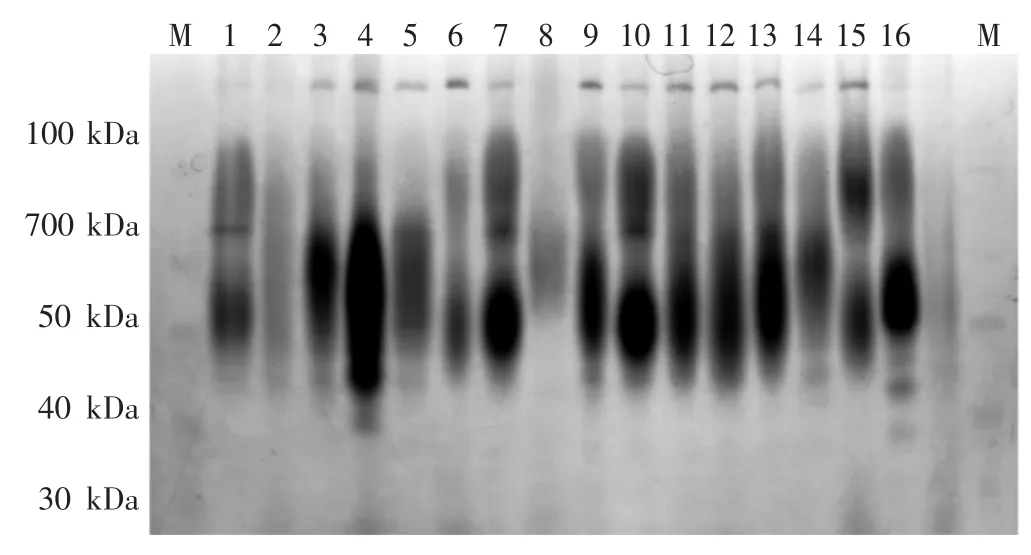
图2 部分固体培养菌丝体酯酶同工酶电泳图Fig.2 Electrophoretic zymogram of esterase isozymes of mycelia cultured on solid medium
如图1、图2所示,暗褐网柄牛肝菌菌株酯酶同工酶条带类型有4种:深色宽、深色窄、浅色宽和浅色窄;根据蛋白Marker确定各条带的相对分子质量及相对迁移率,拟合出标准曲线(R2=0.997)方程为:
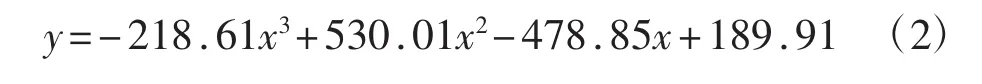
式中:y为相对分子质量;x为相对迁移率(Rf)。
如表2所示,31个暗褐网柄牛肝菌菌株固体培养的菌丝体共检出10条酯酶同工酶条带,其中,Rf为 0.13、0.48、0.55和0.65的条带出现频率较高,各菌株分别具有2条~5条酯酶同工酶条带,共检出21种酶谱类型;液体培养的菌丝体共检出11条酯酶同工酶条带,Rf为0.13、0.55、0.38、0.65和0.75的条带出现频率较高,各菌株分别具有2条~6条酯酶同工酶条带,共检出23种酶谱类型。2种培养方式下所有菌株均检出一条Rf为0.13、相对分子质量为136 kDa的酯酶同工酶条带。不同培养方式酯酶同工酶酶谱相似性系数见表3。

表3 不同培养方式酯酶同工酶酶谱相似性系数Tab.3 Similarity coefficient of esterase isozymes in different medium of the same strain
如表3所示,不同培养方式下,同一菌株的酶谱也存在不同程度的差异,相似性系数介于0.143~1.000,除了MHDL5、HHKY1和JHSC1的相似性系数大于等于0.500外,其他28个菌株的相似性系数均小于0.500。31个菌株中,液体培养菌株的酯酶同工酶条带数与固体培养的比值大于1的菌株有19个,等于1的有8个,小于1的有4个,可见液体培养菌株的酯酶同工酶条带数普遍多于固体培养。
2.2 酯酶同工酶酶谱聚类分析结果
2.2.1 液体培养菌株聚类分析结果
液体培养菌株酯酶同工酶酶谱聚类结果见图3。
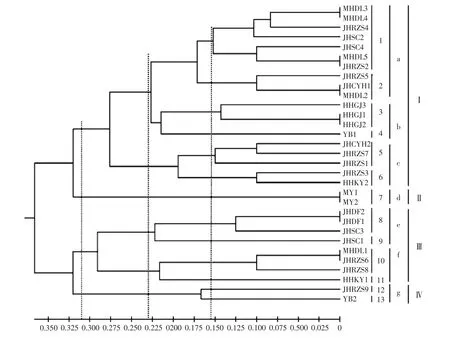
图3 液体培养菌株酯酶同工酶酶谱聚类结果Fig.3 Cluster analysis of esterase isozyme zymogram of mycelia cultured in liquid medium
如图3所示,31个暗褐网柄牛肝菌菌株液体培养菌丝体的酶谱相似性系数为0.111~1.000,遗传距离为0~0.889。31个菌株在遗传距离为0.150、0.220和 0.290时,分别聚为 13(1~13) 类、7(a~g) 类和4(I~IV) 类。在遗传距离为0.150时,除HHGJ、JHDF、MY群体的菌株分别聚在同一分支外,其他群体的菌株聚在不同分支。
2.2.2 固体培养菌株聚类分析结果
固体培养菌株酯酶同工酶酶谱聚类结果见图4。
如图4所示,31个暗褐网柄牛肝菌菌株固体培养的菌丝体酶谱相似性系数为0.125~1.000,遗传距离为0~0.875。31个菌株在遗传距离为0.130、0.225和0.300时,分别聚为16(1~16)类、9(a~i)类和4(I~IV) 类。在遗传距离为0.130时,只有MY群体的2个菌株聚在同一分支,其他群体的菌株聚在不同分支。
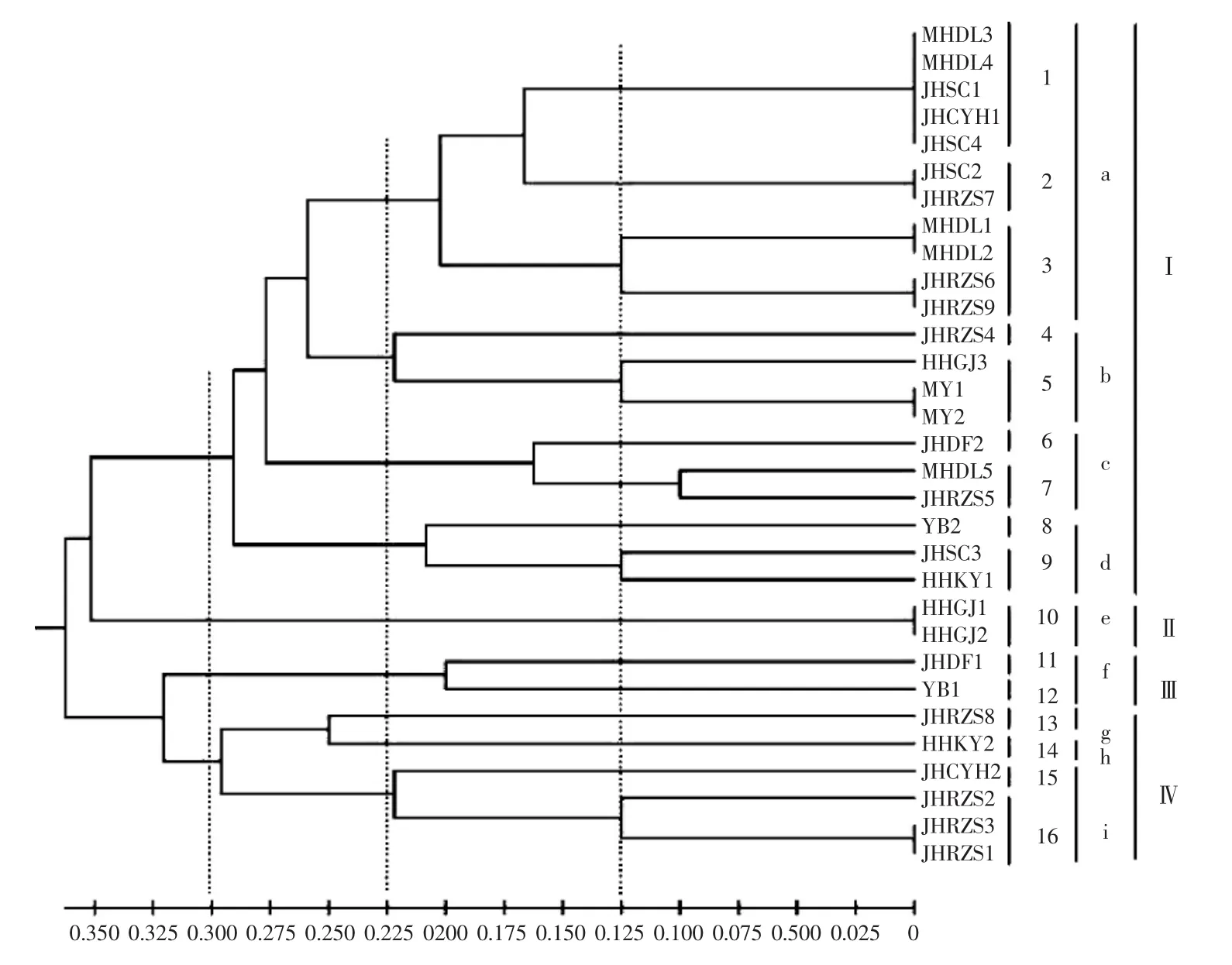
图4 固体培养菌株酯酶同工酶酶谱聚类结果Fig.4 Cluster analysis of esterase isozyme zymogram of mycelia cultured on solid medium
2.2.3 两种培养方式菌株联合聚类分析结果
2种培养方式菌株酯酶同工酶酶谱联合聚类结果见图5。
如图5所示,2种培养方式菌株的酶谱进行联合聚类分析的相似性系数为0.083~1.000,遗传距离为0~0.917。31个菌株在遗传距离为0.160、0.245和 0.280时,分别聚为 13(1~13) 类、7(a~g) 类和4(I~IV) 类。在遗传距离为0.160时,只有MY群体的2个菌株聚在同一分支,其他群体的菌株聚在不同分支。
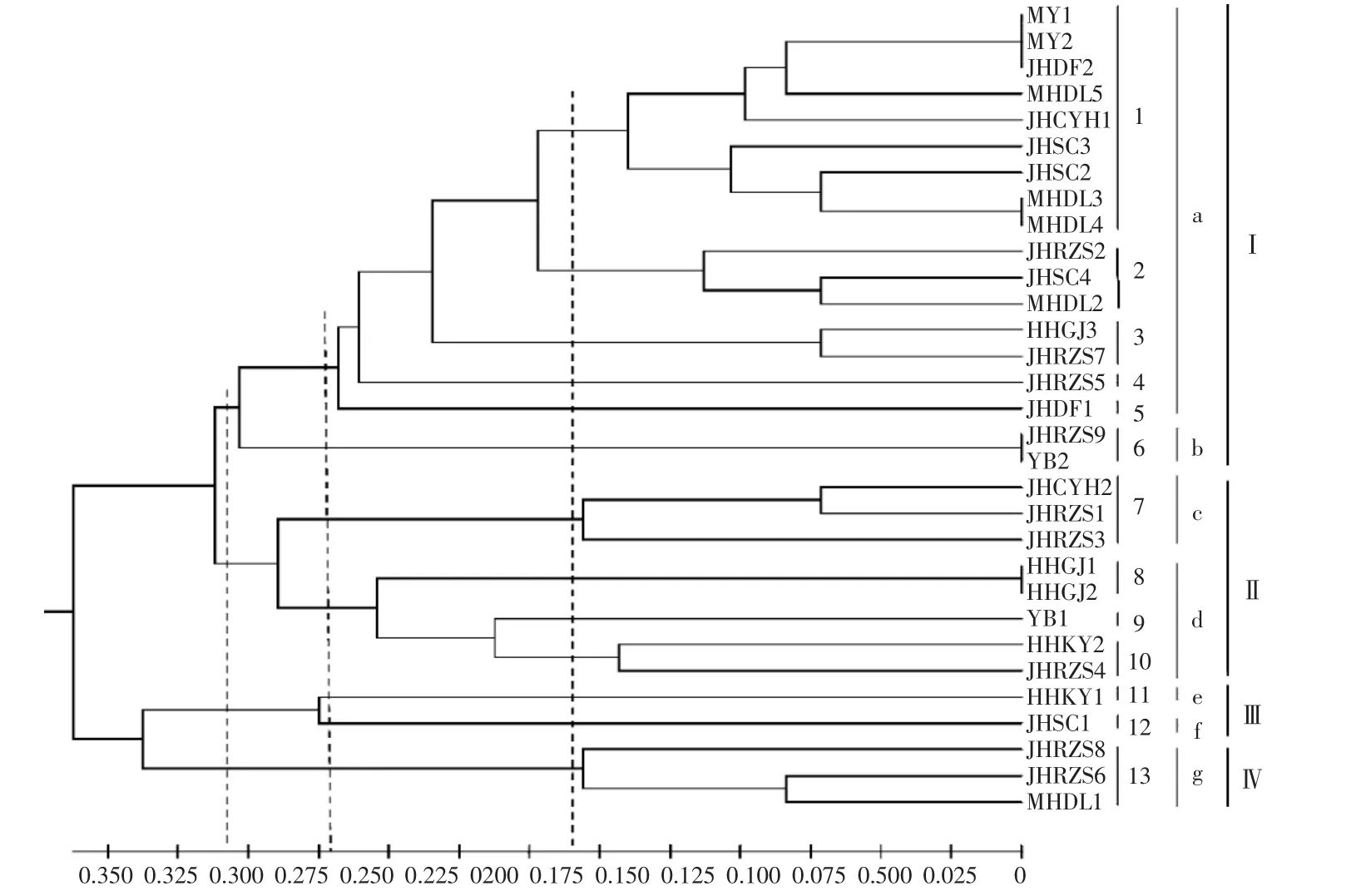
图5 两种培养方式菌株酯酶同工酶酶谱联合聚类结果Fig.5 Combination cluster analysis of esterase isozyme zymogram of solid and liquid media cultured mycelium
3 讨论
多项研究利用酯酶同工酶技术分析野生食用菌菌株的遗传多样性以及与地理分布的相关性。王一等[23]研究发现来源于川西高原的113个蘑菇(Agaricus campestris) 菌株在相似系数为0.50时,分为5大类群,但聚类结果与采集地的地理位置相关性不强。郭金英等[24]研究发现采自神农架的40个野生香菇(Lentinula edodes) 菌株分别具有5条~8条酯酶同工酶条带,在遗传相似系数约0.71时,分为7个类群。翟玉[25]研究发现,除个别菌株外,采自云南省及周边的黑木耳(Auricularia auricular)菌株与东北地区的菌株分别聚在不同的分支上。吴圣进等[16]研究发现广西百色地区采集的23个野生黑木耳菌株在遗传相似系数为0.68时,分为两大类群,大部分菌株聚类结果与地理位置相关。本研究发现基于酯酶同工酶酶谱的暗褐网柄牛肝菌菌株聚类结果与地理来源无相关性,这与多年的生态调查相吻合。暗褐网柄牛肝菌在国内主要分布在公园、果园、苗圃中,在原始森林中极少发现,推测其可能随着宿主植物的引种、移栽发生了人为传播,因此各地域间的菌株未能形成有效的地理隔离。
研究表明,培养时间和培养条件对食用菌酯酶同工酶的表达具有一定的影响。梁建光等[26]分别从培养第7天、第11天、第14天的糙皮侧耳(Pleurotus ostreatus) 菌丝体中检出10条、15条、13条酯酶同工酶条带;在4种不同培养基(I~IV) 培养的菌丝体中分别检出5条、6条、11条、3条酯酶同工酶条带,其中培养基III与IV培养的菌丝体之间的酶谱相似性系数仅为0.273。吴康云等[27]从培养10 d的黑木耳单核菌株J3和H2中分别检出3条、6条酯酶同工酶酶条带,培养30 d时2个菌株均缺少Rf为0.673的条带。李守勉等[28]发现刺芹侧耳(Pleurotus eryngii)2号菌株在培养至第5天时具有Rf为0.200和0.345的2个酯酶同工酶条带,第7天时增加Rf为0.455的条带,9 d~15 d时增加Rf为0.873和0.945的2条条带。宗立立等[29]从PDA液体培养基培养的滑菇(Pholiota nameko) C31菌株菌丝体中检出8个酯酶同工酶条带,而PDA固体培养基培养时只检出7条,两者相似性系数为0.875。本研究中菌丝体液体培养时间为7 d,固体培养时间为18 d,相同菌株不同培养方式的酶谱相似性系数均小于0.500,2种培养方式酶谱的联合聚类分析结果表明暗褐网柄牛肝菌具有丰富的遗传多样性。近年来,分子生物学技术广泛用于食用菌遗传多样性研究[30-32],后续试验将进一步增加暗褐网柄牛肝菌采集地数量,尽可能覆盖其主要分布区域并适当增加分离菌株数,利用SSR、ISSR、RAPD、ITS等分子标记研究其遗传多样性,为后续种质资源保护、种性鉴定、优良菌株选育及菌种管理等提供参考。
猜你喜欢
计算机应用与软件(2022年6期)2022-07-12
自然灾害学报(2022年2期)2022-05-10
文萃报·周五版(2021年30期)2021-09-05
食品安全导刊·中旬刊(2021年6期)2021-08-06
食品安全导刊(2021年17期)2021-07-09
中国学校体育(2021年10期)2021-04-26
江苏农业科学(2016年7期)2016-10-20
饮食与健康·下旬刊(2016年9期)2016-05-14
江苏农业科学(2016年3期)2016-05-03
天津农业科学(2015年10期)2015-10-20
