
干旱复水对5种马尾松菌根化苗木生理特性的影响
2021-04-22 13:20涂桂英
中南林业科技大学学报 2021年4期
王 艺,涂桂英
(1.贵州民族大学 生态环境工程学院,贵州 贵阳 550025;2.贵州大学 林学院,贵州 贵阳 550025)
马尾松Pinus massonianaLamb.是中国南方地区最主要的用材树种、造林先锋树种[1]及产脂树种[2],也是典型的外生菌根树种[3],在我国松属乡土树种中分布最广,速生丰产,适应性强[4],具有良好的生态效益和经济价值,在我国林业中具有重要地位,同时对贵州省储备林建设也发挥着重要作用。
一直以来,干旱都是人们关注的一个重点问题,而干旱也严重影响着植物的生长和发育。现如今,干旱已成为我国,特别是西部地区林业建设的重要限制因子之一。可喜的是已有相关研究表明,菌根真菌可通过建立共生关系,扩大根系吸收面积,改善寄主植物的水分状况和养分代谢,避免或减缓水分胁迫对植物的伤害[5],促进植物生长发育。外生菌根真菌可增强苗木的抗旱能力[6],促进苗木对水分和营养物质的吸收[7],提高苗木的抗逆性和适应能力[8]。目前,虽然在其它植物上利用菌根提高水分利用效率和耐旱性方面已有较多研究[6,9-12],但在持续干旱条件下,研究外生菌根真菌对马尾松幼苗生长、水分代谢和渗透调节物质的影响,以及胁迫解除复水过程中马尾松菌根化苗木在生理生化物质上的变化,国内外尚少见报道。为此,本研究采用盆栽方法,探讨持续干旱及复水后外生菌根真菌对马尾松幼苗生长及生理特性的影响,为外生菌根真菌在马尾松抗旱上的应用提供理论依据。
1 材料与方法
1.1 试验材料
马尾松种子采自贵州省都匀市优良林分。本研究供试的5 种菌种由西南大学黄建国教授提供,分别为:牛肝菌(Sp7)(简称S7)、牛肝菌(Sp1)(简称S1)(二者都为褐环乳牛肝菌Suillus luteus)、鸡油菌Cantharellus cibarius(简称G)、彩色豆马勃Pisolithus tinctorius(简称C)、土生空团菌Cenococcum geophilum(简称T),不接菌为对照(CK)。
1.2 试验方法
1.2.1 菌剂制备及培养
平板培养5 个供试菌种。选用MMN 固体培养基,28℃培养10 d,菌丝长满后备用。液体扩大培养基质,采用改良PD 培养基。首先在量程为500 mL 的三角瓶中装入制备好的培养基,每个三角瓶中装入300 mL,接着在超净工作台内分别用直径1 cm 无菌打孔器切取已培养好的外生菌根菌平板菌落,接种到液体培养基中,每瓶装5 个菌片,封口。接完种后将其放置25℃培养箱中,约20 d,待菌丝长满基质后,作为菌剂使用[13]。
1.2.2 种子及土壤处理
选取籽粒饱满无虫害的马尾松种子,流水冲洗干净后,用75%乙醇表面消毒1 min,无菌水冲洗4 次,然后用初始温度45℃温水浸种24 h,湿纱布包裹,放到25℃恒温箱中催芽,待种子裂嘴露白时即可播种。试验基质是由第四纪红色黏土发育的黄壤与干净的河砂组成,按体积比9∶2比例混合,须在高压灭菌锅内(压力0.14 MPa,124~126℃)连续灭菌2 h 后备用。塑料花盆规格为250 mm×280 mm,用95%酒精擦拭后晾干。供试土壤的基本理化性质见表1。
1.2.3 播种和接种
每盆接入菌丝液1 000 mL。采用点播方式,接种采用“三层接种法”,即首先在试验盆中装入1/3 高度的灭菌基质,然后均匀地喷洒上一层菌丝液,用量大约是每盆接入量的1/3,之后又撒上一层基质至盆高的3/4 处,再喷洒1/3 的菌丝液,覆上一层基质,种子播于其上,最后将剩余的菌丝液喷洒于种子周围,覆土。
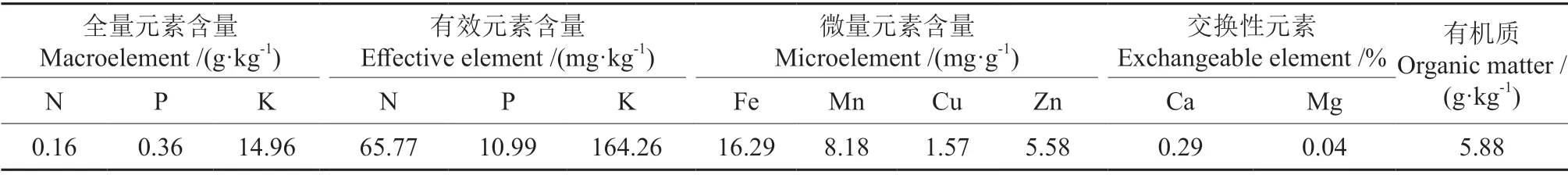
表1 供试土壤基本化学性质Table 1 Basic chemical properties of the soil used in the experimental
1.2.4 试验设计
采取随机区组试验,试验设5 个接种处理、1个对照处理(不接种),每处理25 盆,每盆4 株,每处理3 次重复。分别在持续干旱当天(0 d)和持续干旱第7 天、第14 天、第21 天、第28 天和第35 天进行相关指标的测定,同时在持续干旱的14、21、28 d 对部分苗木进行复水,并在复水7 d后进行取样调查,分别用复水1(R1:持续干旱14 d 后复水)、复水2(R2:持续干旱21 d 后复水)和复水3(R3:持续干旱28 d 后复水)表示。干旱程度进行分级[14]:湿润,RSC ≥70%;轻微干旱,55%≤RSC<70%;中等干旱,45%≤RSC<55%;严重干旱,35%≤RSC<45%;极端干旱,RSC<35%。
1.3 指标的测定
3月开始育苗,待苗木长至6 个月时,从大约200 株苗中随机抽取20 株进行侵染率调查,观察发现接种外生菌根真菌的苗木侵染率达100%,而未接种的苗木侵染率为0。开始水分胁迫后,每10 d 测定1 次苗高和地径,直至胁迫试验结束。将植株冲洗干净后,放置80℃烘箱烘至恒质量,称得干质量,测定生物量。用Epson 数字扫描仪(Expression 10000 XL1.0)对不同处理后的根系进行扫描,然后对根系形态指标进行定量分析。自由水、束缚水、叶片相对含水量、根系活力、蔗糖、可溶性糖、可溶性蛋白、脯氨酸的测定均参考张志良等[15]及郝建军等[16]的方法。
1.4 数据处理
选用Excel 2016 软件对数据进行处理及作图,运用SPSS 26.0 统计软件进行方差分析,并用Duncan 进行多重比较,采用二阶聚类分析法筛选出影响菌根抗旱的关键指标,并按重要程度从高到低依次排序。
2 结果与分析
2.1 持续干旱及复水对不同苗木生长的影响
2.1.1 持续干旱对不同苗木生长的影响
由表2可见,与CK 相比,接种的5 种外生菌根真菌均显著提高了苗木苗高、地径、生物量和根冠比(P<0.05)。其中在苗高方面,接种S1 增幅最大,达37.8%;S7 次之,达32.6%。在地径上,T 增幅最大,达39.9%;S1 次之,达36.5%。生物量增加方面效果最佳的是接种S1 和T,均高出对照的50%以上,其接种效果为:S1>T>C>G>S7。外生菌根真菌对苗木根系及地上部分生长有显著的促进作用,特别是根系生长,增幅最小达20.9%(C),最大达34.6%(S1)。可见,菌根化马尾松苗木根冠比显著高于CK。综上所述,接种S1 和T 对幼苗在生长上促进效果较好,且菌根依赖性最强。

表2 持续干旱对不同苗木生长的影响†Table 2 Effect of continuous drought to the growth of different seedling
2.1.2 持续干旱及复水对不同苗木干物质积累和根冠比的影响
由图1可知,苗木干物质积累随胁迫时间的延长呈先增后降的趋势,在胁迫21 d 后达到最大值,其中接种S1 的效果最好,与CK 相比增加了48.3%,菌根化苗木与CK 间存在显著差异(P<0.05)。复水后外生菌根真菌对苗木产生明显的激发作用,提高了菌根化苗木干物质积累量,且超过复水前。在持续干旱14、21、28 d 复水后(R1、R2、R3),苗木干物质积累分别以接种T、G、S7 的增幅最大,达28.4%、23.3%、19.7%。由此可见,相比长期胁迫,短期胁迫后复水的补偿效应更高。

图1 持续干旱及复水后不同苗木干物质的变化Fig.1 Change of biomass to different seedling under the condition of continuous drought and rewatering
由图2可知,在持续干旱前21 d,苗木根冠比无明显变化,随着胁迫时间的增加(28~35 d),苗木根冠比逐渐增加,且菌根化苗木根冠比一直高于CK,但无显著性差异(P>0.05)。这表明干旱胁迫能够促进苗木根系生长,使根干物质积累量增加,根冠比增大。所有处理复水后的根冠比都小于复水前,持续干旱14 d和21 d复水后(R1,R2),接种G 的苗木降幅最大,分别为37.0%和42.0%;持续干旱28 d 复水后(R3),接种S1 的苗木降幅最大,达36.6%。这可能是由于胁迫后复水增加了菌根化马尾松苗木各个器官的干物质积累量,况且叶和茎对水分敏感,其干物质积累速率大于根。此外,由于速生期(6—8月)菌根化苗木生长重心向地上部分转移,增加了地上部分的干物质分配比例,导致根冠比降低。
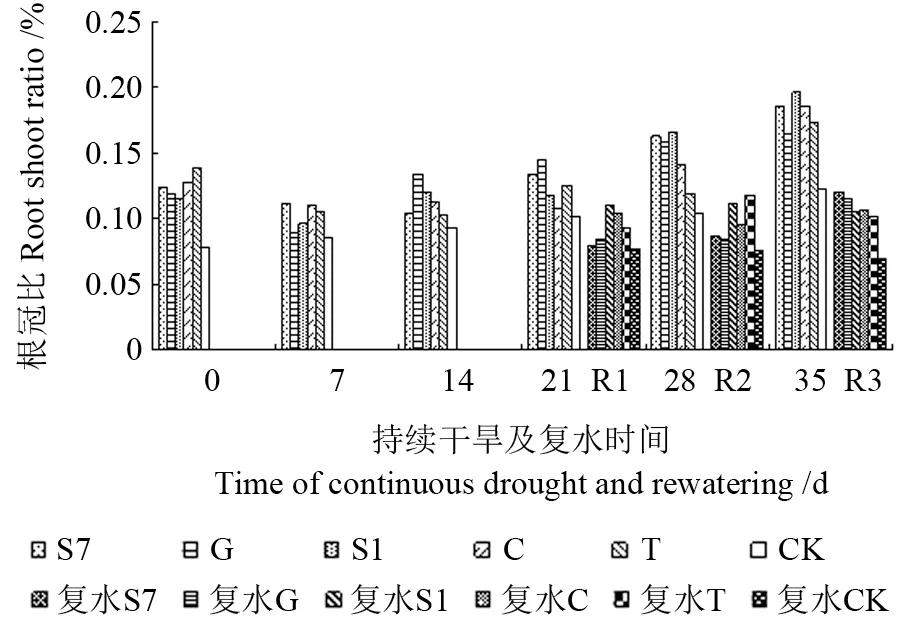
图2 持续干旱及复水后不同苗木根冠比的变化Fig.2 Change of R:S ratio to different seedling under the condition of continuous drought and rewatering
2.2 持续干旱及复水对不同苗木根系形态的影响
由图3A 可知,持续干旱下,外生菌根真菌显著促进了苗木根长生长,整个过程中以接种S1 的苗木根长最长,同CK 比增幅最大达67.4%,最小达48.5%,且菌根化苗木与CK 间有显著性差异(P<0.05)。干旱胁迫诱导根系向下伸长,促使苗木能吸收到更深层土壤的水分,以满足其生长要求。从图3A 可看出,胁迫时间越长其根系越长,复水后根系又进一步生长延伸,根长大于复水前,菌根化苗木根长的生长显著大于CK,其中以接种S1 的苗木在干旱胁迫14 d 复水后(R1),增幅最大达59.6%,这大大增强了菌根化苗木抗倒伏性,增加了后期的耐旱性。
持续干旱使苗木根粗大大降低,但菌根化苗木根粗一直高于CK,其中接种S1 的苗木在持续干旱过程中表现出较好的促生效果,与CK 比增幅最大达44.8%,最小达24.1%。干旱胁迫14 d 和21 d 复水后(R1,R2)苗木根粗较复水前均增加,分别以接种C 和S7 的苗木增幅最大,达18.3%和13.0%;持续干旱28 d 复水后(R3),苗木根粗较复水前降低,但菌根化苗木降幅小于CK,且菌根化苗木根粗最终高于持续干旱35 d。可见,菌根化苗木在干旱及复水过程中能表现出较好的促生效果,短历时(14~21 d)干旱后复水更利于菌根化苗木的恢复(图3B)。
由图3C—D 可知,马尾松苗木侧根数和侧根长随干旱胁迫强度的增加而逐渐增加,最后趋于平稳。整个过程中,菌根化苗木侧根数和侧根长一直高于CK,其中接种S1 的苗木侧根数和侧根长最大,与CK 比增幅最大达122%和204%,最小达94%和74%,菌根化苗木与CK 间差异极显著(P<0.01)。表明在持续干旱下外生菌根真菌能诱导苗木侧根生长并产生更多侧根,扩大根系对水分的吸收范围,从而提高菌根化苗木抗旱性。复水后苗木侧根数和侧根长均有不同程度的恢复,且菌根化苗木恢复度大于CK,其中接种S1 的苗木增幅最大,分别为23.5%、7.1%和5.6%。说明在持续干旱条件下,外生菌根真菌通过促进苗木侧根长和侧根数的生长来适应干旱环境,并在干旱复水后仍能表现出较好的补偿效应。
苗木根体积随干旱胁迫时间的增加而缓慢增加,菌根化苗木根体积一直比CK 高,其中以接种S1 的苗木效果最好,与CK 比增幅最大达450%,最小达97%,且菌根化苗木与CK 间差异极显著(图3E,P<0.01)。与复水前相比,不同干旱胁迫时间复水后苗木根体积增加,且菌根化苗木增幅大于CK,其中接种T 的苗木增幅最大,分别为39.1%、8.7%和15.8%。

图3 持续干旱及复水后不同苗木根系形态变化Fig.3 Change of root to different seedling under the condition of continuous drought and rewatering
在持续干旱条件下,外生菌根真菌能增加苗木根系总吸收面积和活跃吸收面积,从而增强苗木根系活力。由图4可知,随着胁迫时间的增加,苗木根系总吸收面积和活跃吸收面积均表现出降—升—降的趋势,以持续干旱21 d 时苗木根系活力最强,其中接种S1 的苗木根系总吸收面积和活跃吸收面积最大,同CK 相比分别增加了57.0%和48.9%,差异显著(P<0.05)。可见外生菌根真菌适度的干旱胁迫更能激活苗木自身的抗旱调节系统,增强抗旱性。
持续干旱后复水,苗木根系总吸收面积和活跃吸收面积都有所恢复。从图4A 可看出,复水后苗木根系总吸收面积增加,且菌根化苗木增幅大于CK,在持续干旱14、21 和28 d 复水后,分别以接种S7、S1 和G 的苗木增幅最大,达14.7%、21.9%和32.9%。从图4B 可看出,复水后苗木根系活跃吸收面积增加明显,同CK 相比,菌根化苗木差异显著(P<0.05),持续干旱14、21 和28 d 复水后,苗木根系活跃吸收面积较复水前显著增加,其中持续干旱14 d 和21 d 复水后(R1,R2),以接种S7 的苗木增幅最大,达254%和143%;持续干旱28 d 复水后(R3),以接种S1的苗木增幅最大,达48%。结果表明,在持续干旱复水后,外生菌根真菌能增加苗木根系吸收面积,提高苗木根系活力,并表现出一定的补偿效应,其中胁迫14 d 和21 d 复水后菌根化苗木恢复效果最好,说明短期轻度干旱锻炼能有效提高菌根化苗木对干旱胁迫的适应能力。
2.3 持续干旱及复水后不同苗木水分代谢的变化
2.3.1 不同苗木叶片相对含水量RWC 和水分饱和亏缺WSD 的变化
干旱胁迫下,植物叶片含水量的变化可以反映植物叶片持水的能力[17]。植物受到干旱胁迫时,该指标会随水分胁迫强度的增加而减少[18]。随着干旱时间的延长,马尾松苗木叶片的相对含水量降低,但菌根化苗木的相对含水量一直高于CK,且各处理间差异显著(图5A,P<0.05)。这表明外生菌根真菌在持续干旱胁迫下可提高马尾松苗木相对含水量,以此减轻水分胁迫对其造成的影响。
由图5A 可见,在持续干旱14 d 后复水(R1),除接种T外,菌根化苗木相对含水量均超过复水前,以接S1 的苗木增幅最大,达48.6%,各处理间差异显著(P<0.05);在持续干旱21 d 后复水(R2),苗木相对含水量均显著增加,各处理间差异显著(P<0.05);在持续干旱28 d 后复水(R3),与复水前相比菌根化苗木相对含水量略有增加,但差异不显著(P>0.05)。说明在持续干旱不同时间复水后苗木相对含水量有不同程度的提高,表现出一定的补偿效应,而接种外生菌根真菌后显著促进了这一效应。在持续干旱21 d 后复水,苗木相对含水量增幅最大,其中接种G、S1、C和T 的苗木补偿效应最好。
由图5B 所示,随着干旱胁迫时间的增加,苗木叶片水分饱和亏缺逐渐上升,而CK 水分饱和亏缺显著高于菌根化苗木,且在同一胁迫时间内各处理间差异显著(P<0.05)。这表明外生菌根真菌能显著降低苗木水分饱和亏缺,从而提高苗木保水力。在干旱胁迫不同时间复水均能降低苗木水分饱和亏缺(图5B)。干旱胁迫14 d后复水(R1),同复水前相比,苗木水分饱和亏缺下降,且接种G 的苗木降幅最大,为68.5%;干旱胁迫21 d后复水(R2),菌根化苗木水分饱和亏缺明显低于复水前,其降幅大小依次是:S1(74.5%)>C(67.6%)>G(66.0%)>S7(60.5%)>T(59.0%),由此看出,当胁迫时间持续增加时,菌根化马尾松苗木维持胞内水分平衡的自我保护能力也逐渐增强。持续胁迫28 d 后复水(R3),苗木水分饱和亏缺较复水前有所下降,但差异不显著(P>0.05)。说明干旱复水后,菌根化苗木比CK 更能显著降低苗木水分饱和亏缺,提高苗木含水量,增强苗木抗旱性,但随着干旱胁迫的不断增强,苗木恢复能力达到一定程度后减弱,在干旱胁迫第21 天复水后恢复能力最大。
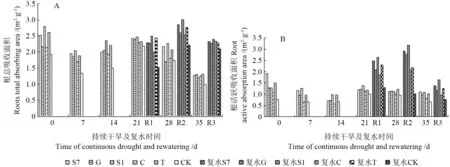
图4 持续干旱及复水后不同苗木根系活力的变化Fig.4 Change of root vigor to different seedling under continuous drought and rewatering
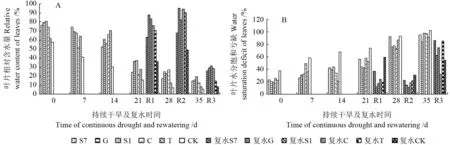
图5 持续干旱及复水对不同苗木相对含水量和水分饱和亏缺的影响Fig.5 Effect of continuous drought and rewatering to different seedling in RWC and WSD
2.3.2 不同苗木叶片自由水/束缚水的变化
由图6可见,随着土壤含水量的降低和胁迫时间的增加,苗木自由水与束缚水的比值先降后增,在持续干旱21 d 时达最小,然后逐渐增加,其中菌根化苗木自由水与束缚水的比值均低于CK,以接种S1 的苗木自由水与束缚水的比值最低,较CK 降低了53.9%,可看出在一定干旱胁迫下,外生菌根真菌通过降低自由水含量,相应增加束缚水的含量,来增强苗木抗旱能力。
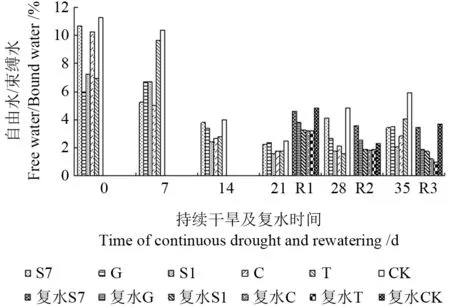
图6 持续干旱及复水后不同苗木自由水/束缚水的变化Fig.6 Change of free water/bound water to different seedling under the condition of continuous drought and rewatering
在干旱胁迫的不同时期复水效果不同,持续干旱14 d 和21 d 后复水(R1,R2),菌根化苗木自由水与束缚水的比值都回升,分别以接种S1和S7 的苗木增幅最大,较CK 增加了36.4%和58.2%;而持续干旱28 d 后复水(R3),苗木自由水与束缚水的比值降低。说明复水后外生菌根真菌能增加苗木自由水含量,以促进各种正常生理代谢,而长历时(28 d)胁迫的恢复能力不如短历时胁迫(14~21 d)。
2.3.3 不同苗木水势的变化
如图7所示,随着干旱胁迫时间的增加,苗木叶水势总体呈上升趋势,而菌根化苗木叶水势一直显著低于CK(P<0.05),其中以接种G 的苗木降幅最大,达63.9%。当苗木受到干旱胁迫时,其水势呈上升趋势,复水后有所恢复。从图7可以看出,持续干旱14 d 后复水(R1),接种S1、C 和T 的苗木叶水势较复水前(14 d)明显下降,降幅分别为30%、31%和39%,菌根化苗木与CK间差异显著(P<0.05);持续干旱21 d后复水(R2),菌根化苗木叶水势均降低,降幅大小依次是:
T(49.3%)>S1(43.7%)>S7(36.0%)>G(12.2%)>C(4.4%),同CK 相比,菌根化苗木有显著性差异(P<0.05);持续干旱28 d后复水(R3),苗木叶水势除接种G 和C 外,其他都高于复水前,且同CK 相比,菌根化马尾松苗木差异显著(P<0.05)。可见菌根化苗木在不同胁迫时间复水后表现有差异,适度胁迫有利于植物水分的补偿作用。
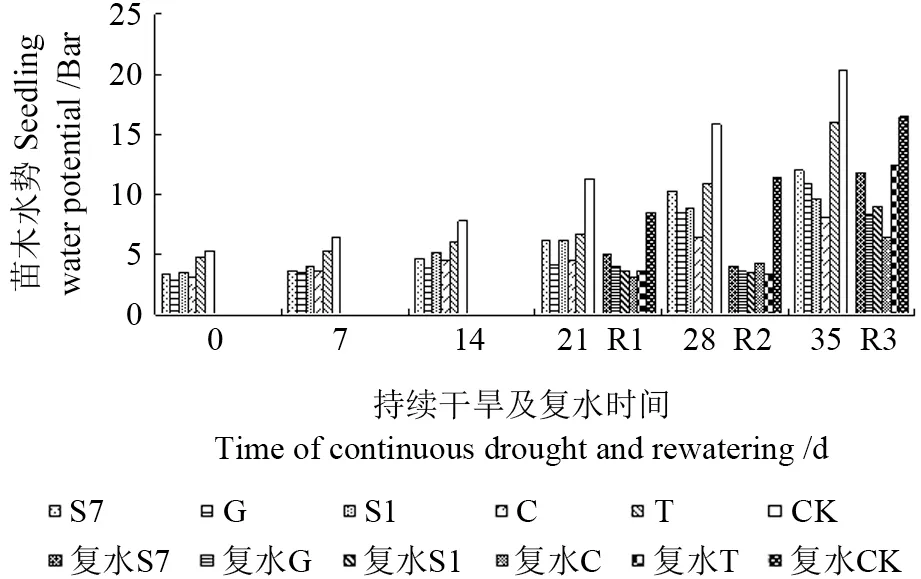
图7 持续干旱及复水后不同苗木水势的变化Fig.7 Change of water potential to different seedling under the condition of continuous drought and rewatering
2.4 持续干旱及复水后不同苗木渗透调节物质的变化
2.4.1 不同苗木可溶性糖的变化
如图8所示,在外生菌根真菌作用下,苗木可溶性糖含量增加,同一胁迫时间下菌根化苗木可溶性糖含量比CK 都高。不同水分胁迫时间对菌根化苗木可溶性糖含量的影响不同,当持续干旱时间逐渐增加时,苗木可溶性糖含量先增后降,在干旱胁迫第14 天达最大,其中接种S1 的含量最高,比CK 提高了33.1%,菌根化苗木与CK 间无显著性差异(P>0.05)。
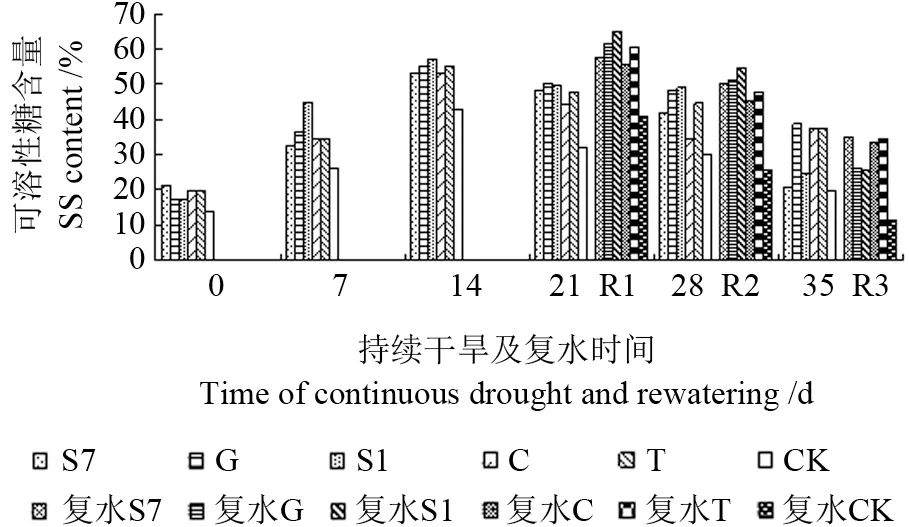
图8 持续干旱及复水后不同苗木可溶性糖的变化Fig.8 Change of soluble sugar to different seedling under the condition of continuous drought and rewatering
复水对苗木可溶性糖有不同影响,恢复程度与胁迫时间有关(图8)。在持续干旱14 d 和21 d后复水(R1,R2),发现菌根化苗木可溶糖含量较复水前增加,而CK 可溶糖含量则降低,各菌根化苗木均以干旱14 d 后复水效果最好,且接种S1的苗木增幅最大,达13.5%,菌根化苗木同CK 相比差异显著(P<0.05)。在持续干旱28 d 后复水(R3),苗木可溶性糖含量下降,而CK 降幅最大,达61.1%,同CK 相比菌根化苗木差异显著(P<0.05)。可见,胁迫程度和持续时间影响着苗木复水后的补偿效应,在轻度(14 d)和中度(21 d)干旱胁迫后,外生菌根真菌能增强苗木在复水后的补偿效应,并以轻度胁迫后的补偿效应最大。当苗木受到严重胁迫(28 d)后复水,自我恢复能力则会减弱,但接种菌根菌可缓解这一情况。
2.4.2 不同苗木蔗糖的变化
由图9可见,在整个水分持续干旱期间,菌根化苗木蔗糖含量均高于CK,并随持续时间的延长,变化趋势呈先增后降的单峰曲线,且峰值均出现在胁迫第14 天,其中接种C 的苗木蔗糖含量最高,较CK 提高了15.5%。持续干旱14 d 后复水(R1),菌根化苗木蔗糖含量与复水前(14 d)相比有所增加,增幅大小为G(109.5%)>S7(28.0%)>S1(7.3%)>C(3.4%)>T(3.3%);在持续干旱21 d 和28 d 后复水(R2,R3),苗木蔗糖含量与复水前相比(21 d,28 d)有所降低,甚至与极端干旱(35 d)时相近。表明在干旱复水后,外生菌根真菌能提高苗木蔗糖含量,而复水后的补偿生长效应表现为胁迫影响程度越大补偿效应越小。
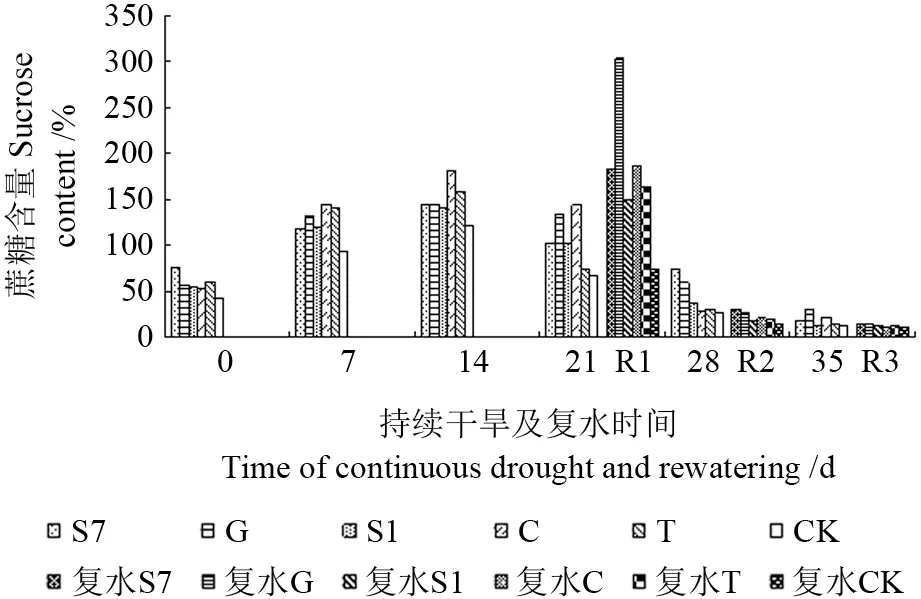
图9 持续干旱及复水后不同苗木蔗糖含量变化Fig.9 Change of sucrose sugar to different seedling under the condition of continuous drought and rewatering
2.4.3 不同苗木游离脯氨酸的变化
由图10可见,脯氨酸含量随胁迫时间的增加呈渐增趋势,且菌根化马尾松苗木脯氨酸含量低于CK,其中接种S1 的苗木脯氨酸含量降幅最大,达66.5%。说明当苗木遭受干旱胁迫时,其自身会产生大量游离脯氨酸进行渗透调节,而外生菌根真菌改善了植株水分代谢,避免过度失水,减轻苗木受胁迫的程度,因此无需菌根化苗木合成更多的脯氨酸进行渗透保护作用。
干旱胁迫不同时间复水后各处理苗木游离脯氨酸含量变化见图10。在干旱胁迫14 d 后复水(R1),菌根化苗木脯氨酸含量较胁迫结束时(14 d)下降,降幅大小为T(61.9%)>C(44.4%)>S7(42.1%)>S1(16.1%)>G(0.06%),而CK 较胁迫结束时(14 d)增加,菌根化苗木同CK 相比有显著性差异(P<0.05);在持续干旱21 d 后复水(R2),菌根化苗木脯氨酸含量与复水前相比显著降低,降幅大小为S1(80.7%)>C(76.7%)>T(75.0%)>G(56.3%)>S7(48.4%);在干旱胁迫28 d 后复水,苗木脯氨酸含量与复水前基本持平。由此可见,不同持续干旱时间复水后,苗木脯氨酸含量的变化也有差异,短期(14~21 d)干旱胁迫后复水的补偿效应明显高于长期(28 d)干旱胁迫。
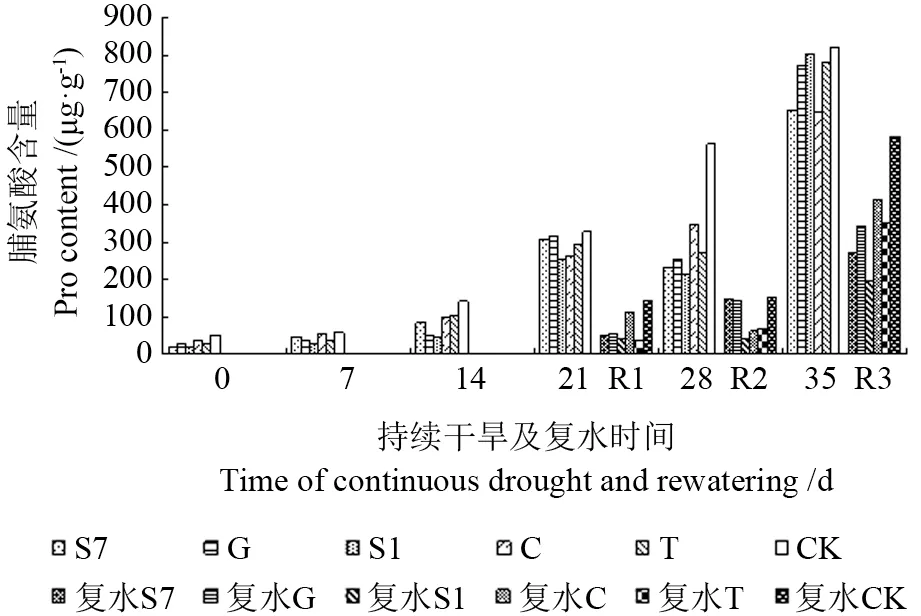
图10 持续干旱及复水后不同苗木游离脯氨酸含量的变化Fig.10 Change of prolineto different seedling under the condition of continuous drought and rewatering
2.4.4 不同苗木可溶性蛋白的变化
图11可见,随干旱胁迫时间的延长,苗木可溶性蛋白含量逐渐下降,但菌根化苗木可溶性蛋白含量一直高于CK,其中接种S1 的苗木增幅最大,达109%,菌根化苗木同CK 相比有显著性差异(P<0.05)。可见在干旱胁迫下,外生菌根真菌能显著提高苗木可溶性蛋白含量,从而提高苗木渗透调节能力,减轻水分逆境造成的代谢受阻,增强苗木抗旱性。
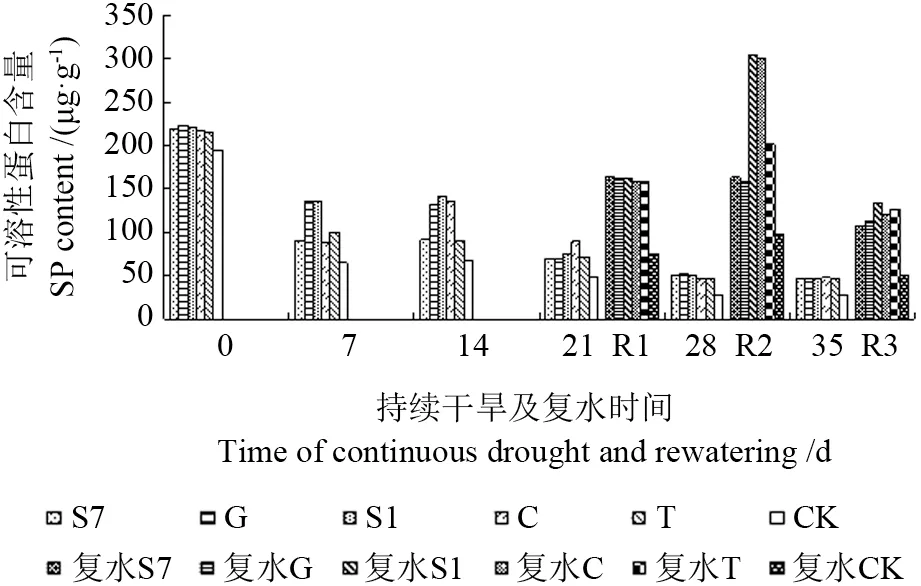
图11 持续干旱及复水后不同苗木可溶性蛋白的变化Fig.11 Change of solubility protein to different seedling under the condition of continuous drought and rewatering
干旱胁迫解除后苗木可溶性蛋白含量都有一定程度的恢复(图11),且菌根化苗木恢复能力大于CK,并与CK 间差异显著(P<0.05)。在持续干旱14 d 后复水(R1),接种S7 的苗木可溶性蛋白含量最高,增幅最大达79.3%;而在持续干旱21 d 后复水(R2),接种S1 和C 的苗木可溶性蛋白含量显著增加,表现出超补偿效应;在持续干旱28 d 后复水,菌根化苗木可溶性蛋白含量仍显著高于复水前(28 d),仍表现出一定的补偿效应,由此可见,马尾松苗木渗透调节物质受干旱胁迫程度影响较大,菌根化苗木可溶性蛋白含量在复水后能维持在较高水平,以调节正常的细胞代谢活动。而不同菌种复水后在不同胁迫时间的表现有差异,持续干旱21 d 后复水(R2),接种S1 和C 的苗木效果最好。
2.5 持续干旱及复水后不同苗木抗旱指标的聚类分析
选取菌种、可溶性糖含量、蔗糖含量、可溶性蛋白含量、脯氨酸含量、根长、根粗、>5 cm 侧根数、>5 cm 侧根长、根体积、根总吸收面积、根活跃吸收面积、叶片相对含水量、自然水分饱和亏、自由水/束缚水、水势、根冠比、生物量18 个指标,对菌根化马尾松苗木的抗旱性进行综合评价。
如图12所示,将18 个指标分为两类,聚类质量基本良好。最大类含样品37 例,占比68.5%;最小类含样品17 例,占比31.5%,两者比为2.18。由表3可看出抗旱指标的分布特征以及这些指标在聚类分析中的重要性,18 个指标的重要程度从高到低依次为:侧根数>侧根长>根体积>根长>自由水与束缚水比>可溶性糖含量>生物量>菌种>根冠比>根活跃吸收面积>根粗>根总吸收面积>脯氨酸含量>水势>蔗糖含量>自然水分饱和亏缺>可溶性蛋白含量>叶片相对含水量。从图12中可知,5 个菌种中S1 在马尾松菌根化苗抗旱中作用最大,效果最好,接种S1 的苗木抗旱能力最强。从表3可看出侧根数、侧根长、根体积以及根长的重要程度较高,它们都属于根系形态,说明根系的生长形态是决定马尾松菌根化苗抗旱性的关键。

图12 持续干旱和复水后影响苗木抗旱性18 个指标的模型概要、聚类质量及聚类大小Fig.12 Model summary,cluster quality and cluster size of 18 indexes affecting drought resistance of seedlings under continuous drought and rewatering
3 结论与讨论
本试验研究结果表明,在干旱胁迫下,接种外生菌根真菌大大促进了苗木根系及地上部分生长,这与王秀丽等[6]及以往[19]的研究结果一致。接种外生菌根真菌后,苗木的苗高、地径、生物量及根冠比被明显提高,其中接种S1 和T 对幼苗在生长上促进效果较好,且菌根依赖性最强,其主要原因在于菌根菌能诱导苗木侧根生长并产生更多侧根,扩大根系对水分的吸收范围,提高苗木根系吸收面积,从而提高菌根化苗木抗旱性,这与简在友等[20]的结果一致。而且在适度的干旱胁迫下,外生菌根真菌更能激活苗木自身的抗旱调节系统,增强抗旱性。
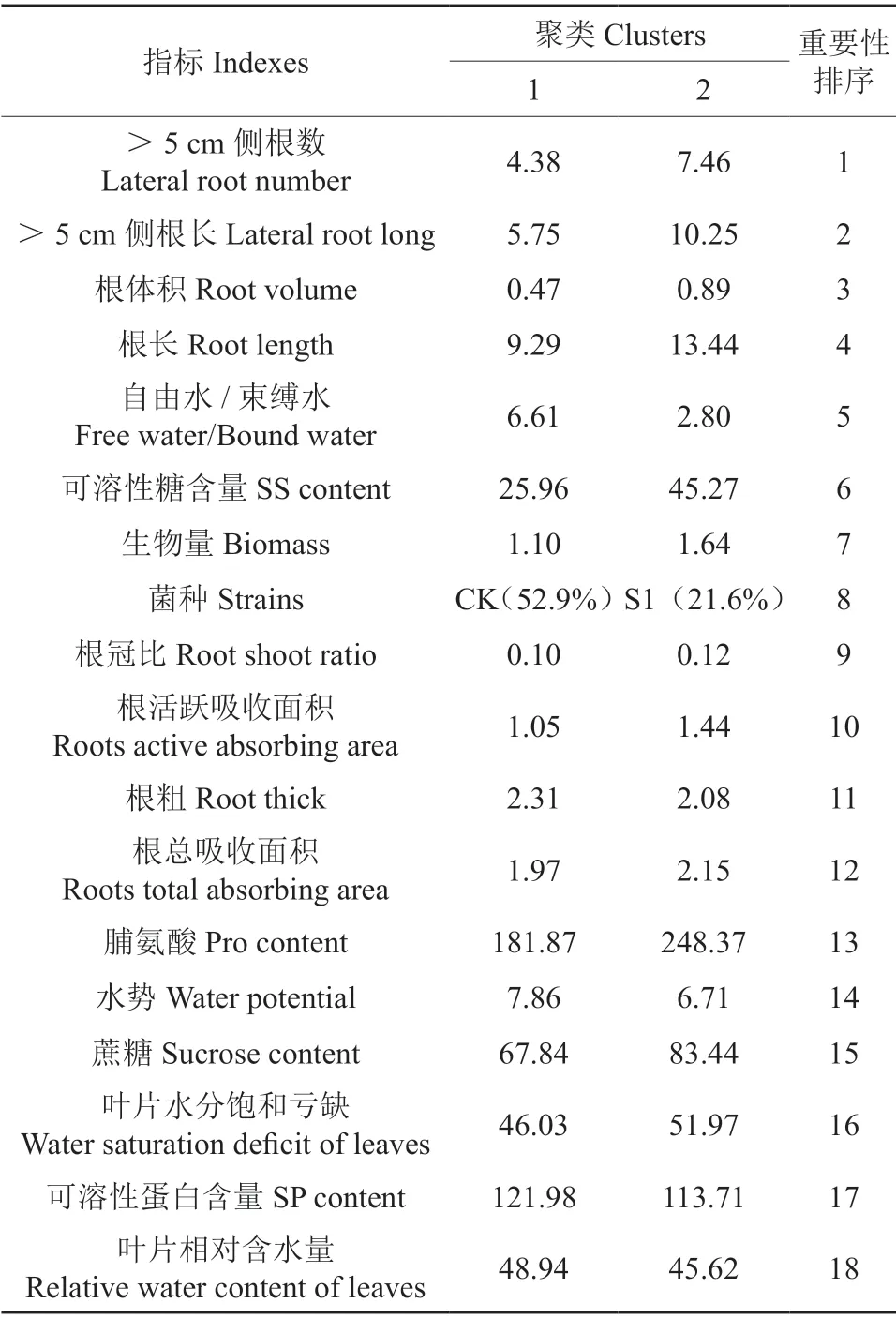
表3 聚类及输入(预测变量)重要性†Table 3 Clusters and input (predictor) important
植物的水分状况直接影响其抗旱性,这是因为植物的水分状况关系到细胞膨压,酶的活性以及有机物的分解、合成、转化、运输和新器官的形成[21]。有研究表明叶片水分饱和亏缺是叶片持水能力的标志,也是植物体内受旱程度的反映[22]。本试验结果表明,干旱胁迫后,菌根真菌显著降低了苗木水分饱和亏缺,苗木叶水势总体呈上升趋势,而菌根化苗木叶水势一直显著低于未接种苗木,这与前人的研究结果一致[21],说明接种菌根菌提高了苗木含水量,增强了苗木抗旱性,从而减轻水分胁迫造成的伤害,促进植物生长发育。本试验还显示菌根化苗木在不同胁迫时间复水后的表现不同,说明适度胁迫有利于植物水分的补偿效应。
渗透调节物质的积累可以稳定植物体内的渗透压平衡,提高植物保水能力[23]。在试验中,干旱胁迫下外生菌根真菌提高了苗木可溶性糖含量和蔗糖含量。叶片中可溶性糖含量的积累可使细胞水势降低,细胞内外渗透势差增加,使外界水分有利于向细胞内扩增,从而维持植株生长[24-25]。脯氨酸是苗木主要渗透调节物质,本试验研究显示,脯氨酸含量随胁迫时间的延长呈渐增趋势,但菌根化苗木脯氨酸含量低于未接种苗木,这证实了张中峰等[26]的研究结果,说明外生菌根真菌改善了植株水分代谢,避免过度失水,减轻苗木受胁迫的程度,因此无需菌根化苗木合成更多的脯氨酸进行渗透保护作用。接种外生菌根真菌也显著增加了可溶性蛋白含量,从而提高苗木渗透调节能力,减轻水分逆境造成的代谢受阻,增强苗木抗旱性。
此外,通过对马尾松不同苗木抗旱指标的二阶聚类分析表明,接种S1 的苗木抗旱能力最强,侧根数、侧根长、根体积以及根长的重要程度最高,可见这几个指标对苗木抗旱具有更为关键的作用。因为根系对土壤水分反应敏感,干旱胁迫诱导根系向下伸长,使其能吸收更深层土壤的水分,以满足苗木生长需求。侧根数量越多、侧根长度越长,就越利于扩大根系对水分的吸收范围,从而提高菌根化马尾松苗木抗旱性。
综上所述,持续干旱及复水后外生菌根能够促进马尾松苗木生长,促进其根系形态形成,改善水分利用状况,提高渗透调节物质含量,其中接种S1 的苗木抗旱能力最强,而根系形态是影响马尾松不同苗木抗旱性的关键指标。目前,本试验仅从干旱复水对马尾松菌根化苗木生理特性进行了研究,在细胞水平上菌根化苗木是如何响应干旱胁迫、其分子机制是怎样以及抗旱基因有哪些等问题,均有待于做进一步深入研究,以便为马尾松菌根化苗木定向培育及育种提供理论依据和基因资源,为菌根技术的开发和利用提供技术支撑。
猜你喜欢
农业灾害研究(2022年6期)2022-12-02
辽宁林业科技(2022年2期)2022-11-24
中国食用菌(2022年5期)2022-06-08
农业工程学报(2022年1期)2022-03-25
河南科技(2022年1期)2022-03-11
新农业(2021年12期)2021-11-29
南方农业·中旬(2021年5期)2021-10-25
绿色科技(2020年1期)2020-11-29
知识就是力量(2017年12期)2018-01-15
农家顾问(2016年7期)2016-05-14
