
转Cry1Ah基因抗虫玉米HGK60对生物多样性的影响
2021-04-25 09:52陈彦君李俊生
环境科学研究 2021年4期
陈彦君, 李俊生, 闫 冰, 关 潇*, 陶 均
1.中国环境科学研究院, 国家环境保护区域生态过程与功能评估重点实验室, 北京 100012 2.海南大学热带作物学院, 海南 海口 570228 3.江西省科学院能源研究所, 江西 南昌 330096
转基因作物商业化25年以来,总种植面积累计达到25×108hm2[1]. 在作物生长过程中,虫害的发生以及除草剂的施用严重危害着其产品品质和产量. 通过转基因技术导入抗虫、抗除草剂基因可有效降低靶标害虫的数量,降低作物对化学杀虫剂的依赖,缓解除草剂对作物的影响[2],但其是否会对生态环境造成影响仍需要进行全方位的安全性测定,这也是转基因作物环境安全评价的重要内容.
对生物多样性的影响研究作为转基因作物环境安全释放的必要环节之一,目前已开展大量相关研究,但结论不尽相同. 部分研究表明,转基因作物种植对根际土壤微生物[3-6]、昆虫[7-10]、杂草[10-12]等的群落结构与组成无显著影响,但也有学者认为其种植会影响田间生物多样性[13-16],研究结果的差异表明转基因作物种植后可能对生物多样性产生的影响并非一概而论,受体及外源基因等的差异都可能影响研究结果. 因此,对于转基因作物的安全性评价工作应遵循“个案原则”[17].
Cry1Ah基因的主要靶标动物为鳞翅目昆虫,对其具有较高的杀虫活性,但对家蚕等重要经济昆虫的杀虫活性相对较低,表现出良好的环境安全性[18],具有广阔的应用前景,但其是否会对生物多样性产生显著影响仍处于探索阶段. 鉴于此,该文以转Cry1Ah基因抗虫玉米HGK60为研究对象,重点讨论其对根际土壤微生物、田间节肢动物以及田间杂草的影响,以期为转Cry1Ah基因抗虫玉米HGK60的环境安全评价提供科学依据,为生物安全管理的健康发展提供支持.
1 材料与方法
1.1 研究区域概况
该研究于中国农业科学研究院海南乐东南繁生物育种试验基地进行. 该基地位于海南岛西南部乐东黎族自治县,属热带季风性气候,年均气温为23.9~25.2 ℃,光照充足,热量丰富,年降水量约 1 000 mm,但水量不平衡,干季、雨季分明. 选用玉米品种为转Cry1Ah基因抗虫玉米HGK60(简称“HGK60”)及其对照常规玉米郑58(简称“郑58”),均由中国农业科学院生物技术研究所提供.
试验采取随机区组设计,试验组为HGK60,对照组为郑58,每个处理均设置4个重复,每个重复(小区)面积为150 m2(10 m×15 m). 各小区间设置1 m宽的空白隔离带,种植方式为一穴一粒、一穴两粒循环播种,行距为60 cm,株距为25 cm. 玉米于2018年11月播种,田间管理同常规种植.
1.2 样品采集与处理
1.2.1根际土壤样品的采集、DNA提取、PCR扩增、高通量测序
样品采集. 在玉米生长的苗期、喇叭口期、抽穗期、完熟期以及玉米收获后,采用“抖根法”按照五点采样法采集根际土壤样品,取样时避开道路等可能影响因素. 土壤样品用干冰保存带回置于-80 ℃ 下.
DNA提取. 选用FastDNA® SPIN Kit for soil(MP Biomedicals, Santa Ana, CA, US)试剂盒提取根际土壤微生物总DNA,采用超微量分光光度计检测其浓度、纯度.
PCR扩增. 针对V1~V9区,以稀释后的基因组DNA为模板,使用通用引物8F(5′-AGAGTTTGATCC TGGCTCAG-3′)和1509R(5′-GNTACCTTGTTACGAC TT-3′)进行PCR扩增. PCR反应体系(50 μL):Trans Fastpfu 1 μL,5×Buffer 10 μL,5×StimuLate 5 μL,dNTPs(2.5×10-3mol/L each) 5 μL,Primer Mix(1 μmol/L) 2 μL,gDNA 1 μL,NFW 26 μL. PCR反应条件:预变性温度98 ℃(2 min);变性温度95 ℃(30 s),退火温度60 ℃(45 s),延伸温度72 ℃(90 s),35个循环后72 ℃终止延伸10 min. 根据PCR产物浓度进行等质量混样,充分混匀后使用1×TAE 浓度为2%的琼脂糖凝胶电泳纯化PCR产物,割胶回收目标条带. 所用试剂盒为QIAGEN公司QIAquick@ Gel Extraction Kit凝胶回收试剂盒.
高通量测序. 使用PACIFIC BIOSCIENCES(PacBio)公司的SMRTbellTMTemplate Prep Kit建库试剂盒进行文库的构建,通过Qubit定量和FEMTO Pulse检测片段大小,库检合格后使用PacBio的Sequel平台进行16S全长rDNA测序. 该研究测序工作委托北京诺禾致源生物信息科技有限公司完成.
1.2.2节肢动物调查
在玉米生长的苗期、喇叭口期、抽穗期和完熟期,采用五点法于每个生育期分别进行3次节肢动物调查:①直接观察法,各点位调查15株玉米,记录植株上节肢动物种类和数量;②陷阱调查法,各小区埋设25个(每个点位放5个)塑料杯,塑料杯直径7.5 cm,间隔0.5 m,于调查的前一天在塑料杯中放置1/3容量的5%防冻液,24 h后记录节肢动物的种类和数量.
1.2.3杂草调查
分别在玉米生长的苗期、喇叭口期、抽穗期、完熟期于各小区按照五点法,避开调查节肢动物时所设陷阱,选取5个1 m2(1 m×1 m)的样方,调查并记录样方内杂草种类和数量.
1.3 数据统计与分析
1.3.1根际土壤微生物
1.3.1.1测序数据处理
将PacBio下机数据导出后,使用Lima软件根据Barcode序列区分各样本数据. 进行CCS(Circular Consensus Sequencing)校正及长度过滤,过滤含有连续相同碱基数>8的序列及引物切除. 对处理后得到的序列进行去除嵌合体处理,通过UCHIME v7.0.1090与Silva全长数据库进行比对检测嵌合体序列,去除嵌合体序列,得到最终有效数据.
1.3.1.2OTU聚类和物种注释
利用Uparse v7.0.1001软件对所有样品的全部有效数据进行聚类,以97%的一致性将序列聚类成可操作分类单元OTUs(Operational Taxonomic Units). 依据相应算法原则,筛选OTUs中出现频数最高的序列作为其代表序列. 对OTUs代表序列进行物种注释,用Mothur方法与Silva(http://www.arb-silva.de)的SSUrRNA数据库进行物种注释分析(设定阈值为0.8~1),获得分类学信息并统计各分类水平上各样本的群落组成. 使用MUSCLE v3.8.31软件进行快速多序列比对,得到所有OTUs代表序列的系统发生关系. 最后以数据量最少的样品为标准,对各样品数据进行均一化处理,后续分析均基于均一化处理后的数据进行. 使用R v2.15.3软件绘制门水平上的前10位物种相对丰度柱形图与属水平上的前35位物种相对丰度聚类热图,并对同一生育期的组间差异进行分析(T检验).
1.3.1.3样品复杂度分析(alpha diversity)
使用Qiime v1.9.1软件计算Shannon-Wiener指数、Simpson指数、Chao1指数、ACE指数、Phylogenetic diversity指数(简称“PD指数”),使用R v2.15.3软件进行alpha多样性指数组间差异分析(非参数Wilcoxon检验).
1.3.1.4多样品比较分析(beta diversity)
用Qiime v1.9.1软件计算Weighted-Unifrac距离. 使用R v2.15.3软件进行(NMDS),并进行beta多样性指数组间相似性分析(Analysis of Similarities, ANOSIM).
1.3.2节肢动物与杂草的统计与分析
统计分析玉米各生育期田间节肢动物和杂草的群落结构与组成,计算各生态学指数.
Margalef指数(Dmg):
Dmg=(S-1)/lnN
(1)
Shannon-Wiener指数(H):
(2)
Simpson指数(D):
(3)
Pielou指数(J):
J=H/lnS
(4)
式中:S为物种个数;Pi=Ni/N,其中Ni为第i个物种的个体数,N为总个体数[19].
利用Excel 2010软件进行数据整理,利用SPSS 16.0软件进行统计分析,进行两两T检验.
2 结果与讨论
2.1 转Cry1Ah基因抗虫玉米HGK60对根际土壤微生物的影响
将根际土壤样品的高通量测序序列经过质控过滤后,共得到 633 812 条有效序列. 各样品的高通量测序序列范围为 6 480~55 762 条,平均有效序列为 21 127 条. 根据样品中最小有效序列数,每个样品随机挑选 6 480 条有效序列进行样品测序深度的均一化处理. 以97%的一致性(Identity)将其聚类成OTUs,共注释到34门、188科、524属.
2.1.1物种相对丰度统计
统计根际土壤微生物门水平上的物种相对丰度,丰度前10位的门分别为厚壁菌门(Firmicutes)、变形菌门(Proteobacteria)、酸杆菌门(Acidobacteria)、放线菌门(Actinobacteria)、浮霉菌门(Planctomycetes)、拟杆菌(Bacteroidetes)、绿弯菌门(Chloroflexi)、芽单胞菌门(Gemmatimonadetes)、未鉴定出的细菌(unidentified-bacteria)、蓝细菌门(Cyanobacteria). 其中,厚壁菌门在各处理中的相对丰度为18.88%~45.61%、变形菌门为24.52%~37.68%、酸杆菌门为9.28%~13.16%,且在各处理中相对丰度均大于5%,为优势种群.
结合T检验,比较HGK60与郑58根际土壤微生物发现,酸杆菌门、浮霉菌门、绿弯菌门在各生育期的相对丰度均差异不显著(P>0.05);厚壁菌门仅在抽穗期和完熟期表现为HGK60显著高于郑58(P<0.05),变形菌门仅在抽穗期和完熟期表现为HGK60显著低于郑58(P<0.05),放线菌门仅在完熟期表现为二者差异显著(P<0.05),拟杆菌门、芽单胞菌门仅在抽穗期表现为二者差异显著(P<0.05),但收获后HGK60与郑58在前10个门的相对丰度差异均不显著(P>0.05)(见图1).
根据所有样品属水平上的物种注释,选取相对丰度前35位的属绘制聚类热图(见图2). 比较同一生育期HGK60与郑58根际土壤微生物发现,部分物种存在一定差异,但仅出现在某个生育期,并未在整个生育期持续出现. 苗期,Chitinophaga、金黄杆菌属(Chryseobacterium)等的相对丰度均表现为HGK60低于郑58,寡养单胞菌属(Stenotrophomonas)、Pelomonas等的相对丰度则表现为HGK60高于郑58;喇叭口期,嗜糖假单胞菌属(Pelomonas)、Occallatibacter等的相对丰度表现为HGK60高于郑58;抽穗期,Haliangium、Crenobacter等的相对丰度表现为HGK60低于郑58;完熟期,Aquicella、Dechlorobacter等的相对丰度均表现为HGK60低于郑58. 这表明HGK60的种植会在某些生育期对部分属水平上的物种产生一定影响.
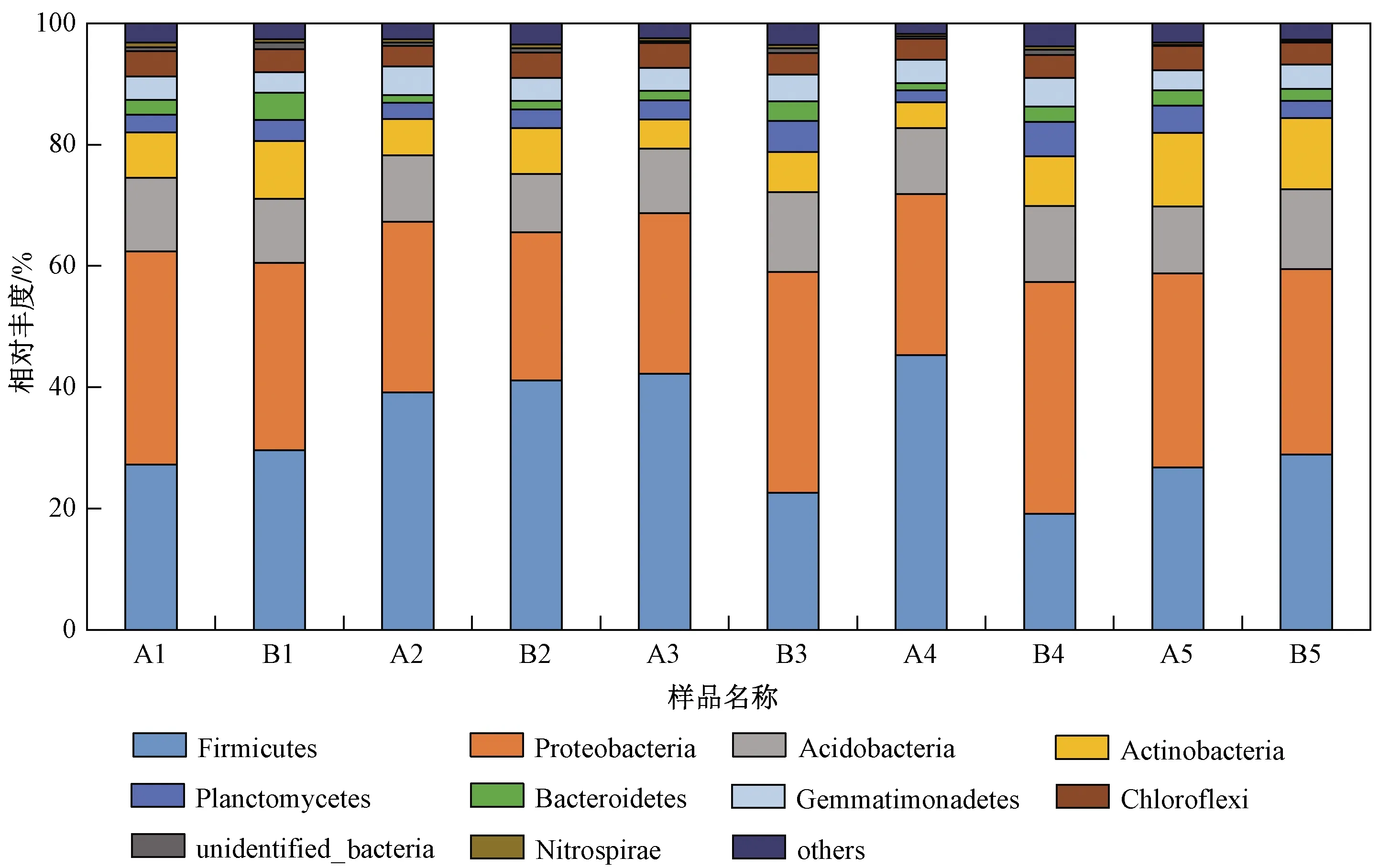
图1 门水平上的前10的物种相对丰度Fig.1 The relative abundance of top 10 soil microorganisms at the phyla level
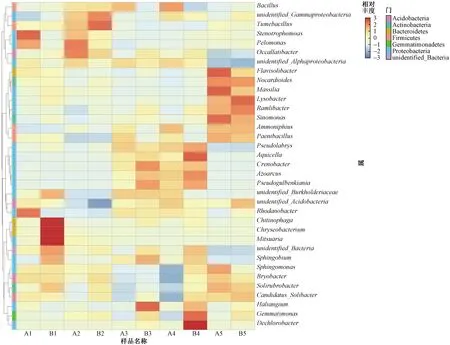
图2 不同植株根际土壤中微生物前35个属相对丰度聚类热图Fig.2 The relative abundance histogram of top 35 soil microorganisms at the genus level by heatmap
从生育期的角度来看,玉米根际土壤微生物部分属水平上的物种呈现一定的季节性差异. 溶杆菌属(Lysobacter)、类诺卡氏属(Nocardioides)等的相对丰度主要表现为收获后相对较高,玉米生长期间则相对较低;Tumebacillus等的相对丰度表现为喇叭口期明显高于其他生育期;Pseudogulbenkiania、Crenobacter、Pseudolabrys、Aquicella等的相对丰度主要表现为抽穗期、完熟期相对较高,其他生育期则相对较低. 部分物种呈现一定的季节性特征,说明生育期是影响根际土壤微生物的因素之一. 但是由于聚类热图只能通过图中不同处理间颜色的差异对处理间差异进行判断,无法进行有效量化,需要结合多样性统计结果做进一步分析.
2.1.2alpha多样性指数与beta多样性聚类分析
对HGK60与郑58各生育期根际土壤细菌群落的alpha多样性进行非参数Wilcoxon检验. Shannon-Wiener指数分析结果显示,HGK60与郑58仅在抽穗期、完熟期差异显著(P<0.05);Simpson指数、Chao1指数在各生育期均表现为二者差异不显著(P>0.05);ACE指数仅表现为收获后二者差异显著(P<0.05);PD指数则表现为仅抽穗期、完熟期二者差异显著(P<0.05). 从整体上看,HGK60的种植不会对土壤微生物alpha多样性产生持续性影响,部分影响主要存在于抽穗期与完熟期,除ACE指数外,收获后HGK60与郑58在各生态指标上差异均不显著(P>0.05)(见表1).
在beta多样性分析中,根据不同生育期玉米根际土壤样品的测序结果,基于Weighted-UniFrac进行NMDS分析,以此对HGK60与郑58根际土壤微生物群落结构的相似性及差异性进行定性分析(见图3). 结合ANOSIM分析,比较同一生育期HGK60与郑58,发现抽穗期、完熟期二者间差异显著(P<0.05),苗期、喇叭口期、收获后二者均差异不显著(P>0.05),表明HGK60种植产生的土壤微生物群落结构的影响仅出现在某些生育期,并未在整个生长阶段持续存在,且收获后差异不显著(P>0.05). 从生育期的角度来看,不同生育期玉米根际土壤样品细菌群落存在明显差异,喇叭口期与抽穗期、完熟期与收获后均表现为差异显著(P<0.05).

表1 玉米根际土壤微生物的alpha多样性指数Table 1 Alpha diversity indexes of rhizosphere soil microorganism

图3 非度量多维尺度(NMDS)分析Fig.3 Non-metric multidimensional scaling
2.2 转Cry1Ah基因抗虫玉米HGK60对田间节肢动物的影响
在HGK60与郑58田间共调查到节肢动物 19 835 头,二者分别有 9 727 和 10 108 头,隶属于7目19科60种(见表2). 比较同一生育期HGK60与郑58玉米田间节肢动物数量分析,螟蛾科在苗期、喇叭口期、抽穗期均表现为HGK60显著低于郑58(P<0.05),其他物种的差异只表现在某个生育期,并不会在整个生育期持续出现(见表3),夜蛾科仅在抽穗期表现为HGK60显著低于郑58(P<0.05),缘蝽科、盲蝽科、叶蝉科仅在喇叭口期表现为HGK60显著高于郑58(P<0.05),飞虱可仅在完熟期HGK60显著低于郑58(P<0.05),蚜科仅在苗期、完熟期表现为二者差异显著(P<0.05),草蛉科仅在抽穗期表现为HGK60显著低于郑58(P<0.05),瓢虫科仅在苗期表现为HGK60显著高于郑58(P<0.05),蓟马总科则仅在喇叭口期HGK60显著低于郑58(P<0.05).

表2 田间节肢动物群落的目、科、种分布Table 2 Patterns of the arthropod community in the fields in different orders, families and species
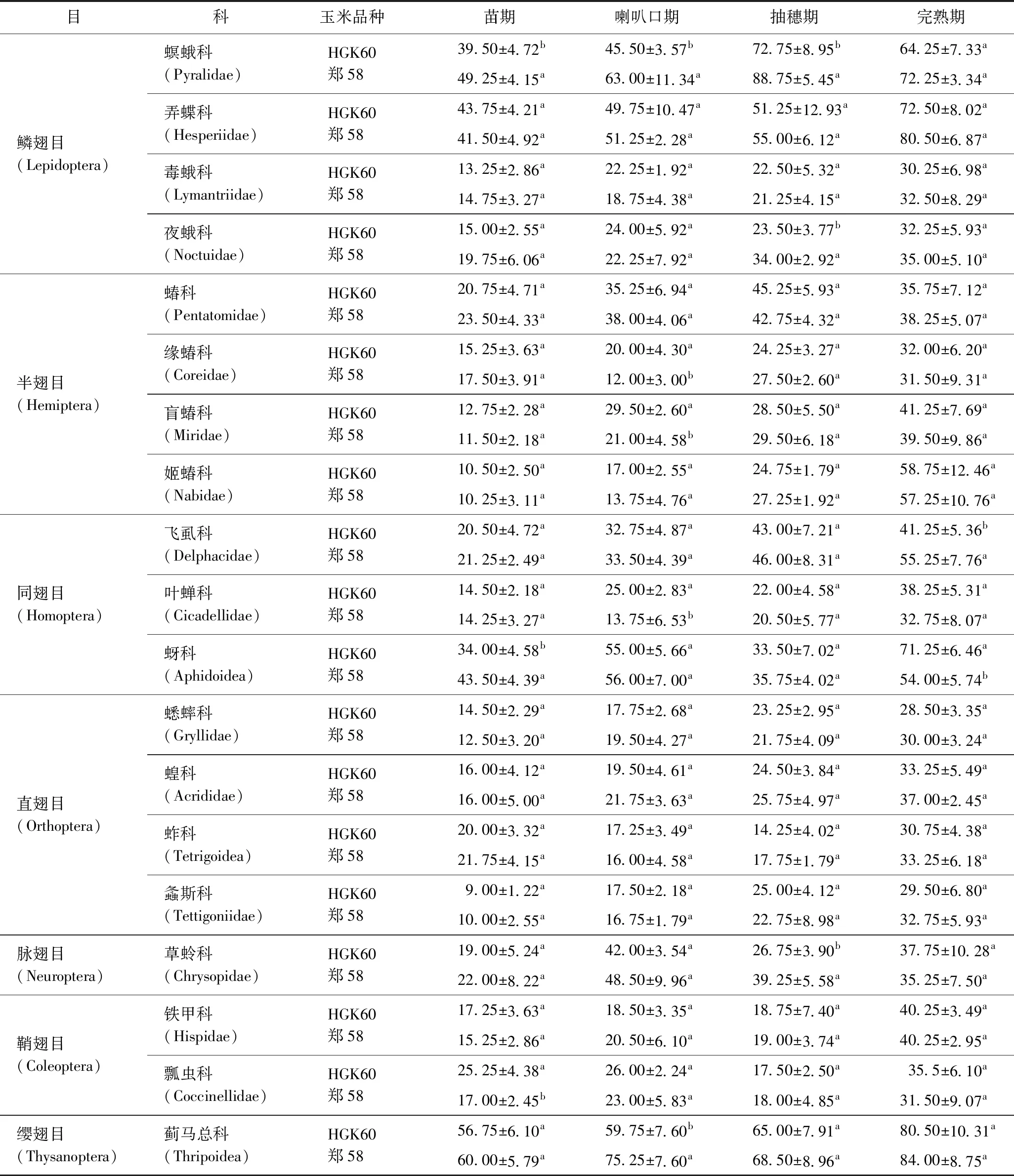
表3 各生育期田间节肢动物数量Table 3 The number of arthropods in each growth period 头
比较HGK60与郑58田间节肢动物群落的Margalef指数发现,仅抽穗期表现为HGK60显著高于郑58(P<0.05),其他生育期均表现为二者差异不显著(P>0.05)〔见图4(A)〕. Shannon-Wiener指数分析结果显示,HGK60与郑58在各生育期差异均不显著(P>0.05)〔见图4(B)〕. Simpson指数表现为仅喇叭口期HGK60与郑58差异显著(P<0.05),其他生育期二者差异均不显著(P>0.05)〔见图4(C)〕. Pielou指数分析结果显示,HGK60与郑58田间节肢动物Pielou指数的变化范围在0.9~1.0之间,且二者在各生育期差异均不显著(P>0.05)〔见图4(D)〕.

图4 田间节肢动物群落各生态指标Fig.4 The ecological indexes of arthropods in the fields
2.3 转Cry1Ah基因抗虫玉米HGK60对田间杂草的影响
调查结果显示,HGK60与郑58田间杂草发生种类有8科16种,多为一年生杂草(见表4). 各生育期内HGK60与郑58的田间杂草密度差异均不显著(P>0.05)(见图5). 比较田间各杂草种类,发现仅喇叭口期的苍耳表现为HGK60与郑58差异显著(P<0.05),其他生育期、其他物种均表现为二者差异不显著(P>0.05).
由图6可见,HGK60与郑58田间杂草群落Margelef指数、Shannon-Wiener指数、Simpson指数的变化趋势一致,且各生育期二者无显著差异(P>0.05);HGK60与郑58的Pielou指数变化范围在0.8~1.0之间,HGK60仅在苗期显著高于郑58(P<0.05),其他生育期二者差异不显著(P>0.05).
2.4 讨论
2.4.1转Cry1Ah基因抗虫玉米HGK60对根际土壤微生物的影响
该研究结果显示,HGK60种植后对根际土壤微生物的影响仅出现在某些生育期,并不会在整个生育期持续出现. 对门水平上相对丰度前10位根际土壤微生物的研究发现,厚壁菌门、变形菌门、酸杆菌门在各生育期均为优势种群,其中厚壁菌门与变形菌门的相对丰度均在抽穗期和完熟期表现为HGK60与郑58差异显著,其他生育期之间差异均不显著;酸杆菌门则在各生育期均表现为二者差异不显著. 玉米收获后前10个门均表现为HGK60与郑58差异不显著,认为HGK60的种植不会对玉米根际土壤微生物门水平产生持续且显著的影响. 厚壁菌门的内在物种具有降解土壤中难溶化合物[20]、固定空气中氮[21]、防治植物病虫害[22]等作用,具有较强的降解能力和代谢活性,其在抽穗期、完熟期HGK60根际土壤中的相对丰度显著高于郑58,表明该生长阶段HGK60的种植可有效增强土壤固氮、病虫害防治及代谢能力等. 变形菌门广泛分布于各类环境中,其在农业、环保、卫生等领域具有重要价值与作用[23-25]. 变形菌门的相对丰度在抽穗期、完熟期HGK60根际土壤中显著低于郑58,但收获后二者差异不再显著,表明HGK60的种植对变形菌门的影响过程相对较短且不持续.

表4 田间杂草数量调查结果Table 4 The number of each species in the fields 株

图5 田间杂草密度Fig.5 Weed density in the fields

图6 田间杂草各生态指标Fig.6 The ecological indexes of weeds in the fields
对前35个属的研究发现,同一生育期HGK60与郑58根际土壤微生物中金黄杆菌属、寡养单胞菌属、嗜糖假单胞菌属等的相对丰度存在差异,但亦只出现在某些生育期,并未在整个生育期持续出现. 值得注意的是,金黄杆菌属可降解苯胺、甲基叔丁基醚等难降解有机物[26];寡养单胞菌属对甲基对硫磷具有较高的降解性能[27];嗜糖假单胞菌属也具有降解有机物的作用[28]. 虽然以上物种均未对玉米根际土壤产生持续且显著的影响,但仍值得关注,以充分发挥其功效,增加其对土壤微环境的良性作用.
对根际土壤微生物alpha多样性的研究结果显示,HGK60种植对土壤微生物的影响主要存在于抽穗期与完熟期,收获后HGK60与郑58在Shannon-Wiener指数、Simpson指数、Chao1指数、PD指数上的差异均不显著;beta多样性研究亦发现,抽穗期、完熟期HGK60与郑58差异显著,苗期、喇叭口期、收获后二者均差异不显著,与alpha多样性的研究结果较为一致,均表明HGK60的种植虽在部分生育期对根际土壤微生物产生了显著影响,但该影响并未持续存在,玉米收获后HGK60与郑58间差异不显著. 已有学者对转基因作物根际土壤微生物开展了研究,但并未得到十分明确的结论. 部分研究认为转基因作物种植对根际土壤微生物无显著影响[29-34];但也有研究认为转基因作物的种植会对土壤微生物群落结构、多样性等产生显著影响[35-36]. 上述差异说明转基因作物种植对土壤微生物多样性的影响可能与作物类型、转入的外源基因、研究方法等有关,不可一概而论,需要进一步深入研究.
从生育期的角度分析玉米根际土壤微生物的动态变化,发现生育期的推进会对其产生一定影响. ANOSIM分析结果显示,HGK60与郑58均表现出在喇叭口期与抽穗期、完熟期与收获后两两生育期间的显著差异. 一些学者也得出了生育期会对作物根际土壤微生物产生一定影响的结论[37-39].
2.4.2转Cry1Ah基因抗虫玉米HGK60对田间节肢动物的影响
通过对田间节肢动物群落的研究发现,HGK60主要对螟蛾科昆虫有抑制作用. 从整体上看,HGK60种植对田间节肢动物的多个生态指标均未产生明显影响,差异仅出现在个别生育期,并未在整个生育期持续出现. 部分研究亦得出了转基因玉米种植不会对田间节肢动物产生显著影响的结论[7-9],与笔者所得结果较为一致. 但是也有研究得出了不同的结论,认为转基因抗虫作物的种植会在一定程度上造成田间天敌昆虫数量的下降以及非靶标昆虫数量的上升[40];也有部分研究认为转基因抗虫作物可能会提高田间节肢动物群落的丰富度和多样性,从而提高生态系统稳定性[41].
2.4.3转Cry1Ah基因抗虫玉米HGK60对田间杂草的影响
对田间杂草的研究结果显示,HGK60的种植对田间种类、数量及各生态学指标等均无显著影响,与赵思楠[42]得出的转G10evo-epsps基因和Cry1Ab/Cry2Aj融合基因抗虫耐草甘膦玉米双抗12-6对田间杂草多样性无显著影响的研究结果较为一致;刘来盘等[10]对抗除草剂大豆田间杂草进行调查,认为其对杂草群落组成与多样性也无显著影响. 但也有研究认为,转基因作物种植可改变田间单子叶、双子叶杂草比例[43],其竞争优势抢占生存空间及次生代谢发生改变,可能产生“化感”作用,抑制杂草生长,抗虫作物通过影响害虫取食习性变化也可对田间杂草产生间接影响[44]. 从已有研究结果来看,不同作物、不同转入基因等对田间杂草的研究结果差异较大,表明了转基因作物对环境压力反应的复杂性.
3 结论
a) 转Cry1Ah基因抗虫玉米HGK60种植对根际土壤微生物的影响仅出现在某些生育期,其对抽穗期、完熟期的影响相对较大,但该影响并不会在玉米的整个生长阶段持续出现,玉米收获后HGK60与郑58差异不显著(P>0.05);生育期是影响根际土壤微生物的重要因素之一.
b) 转Cry1Ah基因抗虫玉米HGK60对螟蛾科昆虫具有显著抑制作用(P<0.05),但对田间节肢动物的种类及Margalef指数、Shannon-Wiener指数、Simpson指数、Pielou指数等生态学指标均无显著影响(P>0.05).
c) 转Cry1Ah基因抗虫玉米HGK60种植对田间杂草密度及Margalef指数、Shannon-Wiener指数、Simpson指数、Pielou指数等生态学指标均无显著影响(P>0.05).
猜你喜欢
浙江农业科学(2022年7期)2022-07-15
中国土壤与肥料(2021年5期)2021-12-02
四川农业科技(2021年7期)2021-12-01
科学大众(中学)(2021年3期)2021-05-17
林业科技(2020年3期)2021-01-21
中国果业信息(2020年10期)2020-12-15
课外生活(小学1-3年级)(2018年10期)2018-11-19
湖北农业科学(2017年6期)2017-04-26
山东农业科学(2017年3期)2017-03-29
江苏农业科学(2016年10期)2017-02-05
