
遮光和施肥对苹婆幼苗生理特性和氮素积累的影响
2021-08-18 02:51潘陆荣周袁慧子王艺锦苏远玉王凌晖
湖北农业科学 2021年14期
潘陆荣,周袁慧子,王艺锦,苏远玉,王凌晖
(1.广西大学林学院,南宁 530004;2.广西民族大学建筑工程学院,南宁 530004;3.广西国有钦廉林场,广西 钦州 535000)
苹婆(Sterculia nobilisSmith)属梧桐科(Sterculiaceae)苹婆属(SterculiaLinn.),又名凤眼果、九层皮、频婆等,作为中国华南地区的乡土树种,是具有粮用、造林、景观绿化、生态修复等多种用途的优势常绿乔木,具有开发利用价值。针对苹婆的研究包括栽培现状[1]、栽培管理[2]以及发展前景[3]等方面,但是苹婆目前多为零星种植,未形成规模化种植,苹婆优质苗木及相关产品供不应求。
光照和施肥是影响植物生长的重要环境因子,植物通常会改变自身的形态以及一系列生理变化等过程来更好地适应光照变化,从而维持系统的平衡以及正常的新陈代谢活动[4]。前人对光照或施肥单一因素对植物影响做了大量研究,孟祥海[5]研究表明,适度遮阴能够缓解半夏膜系统受到的伤害,随着遮阴程度的增加,抗氧化酶活性呈先下降后升高的趋势,丙二醛含量则随着遮阴程度增加持续升高。谢美玲[6]研究表明,施氮水平提高有利于增加烤烟苗期叶片中过氧化氢酶(CAT)的活性,降低叶片丙二醛(MDA)的含量,有效地缓解了烤烟在高氮胁迫下的膜脂氧化现象。遮阴和施肥互作在提高了植物对光的利用效率以及对氮肥的吸收利用率方面具有重大作用。罗文姬[7]研究表明,相同氮素下,遮阴有利于缓解海南风吹楠苗木在全光照下的强光胁迫,明显提高SP、POD活性和SOD活性,苗木通过提高抗氧化酶活性清除活性氧来维持植株正常生长。韦艺[8]研究表明,施加相同氮素条件下,适度遮阴对闽楠幼苗的生理特性有促进作用,随着光照度的降低其生理特性表现出先升高后降低的趋势;相同遮阴条件下,适量添加氮对闽楠生理特性有促进作用,也呈现出低促高抑的相似规律。但是有关遮阴、施肥互作对苹婆的影响研究鲜见报道。目前,苗木生产过程中存在盲目施肥的现象,氮肥过量施用对促进植物生长无显著影响且降低了氮肥利用率,而且还会带来严重的环境污染问题[9]。因此,本研究针对苹婆幼苗进行不同遮光处理和氮肥条件控制,探索适合苹婆生长的光照和氮素,对提高氮肥利用率,促进苹婆种植业规模化、产业化发展提供参考。
1 材料与方法
1.1 试验地概况
试验地位于广西大学林学院苗圃教学实习基地(22°50′N,108°18′E),属亚热带季风气候,年均温21.5℃,年降雨量1 304.2 mm左右,年均相对湿度约为79%,光热充足,无霜期长,适宜苹婆植株的生长。
1.2 试验材料
试验材料为0.5年生苹婆幼苗,2014年12月初将苹婆幼苗(株高约5.0 cm,地径约3.5 mm)移植到23 cm(径)×25 cm(高)的有孔塑料盆,培养基质由体积比为3∶1的腐殖质土与苗圃熟土混合消毒而成,每盆1株苹婆幼苗,统一水肥管理,缓苗3个月后开始处理。2015年3月选取长势基本一致且健康无病虫害的苹婆幼苗160株,分为16组,每组10株,进行遮光、施肥处理。基地土壤为偏酸性赤红壤,有机质含量14.6 g/kg,全氮含量0.59 g/kg,全磷含量0.45 g/kg,全钾含量0.51 g/kg,碱性氮含量39.72 mg/kg,速效磷含量36.28 mg/kg,速效钾含量109.47 mg/kg,pH值为4.6。
1.3 试验方法
试验采用遮光、施肥双因素完全随机设计,各设置光照(L)和氮素(N)4个水平,组成16个处理组合(LmNn),每个处理10次重复,共160盆苹婆幼苗。通过搭建不同层数尼龙网进行遮光处理,营造4种不同光照环境,分别为自然光(NS,nature source light)的100.0%(L0)、41.3%(L1)、14.3%(L2)、3.6%(L3);4种氮素水平(以纯N含量计算),分别为0(N0)、2 000(N1)、4 000(N2)、6 000 mg/株(N3),试验氮素来源于尿素(N:46.67%)。从2015年3月15日开始每15 d处理1次,其余浇水、除草、松土等管理措施保持一致,具体处理如表1所示。

表1 试验设计处理及光氮水平
1.4 指标测定
分别于2015年3、6、9、12月采摘各处理成熟功能叶进行生理指标测定及植物叶片氮素积累量测定,各指标重复测定3次。其中,丙二醛(MDA)含量采用硫代巴比妥酸(TBA)法[10]测定;超氧化物岐化酶(SOD)活性采用NBT光化还原法[11]测定;过氧化物酶(POD)活性采用愈创木酚法[12]测定;过氧化氢酶(CAT)活性采用紫外分光光度法[13]测定;叶片全氮含量采用凯氏定氮法[12]测定。
1.5 数据处理
采用Excel 2010软件录入并整理试验数据、绘制图表,采用DPS软件进行方差分析,采用LSD方法进行多重比较。
2 结果与分析
2.1 遮阴和施肥对苹婆幼苗叶片丙二醛(MDA)含量的影响
由表2可知,光照度、氮素与两者交互对苹婆幼苗6、9、12月MDA含量均产生极显著的影响(P<0.01),其效应强弱表现为光>氮>光×氮,即以光照度的影响最大。如图1所示,3月平均MDA含量为9.44 μmol/g(FW),随着时间的推移,各处理MDA含量随之增加,在12月达到最大值,平均MDA含量为29.68 μmol/g(FW),是3月的3.14倍,6月、9月平均MDA含量分别为14.43、25.34 μmol/g(FW)。
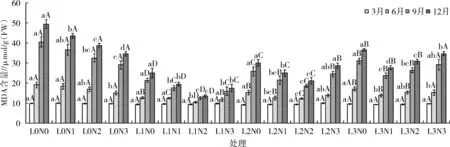
图1 苹婆幼苗不同时期丙二醛(MDA)含量动态变化
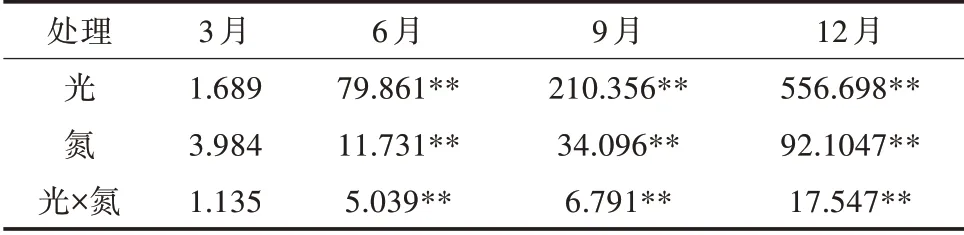
表2 光氮互作对苹婆幼苗丙二醛含量影响的显著性F测验结果
随着遮光程度加强,光照减弱,6、9、12月MDA含量整体呈先下降后升高的趋势,均以L0N0处理最高,分别为18.97、40.46、49.27 μmol/g(FW),以L1N2处理最低,分别为10.31、12.47、13.31 μmol/g(FW),整体排序为L0>L3>L2>L1。在全光照条件下,丙二醛含量随氮素增加不断减少,在各遮阴处理下,丙二醛含量先降后升,均以N0施氮水平最高;在同一氮素水平下,丙二醛含量均随光照度减弱呈先下降后增加的变化趋势,丙二醛含量均以L1光照处理最低,N0、N1、N2氮素水平下以L0光照处理最高,N3氮素水平下以L3光照处理最高。可见,光照过强或过弱,氮素水平过多或缺乏都显著引起MDA含量增加,适当遮阴和施肥有利于降低MDA含量,减少苹婆幼苗受到的伤害,且存在光氮互作效应,以L1N2处理表现较佳。
2.2 遮阴和施肥对苹婆幼苗叶片超氧化物歧化酶(SOD)活性的影响
由表3可知,光照度、氮素与两者交互对苹婆幼苗6、9、12月SOD活性均产生极显著的影响(P<0.01),其中,氮素为主导因素,光氮互作次之,最后为光照单独作用。如图2所示,SOD变化在不同季节表现不同,整体趋势为9月>6月>12月>3月。3月平均SOD活性为289.75 U/g(FW);经过施N肥和遮阴处理后SOD活性变化较大,9月SOD平均活性最高,为549.25 U/g(FW),6月、12月SOD平均活性分别为487.19、481.85 U/g(FW)。
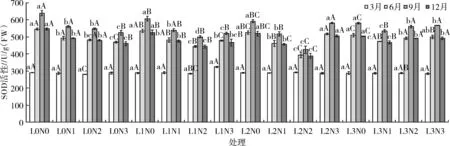
图2 苹婆幼苗不同时期超氧化物歧化酶(SOD)活性动态变化

表3 光氮互作对苹婆幼苗SOD活性影响的显著性F测验结果
随着光照度逐渐降低,6、9、12月SOD活性整体呈“V”字型变化,大小排序为L0>L3>L1>L2。在L0光照条件下,随着氮素增加,6、9、12月SOD活性不断下降,均在N0施氮水平下达到最大值,而L1、L2、L3光照条件下,SOD活性均表现为先降后升的趋势,同样在N0施氮水平下达到最大值。在N0氮素水平下,随着光照度降低SOD活性不断降低,在N1、N2氮素水平下SOD活性先降后升,在N3氮素水平下SOD活性整体呈上升趋势。总体来说,L2N2处理下SOD活性最小,L0N0处理下SOD活性最大,说明光照过强、缺少氮素不利于植物生长,导致SOD活性升高来保护植物,适当遮阴、施肥可减少细胞膜系统受到的伤害。
2.3 遮阴和施肥对苹婆幼苗叶片过氧化氢酶(CAT)活性的影响
由表4可知,光照度、氮素与两者交互对苹婆幼苗6、9、12月CAT酶活性均产生极显著的影响,6、9、12月光强、氮素与光氮互作对苹婆幼苗CAT酶活性的影响大小顺序为光>光×氮>氮。如图3所示,3月平均CAT活性最低,为197.41 U/(g·min)(FW),6月CAT平均活性为368.85 U/(g·min)(FW),12月CAT平均活性为415.08 U/(g·min)(FW),9月CAT平均活性活性达到最高值,为447.01 U/(g·min)(FW)。

图3 苹婆幼苗不同时期过氧化氢酶(CAT)活性动态变化
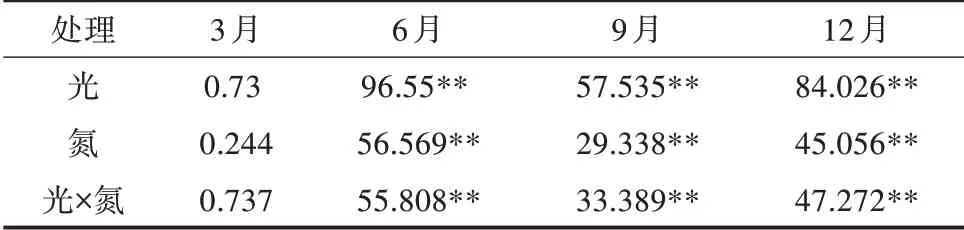
表4 光氮互作对苹婆幼苗CAT活性影响的显著性F测验结果
随着光照度减弱CAT活性呈“降-升-降”的变化趋势,CAT活性大小排序为L2>L0>L3>L1。在L0、L1光照条件下,随着氮素的增加,6、9、12月CAT酶活性均在N1施氮水平下达到最大值,而L2、L3光照条件下,CAT酶活性均在N3施氮水平下达到最大值。在N0、N1氮素水平下,CAT酶活性在L0光照条件下达到最大,N2、N3氮素水平下CAT酶活性分别在L2、L3光照条件下较大。总体而言,CAT酶活性在L0N1处理下达到最大值,L0N3处理下最小。
2.4 遮阴和施肥对苹婆幼苗叶片过氧化物酶(POD)活性的影响
由表5可知,光照度、氮素与两者交互作用对苹婆POD活性的影响在6、9、12月均达极显著水平(P<0.01),其影响大小顺序为光>氮>光×氮。如图4所示,POD活性变化在不同季节表现不同,9月POD平均活性最高,为354.45 U/(g·min)(FW),其次为12月、6月,分别为303.73、152.64 U/(g·min)(FW),而3月最低,为100.84 U/(g·min)(FW)。
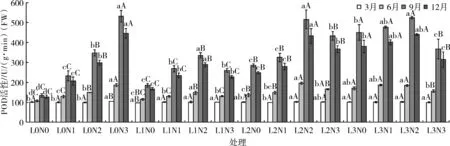
图4 苹婆幼苗不同时期过氧化物酶(POD)活性动态变化
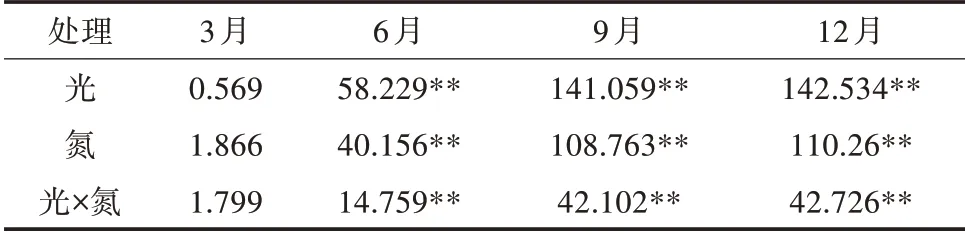
表5 光氮互作对苹婆幼苗POD活性影响的显著性F测验
随着遮阴网加厚、光照度减弱,POD活性呈先下降后上升的趋势,不同光照度POD活性排序为L3>L2>L0>L1。在全光照条件下,6、9、12月POD活性随氮素增加而增加,均以N3施氮水平最高;L1、L2光照处理下,POD活性随氮素增加先升后降,均以N2施氮水平最高,L3光照处理下也表现出相似趋势。在N0、N1氮素水平下,POD活性均以L3光照度条件下最高,N2氮素水平下,9、12月POD活性以L3光照度下最高,N3氮素水平下则以L0光照条件下最高。总体来看,L0N3处理下POD活性最高,L0N0处理下POD活性最小。说明适当遮阴施肥有利于提高细胞抗氧化能力,维持自由基的产生与清除之间的平衡,提升植物抗逆性。
2.5 遮阴和施肥对苹婆幼苗叶片N素浓度的影响
由表6可知,光照度、氮素与两者交互作用对苹婆N素浓度的影响在6、9、12月均达极显著水平(P<0.01),且影响趋势为氮>光>光×氮,氮对苹婆N素浓度的影响显著大于光的影响。如图5所示,3、6、9、12月平均叶片N素浓度表现为先上升后下降,大小排序为9月>6月>12月>3月。其中,3、6、9、12月N素浓度分别为10.21、18.79、21.22、16.88 mg/g。

表6 光氮互作对苹婆幼苗叶片N素浓度影响的显著性F测验
施N肥和遮阴处理后叶片N素浓度变化较大,在相同光照处理下,6、9、12月平均叶片N素浓度均随着氮素的增加而增加,均是N0水平的苹婆幼苗叶片N素浓度最小,在不同光照下苹婆幼苗叶片N素浓度大小排序为L2>L1>L3>L0。相同氮素水平,在一年中不同季节叶片N素浓度不同光照处理下反应不同,当氮素水平分别为N0、N1时,随着遮光强度增加,6月、12月氮浓度均呈先升后降的趋势,而9月氮浓度变为先降后升;在N2、N3氮素水平下,随着遮光强度增加,6月N素浓度不断下降,而12月N素浓度先升后降。总体而言,在L1N3处理下取得最大值,在L3N0处理下最小。
2.6 遮阴和施肥对苹婆幼苗生理指标及养分特征的隶属分析
根据隶属值大小可以将植物对环境的适应程度分为4个等级[14]:高抗型(1≥隶属值≥0.6)、中抗型(0.6>隶属值≥0.4)、弱抗型(0.4>隶属值≥0.2)、不抗型(隶属值<0.2)。由表7可知,苹婆幼苗平均隶属值为0.524,属于中抗型,表明苹婆对环境有较好适应能力、抗性,以及对病虫害、生物侵蚀等的耐力。在隶属函数模糊综合评价分析下,苹婆幼苗的隶属值最大的是L1N3,说明该光氮条件下苹婆的苗木综合情况最好,其次是L1N2,隶属值最小的是L0N0,光照过强、光照不足、缺少氮肥均不利于苹婆生长,适度遮阴(41.3%自然光)和施加氮肥(6 000 mg/株)能有效促进苹婆幼苗生长发育。
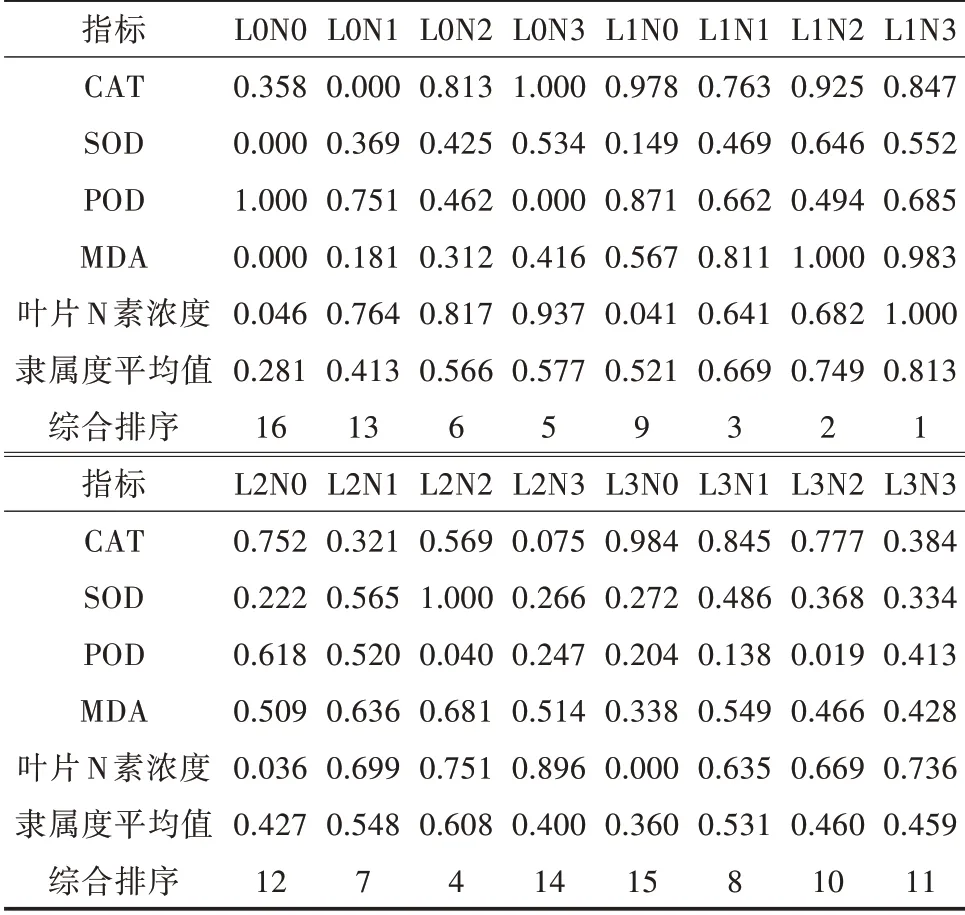
表7 苹婆幼苗在不同光氮条件下生理指标及养分特征的隶属分析
3 小结与讨论
光是影响和调节植物抗氧化能力的重要因子,逆境条件下,植物内部氧自由基积累量增加,膜脂过氧化加剧,导致膜系统受到损伤,影响植物的生长[15]。本研究结果表明,MDA含量随试验时间的延长而不断增加,在12月达到最大值,不同处理下,叶片中MDA含量差异明显,光照过强或过弱,氮素水平过多或缺乏都显著引起MDA含量增加,适当遮阴和施肥有利于降低MDA含量,减少苹婆幼苗受到伤害,这与何静雯等[16]对鄞红葡萄(Vitis vinifera)的研究结果相似。
逆境环境下,植物会启动一系列保护酶调整和降低活性氧含量[17],SOD、CAT和POD是植物体内重要保护酶,植物在逆境下的适应能力也可以根据其活性变化来反映,3种保护酶活性均在9月最高,3月最低,SOD季节变化整体表现为9月>6月>12月>3月,而POD、CAT季节变化整体均表现为9月>12月>6月>3月。叶片SOD、POD、CAT虽然都属于保护酶,但活性变化在不同光氮条件下表现也不同,SOD活性随着光照度逐渐降低整体呈“V”字型变化,在全光照条件下随氮素增加而不断下降,说明SOD活性在强光和弱光条件下表现比较活跃,SOD活性在L0N0处理下达到最大值,施加氮素缓解逆境伤害从而使SOD活性降低,L2N2处理下SOD活性最低,说明适当遮阴、施肥最有利于植物生长。随着光照度减弱,POD活性呈先下降后上升的趋势,在L3光照条件下达到最大,POD对不同氮素水平的反应与SOD酶活性不同,适量的施用氮素反而能提高POD的活性,这与丁雪梅等[18]研究结果相似,施用氮肥可显著提高过氧化物酶活性,且随施氮量的增加,抗氧化酶活性不同程度地增加。说明可以通过施加氮素提高叶片酶活性来清除叶片中氧自由基,降低逆境对植物的危害。
在本研究中随着光照度减弱,CAT活性变化整体呈波浪形,大小排序依次为L2>L0>L3>L1,CAT活性随着氮素水平的变化反应不同。在L0、L1光照条件下,CAT酶活性均在N1施氮水平下达到最大值,而L2、L3光照条件下,CAT酶活性均在N3施氮水平下达到最大值,体现光氮互作效应,植物在适当的胁迫下能提高CAT活性,但胁迫过多,反而不利于CAT酶的生成[19]。酶活性对不同光氮处理变化不同,总体来说L1光照处理下、N3氮素水平下3种酶活性相对于其他光照条件、氮素水平偏低。研究表明,适量施氮更有利于植物生长和氮素贮藏。张永发等[20]研究发现,橡胶树(Hevea brasiliensis)氮肥施用量直接影响贮藏氮的积累,不同供氮量贮藏氮水平各异,适量施氮更有利于贮藏氮的积累,尤其是主干的贮藏氮积累。本研究中,苹婆幼苗施氮后叶片氮素含量显著提升,叶片N素浓度在一年中9月>6月>12月>3月,相同光照条件下,均是N0水平的苹婆幼苗叶片N素浓度最小,各光氮处理下平均叶片N素浓度变化均随着氮素水平的增加,叶片N素浓度不断上升。
通过对苹婆幼苗各指标进行隶属函数综合评价得出,苹婆幼苗隶属值最大的是L1N3。综上所述,光照过强、光照不足、缺少氮肥均不利于苹婆生长,适度遮阴(41.3%自然光)和施加氮肥(6 000 mg/株)能有效促进苹婆幼苗生长发育。
猜你喜欢
传感器世界(2022年7期)2022-10-12
中国机械工程(2022年8期)2022-05-09
中国机械工程(2021年8期)2021-05-07
音乐教育与创作(2019年8期)2019-05-16
农业工程技术·温室园艺(2016年9期)2017-04-23
河南科技(2014年4期)2014-02-27
植物营养与肥料学报(2011年5期)2011-11-06
植物营养与肥料学报(2011年2期)2011-10-26
植物营养与肥料学报(2011年3期)2011-10-24
植物营养与肥料学报(2010年3期)2010-11-16
