
吡虫啉对异迟眼蕈蚊抗性筛选及其对解毒酶活性的影响
2021-09-04 07:44沈登荣袁盛勇
河南农业科学 2021年7期
张 睿,何 超,李 河,沈登荣,袁盛勇,李 珣
(红河学院 生命科学与技术学院/云南省高校滇南特色生物资源研究与利用重点实验室,云南 蒙自 661199)
异迟眼蕈蚊(Bradysia difformisFrey)是一种世界性的农、林业害虫[1],以幼虫取食危害,造成食用菌出菇量显著下降,引起植物体内营养和水分运输受阻,严重影响农作物的产量和质量[2-3]。该虫自2009年首次在我国云南省栽培食用菌上发现以来,目前已在甘肃、广西、江苏、浙江等省(区)的食用菌栽培区造成危害[3-5]。在甘肃天水地区还发现该虫是韭菜种植区的优势种,常与韭菜迟眼蕈蚊(Bradysia odoriphaga)混合发生[3]。由于该虫世代周期短、繁殖力强、具有钻蛀危害等习性,目前在生产中主要采取化学药剂对其进行防治。吡虫啉是一种新烟碱类的高效杀虫剂,目前已广泛用于大田作物、蔬菜、果树和花卉植物上的害虫防治[6]。研究表明,吡虫啉是一种对食用菌、韭菜等作物安全性高、残留量低,对眼蕈蚊科害虫具有高效作用的杀虫剂[7-10]。随着杀虫剂的频繁使用,国内外已有眼蕈蚊抗药性的研究报道,如美国宾夕法尼亚州和特拉华州的厉眼蕈蚊(Lycoriella mali)已对苄氯菊酯产生了高抗性[11]。我国7个韭菜主产区的韭菜迟眼蕈蚊(B.odoriphaga)均对辛硫磷、毒死蜱等有机磷类杀虫剂产生了抗性,部分主产区对吡虫啉、噻虫嗪、噻虫胺等新烟碱类杀虫剂产生了中、高抗性[12-13]。
在昆虫抗药性机制中,昆虫解毒酶代谢作用的增强是昆虫对杀虫药剂产生抗药性的重要机制之一。研 究 发 现,棉 蚜(Aphis gossypii)[14]、西 花 蓟 马(Frankliniellaoccidentalis)[15]、褐 飞 虱(Nilaparvata lugens)[16]等昆虫体内多功能氧化酶(O-脱甲基,MFO)、羧酸酯酶(CarE)、谷胱甘肽S-转移酶(GST)活性增强是昆虫对吡虫啉代谢抗性形成的主要原因。异迟眼蕈蚊是一类腐食性昆虫,可在设施栽培作物(食用菌、温室花卉等)和大田作物(韭菜、葱等)之间交替危害。近年来,吡虫啉已逐渐成为我国蔬菜上的骨干药剂,目前国内外尚无异迟眼蕈蚊对吡虫啉抗性的研究报道。鉴于此,采用吡虫啉对异迟眼蕈蚊进行抗性筛选,分析吡虫啉亚致死剂量对异迟眼蕈蚊敏感和抗性品系MFO、CarE、GST活性的影响,旨在评价异迟眼蕈蚊对吡虫啉产生抗药性的潜在风险,探讨其对吡虫啉代谢抗性形成的生化机制,为该虫的抗性预防和治理提供理论依据。
1 材料和方法
1.1 供试虫源
异迟眼蕈蚊于2014年采自云南省蒙自市文澜镇平菇上,在室内采用灭菌的腐殖土作为饲养基质进行连续饲养,幼虫期添加2~4次黄豆粉(购于云南省蒙自市文澜镇农贸市场)作为营养补充。在饲养期间不接触任何药剂,已连续饲养4 a。饲养和试验处理条件:温度为(25±1)℃、相对湿度为(70±5)%、光周期为16L∶8D。
1.2 供试试剂
供试杀虫剂:70%吡虫啉水分散粒剂(拜耳作物科学有限公司)。主要化学试剂:α-乙酸萘酯、毒扁豆碱、固蓝B盐、十二烷基硫酸钠、还原型谷胱甘肽(GSH)、1-氯-2,4-二硝基苯(CDNB)、对硝基苯酚、牛血清蛋白、考马斯亮蓝G250、还原型辅酶Ⅱ(NADPH)、对硝基苯甲醚,均购于上海阿拉丁生化科技公司。
1.3 试验方法
1.3.1 吡虫啉对异迟眼蕈蚊的毒力测定 采用慕卫等[17]的胃毒触杀联合毒力法,略做改动。采用蒸馏水将吡虫啉稀释为5个质量浓度,设置蒸馏水为对照。取晒干黑木耳(购于云南省蒙自市文澜镇农贸市场)3~4片放入各处理药液中浸泡30 min,待木耳泡开后用圆形打孔器(直径3.5 cm)打取木耳圆片,将木耳圆片放入50 mL塑料瓶(直径4 cm,高6.3 cm)中,每瓶1片。挑选个体大小一致的三龄幼虫30头放在木耳上,用纱布封口,各处理设3次重复。处理后的幼虫采取黑暗处理,置于光照培养箱中饲养,24 h后统计幼虫死亡数。
1.3.2 异迟眼蕈蚊对吡虫啉的抗性筛选 采用单体筛选法进行敏感品系和抗性品系的分离。将羽化12 h以内的单对成虫放入产卵杯(直径8.5 cm,高11 cm)中,杯内预先放置3%水琼脂用来收集卵粒,幼虫孵化后转移至灭菌的腐殖土中饲养繁殖1代,建立20个稳定种群。根据1.3.1结果,采用5.0 mg/L吡虫啉(死亡率10%~15%)处理各种群3龄幼虫,处理方法同1.3.1。挑选死亡率较高的2~3个种群混合饲养作为敏感种群正常饲养(期间不接触任何化学农药)。挑选死亡率较低的2~3个种群混合饲养作为抗性种群进行抗性筛选,采用吡虫啉(选择上一代幼虫死亡率为40%~50%的质量浓度)处理三龄幼虫,将幼虫放入药液中浸渍30 s,用相同浓度药液处理灭菌腐殖土用于幼虫饲养,期间添加1~2次黄豆粉,至三龄幼虫发育至成虫。将羽化12 h的成虫接入不含药剂的腐殖土中饲养至三龄幼虫,作为下一代抗性筛选的虫源,每3代进行1次毒力测定,完成21代的抗性筛选。
1.3.3 抗性现实遗传力和抗性风险评估 采用TABASHNIK等[18]的阈性状分析法计算抗性现实遗传力h2,参照王建军等[19]方法进行抗性风险评估。
1.3.4 异迟眼蕈蚊解毒酶活性测定 采用1.3.1的测定方法计算吡虫啉对异迟眼蕈蚊敏感品系和抗性品系三龄幼虫的亚致死剂量LC10、LC20和LC40(即引起三龄幼虫死亡率分别达到10%、20%和40%的药剂质量浓度)。采用亚致死剂量LC10、LC20和LC40分别处理敏感品系和抗性品系异迟眼蕈蚊三龄幼虫,设置蒸馏水为空白对照,各质量浓度处理90头三龄幼虫,3次重复,分别于24、48、72 h后收集各质量浓度处理存活的三龄幼虫,用于解毒酶活性的测定与分析。MFO、CarE、GST活性测定参照余慧灵等[20]的方法,单位分别为nmol/(min·mg)、mmol/(min·mg)、mmol/(min·mg)。酶原蛋白质含量测定参照BRADFORD等[21]的考马斯亮蓝G-250测定法。
1.4 数据处理
采用SPSS 19.0进行数据分析,计算毒力回归方程、致死中质量浓度(LC50)、亚致死剂量和95%置信限,采用Duncan’s法进行差异显著性分析。
2 结果与分析
2.1 利用吡虫啉对异迟眼蕈蚊抗性的筛选结果
从表1可看出,经过21代的抗性筛选,吡虫啉对异迟眼蕈蚊三龄幼虫的致死中质量浓度LC50由13.302 mg/L上升到54.449 mg/L,抗性倍数为4.09倍。室内筛选21代后该虫处于敏感性下降水平。从抗性选育中可看出,F0—F9代的抗性倍数增长缓慢;F9—F21代的抗性倍数增长较快,抗性水平呈现逐渐增长的趋势。
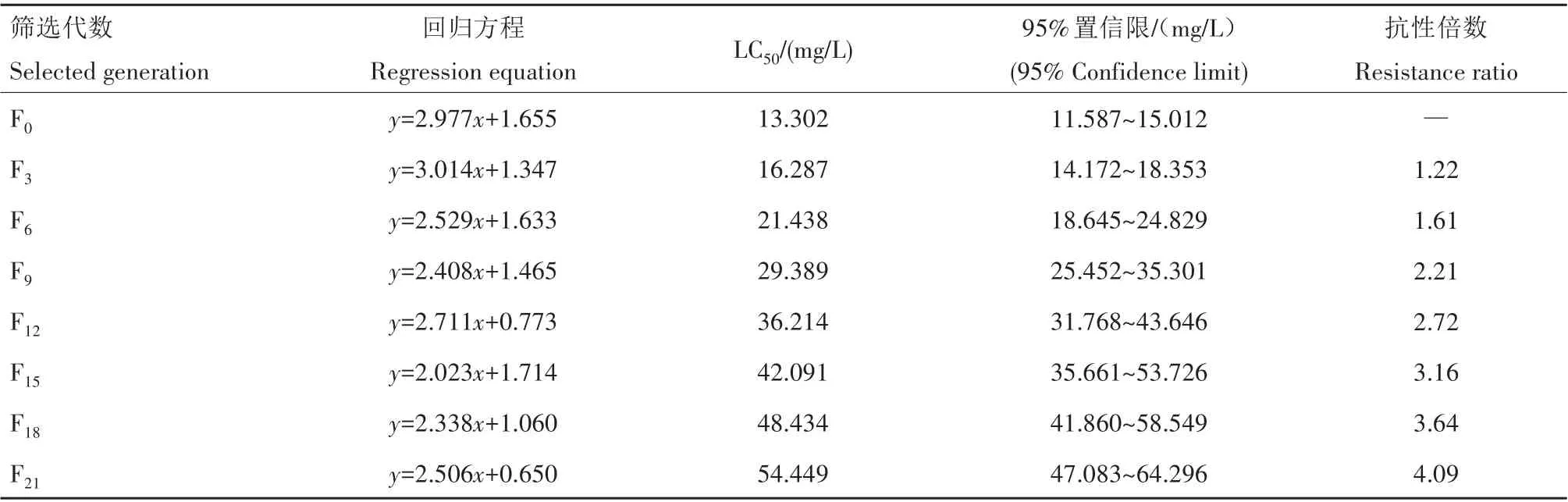
表1 利用吡虫啉对异迟眼蕈蚊抗性的筛选结果Tab.1 Resistance selection of B.difformis to imidacloprid
2.2 异迟眼蕈蚊对吡虫啉的抗性现实遗传力和发展速率预测
从表2可看出,吡虫啉对异迟眼蕈蚊室内连续筛选21代后,其抗性现实遗传力(h2)为0.091 3,毒力回归方程的平均斜率为2.555 3,表现型标准方差δp为0.391 3。假定以吡虫啉致异迟眼蕈蚊的死亡率分别为50%、60%、70%、80%、90%、99%作为药剂选择压力,预测抗性提高10倍所需的代数如图1所示。随着吡虫啉选择压力的增加,异迟眼蕈蚊抗性水平增长10倍所需代数逐渐减少,选择压力在50%~70%时,抗性增加10倍需要24.1~35.1代;选择压力在80%~90%时,抗性增加10倍需要15.9~20.1代。说明异迟眼蕈蚊对吡虫啉产生抗性的风险较低。

图1 不同选择压力下异迟眼蕈蚊对吡虫啉抗性提高10倍所需代数Fig.1 Generations for developping 10-fold resistance of B.difformis to imidacloprid under different selection pressures
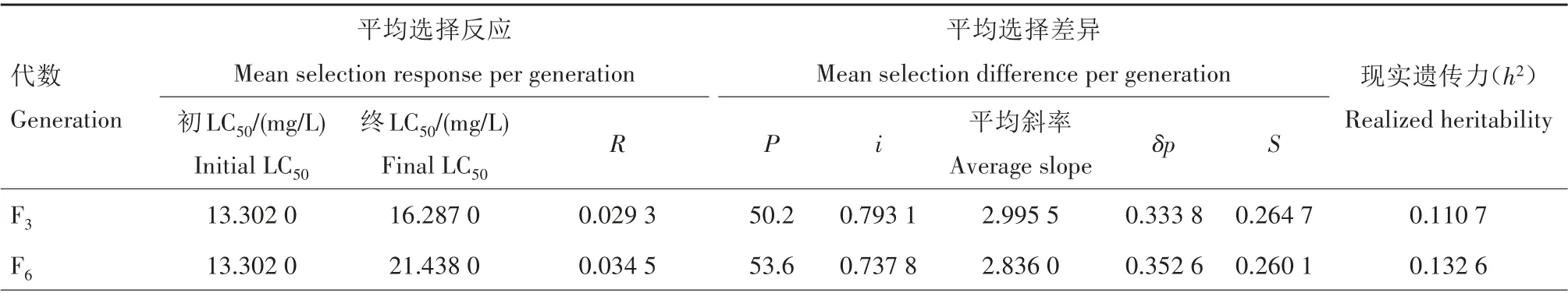
表2 异迟眼蕈蚊对吡虫啉的抗性现实遗传力Tab.2 Realized heritability of the resistance of B.difformis to imidacloprid

续表2 异迟眼蕈蚊对吡虫啉的抗性现实遗传力Tab.2(Continued) Realized heritability of the resistance of B.difformis to imidacloprid
2.3 吡虫啉亚致死剂量对敏感和抗性品系异迟眼蕈蚊多功能氧化酶O-脱甲基活性的影响
从图2可知,随着吡虫啉亚致死剂量的提高,敏感和抗性品系异迟眼蕈蚊的多功能氧化酶O-脱甲基活性逐渐增大,其中敏感品系在亚致死剂量LC20和LC40处理下多功能氧化酶O-脱甲基活性显著高于空白对照(P<0.05),抗性品系在亚致死剂量LC10、LC20和LC40处理下多功能氧化酶O-脱甲基活性均显著高于空白对照(P<0.05)。敏感和抗性品系异迟眼蕈蚊的多功能氧化酶O-脱甲基活性在亚致死剂量LC10、LC20和LC40处理24~72 h呈现先升高后下降的趋势,均在48 h达到峰值,敏感和抗性品系在LC40处理48 h时酶活性较空白对照分别上升了63.81%和119.42%。表明吡虫啉亚致死剂量对抗性品系多功能氧化酶O-脱甲基活性有较强的提升作用,该酶活性的增强与异迟眼蕈蚊对吡虫啉的代谢抗性有关。

图2 吡虫啉亚致死剂量对异迟眼蕈蚊多功能氧化酶O-脱甲基活性的影响Fig.2 Effectsof sublethal doses of imidacloprid on MFO O-demethylation activity in B.difformis
2.4 吡虫啉亚致死剂量对敏感和抗性品系异迟眼蕈蚊谷胱甘肽S-转移酶活性的影响
从图3可知,随着吡虫啉亚致死剂量的提高,敏感和抗性品系异迟眼蕈蚊的谷胱甘肽S-转移酶活性逐渐提高,同一处理时间不同处理浓度以及不同处理时间同一处理浓度下各品系异迟眼蕈蚊谷胱甘肽S-转移酶活性与空白对照均无显著差异,且该酶在敏感和抗性品系间变化水平也较小。表明吡虫啉亚致死剂量对谷胱甘肽S-转移酶活性无明显的诱导作用。
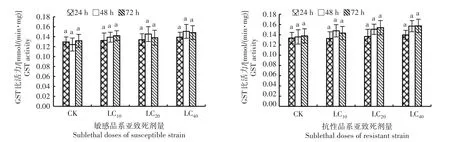
图3 吡虫啉亚致死剂量对异迟眼蕈蚊谷胱甘肽S-转移酶活性的影响Fig.3 Effects of sublethal doses of imidacloprid on GST activity in B.difformis
2.5 吡虫啉亚致死剂量对敏感和抗性品系异迟眼蕈蚊羧酸酯酶活性的影响
从图4可知,随着吡虫啉亚致死剂量提高,敏感和抗性品系异迟眼蕈蚊的羧酸酯酶活性逐渐增大,其中敏感品系在亚致死剂量LC20和LC40处理48 h的羧酸酯酶活性显著高于空白对照(P<0.05);抗性品系除LC10处理24 h外,其余各浓度处理不同时间的羧酸酯酶活性均显著高于空白对照(P<0.05)。各处理酶活性均在处理48 h达到峰值,敏感和抗性品系在LC40处理48 h时酶活性较空白对照分别上升了18.40%和48.13%。表明吡虫啉亚致死剂量对抗性品系羧酸酯酶活性具有较强的提升作用,该酶活性的增强与异迟眼蕈蚊对吡虫啉的代谢抗性有关。
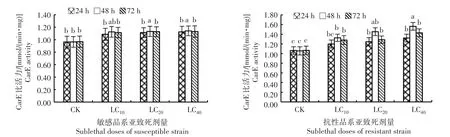
图4 吡虫啉亚致死剂量对异迟眼蕈蚊羧酸酯酶活性的影响Fig.4 Effects of sublethal doses of imidacloprid on Car E activity in B.difformis
3 结论与讨论
本研究发现,经过室内抗性筛选21代后,异迟眼蕈蚊对吡虫啉的抗性倍数增长了4.09倍,筛选后的异迟眼蕈蚊抗性仍处于敏感性下降阶段。目前,国内外尚无异迟眼蕈蚊对吡虫啉抗性的研究报道,但在国内已有韭菜迟眼蕈蚊对吡虫啉等烟碱类杀虫剂产生抗药性的研究报道[12-13]。究其原因,主要是近年在我国北方主要韭菜种植区大面积推广使用吡虫啉等烟碱类杀虫剂控制韭菜迟眼蕈蚊所致[13],各地区因用药频率的差异表现出对吡虫啉不同的抗性水平,而在食用菌中使用吡虫啉控制菇蚊、菇蝇的时间还较短,且食用菌生长周期短,频繁使用化学杀虫剂极易引起农药残留[22],因此在食用菌上吡虫啉对害虫的药剂选择压力较小。另外,韭菜属于多年生的蔬菜植物,而食用菌属于阶段性种植的蔬菜,两者在用药浓度和频率上也存在较大差异。
昆虫抗药性风险评估是在抗性选育基础上,依据抗性现实遗传力预测不同选择压力下昆虫的抗性发展速率。该方法已用于许多昆虫抗性的风险评估,例如溴氰菊酯对异迟眼蕈蚊选择压力在80%~90%时,抗性增加10倍需要11~14代[23];吡虫啉对棉蚜选择压力在80%~90%时,抗性增加100倍需要30~38代[24];吡虫啉对白背飞虱(Sogatella furcifera)选择压力在80%~90%时,抗性增加10倍仅需4~5代[25];吡虫啉对新疆烟粉虱(Bemisia tabaci)隐种选择压力在70%~90%时,抗性增加10倍仅需4~6代[26]。本研究表明,当吡虫啉对异迟眼蕈蚊选择压力在80%~90%时,抗性增加10倍需要15.9~20.1代,说明异迟眼蕈蚊对吡虫啉产生抗性的风险较低。考虑到该虫食性较杂,可在设施栽培作物(食用菌、温室花卉等)和大田作物(韭菜、葱等)之间交替危害,为避免该虫过早出现抗性问题,在不同作物间作中也要尽量降低吡虫啉的施用量和使用频率。
研究证实,昆虫解毒酶代谢作用的增强是昆虫对杀虫药剂产生抗药性的重要机制之一[6]。本研究表明,吡虫啉亚致死剂量能显著提升异迟眼蕈蚊抗性品系多功能氧化酶O-脱甲基和羧酸酯酶的活性,在吡虫啉各亚致死剂量处理下异迟眼蕈蚊抗性品系多功能氧化酶O-脱甲基和羧酸酯酶均在处理后48 h达到峰值,表明上述2种解毒酶活性的增强与异迟眼蕈蚊对吡虫啉抗性的提高有密切关系。这与崔丽等[14]研究棉蚜抗吡虫啉品系和敏感品系的多功能氧化酶O-脱甲基和羧酸酯酶活性、王圣印等[15]研究西花蓟马抗吡虫啉品系和敏感品系的羧酸酯酶活性、WEN等[27]研究褐飞虱抗吡虫啉品系和敏感品系的多功能氧化酶活性的结果相似。本研究结果也为生产上利用增效醚(多功能氧化酶抑制剂)、磷酸三苯酯(羧酸酯酶抑制剂)增强吡虫啉对异迟眼蕈蚊抗性种群的防治效果提供了理论依据。在后续研究中,还需从2种解毒酶的抗性基因和表达水平深入研究,以全面揭示异迟眼蕈蚊对吡虫啉的代谢抗性形成及相关机制,为异迟眼蕈蚊对吡虫啉的抗性预防和吡虫啉的科学使用提供科学依据。
猜你喜欢
农产品加工(2022年15期)2022-09-26
湖南农业科学(2022年1期)2022-06-06
广东农业科学(2022年1期)2022-03-03
广西蚕业(2021年3期)2021-10-28
水产科学(2021年4期)2021-07-21
当代水产(2021年1期)2021-03-19
农药科学与管理(2021年2期)2021-03-16
四川蚕业(2021年4期)2021-03-08
世界热带农业信息(2018年3期)2018-09-26
热带作物学报(2018年5期)2018-05-14
