
转BpGLK1基因白桦叶色变异规律及生长特性分析
2021-09-08 01:11刘佳琦宋逸欣成星川金冬雪
江西农业学报 2021年8期
刘佳琦,宋逸欣,成星川,王 庆,金冬雪,姜 静
(东北林业大学 林木遗传育种国家重点实验室,黑龙江 哈尔滨 150040)
Golden2-like(GLK)转录因子,是一类植物中广泛存在的转录因子,属于GARP转录因子家族,GLK主要调控植物的叶绿体发育、影响果实品质,同时也参与植物生物胁迫和非生物胁迫、植物衰老和激素信号转导等[1-3]。研究证明,不同植物中GLK家族成员功能不尽相同,例如,拟南芥(Arabidopsisthaliana)AtGLK1蛋白可以直接结合到一些天线蛋白和叶绿素合成酶等基因的启动子上并促使其表达量在转录水平上提高[4]。有些植物的GLK成员则在不同组织中起作用,例如,玉米(ZeamaysL.)ZmGLK1和ZmGLK2基因分别特异性地在束鞘和叶肉细胞中起作用,调控束鞘和叶肉细胞中的二型叶绿体的发育[4]。番茄(Solanumlycopersicum)和辣椒(Capsicumfrutescens)的SlGLK1和CaGLK1主要在叶片中表达;SlGLK2和CaGLK2在果实中起重要作用[2,5]。有研究证明:SlGLK2基因能够明显增强番茄未成熟果实叶绿体的功能, 提高果实叶绿素的含量,从而通过过表达SlGLKs基因能够在一定程度上提高番茄果实的可溶性固形物含量[6]。白桦(BetulaplatypHyllaSuk.)基因组中GLK成员仅有1个(BpGLK1)[7,8]。研究团队已证明,白桦BpGLK1基因的缺失突变株及BpGLK1干扰表达转基因株系均表现出叶片叶绿素含量降低,叶片呈现黄绿色[8,9],将银中杨(Populusalba×P.berolinensis)PaGLK1基因干扰表达后,转基因株系同样呈现亮黄绿叶色,上述研究证明,GLK基因是创制黄叶植物的首选基因。
白桦做为园林绿化的观赏树种,被广泛应用于城市的道路、公园、庭院及小区绿化[10]。白桦BpGLK1干扰表达株系的叶色在春、夏、秋季节持续呈现醒目的亮黄绿色,提升了白桦的观赏价值,其应用前景更加广阔。然而,BpGLK1干扰表达株系叶片叶绿素含量的降低对其生长是否产生影响,目前尚未研究。为此,在获批转BpGLK1基因白桦中间试验行政许可的基础上(林技许准字[2017]12号),本试验以前期获得的7个BpGLK1干扰表达株系、3个BpGLK1过表达株系为材料,测定了叶绿素相对含量与叶色的时序变异规律,分析了转基因白桦的生长特性,旨在为后续转BpGLK1白桦的生产性试验提供指导。
1 材料与方法
1.1 材料
植物材料:2年生的转BpGLK1基因白桦(Betulaplatyphylla×B.pendula),包括BpGLK1过表达株系(OE1、OE2、OE3)、BpGLK1干扰表达株系(RE1、RE2、RE3、RE4、RE5、RE6、RE7)、非转基因野生型白桦(WT),每个株系30株,种植于30 cm×20 cm花盆中,置于白桦育种基地。
主要质粒载体:过表达载体p35S::GLK,干扰表达载体p35S::GLK-RNAi,由研究团队保存。
1.2 方法
1.2.1 转基因白桦的分子检测BpGLK1过表达株系的PCR检测:分别提取参试株系的总DNA,以其为模板,以p35S::GLK质粒为阳性对照,以WT白桦的总DNA为阴性对照,进行BpGLK1基因的PCR扩增,引物分别为:BpGLK-F:5′-ATGCTTGCTGTGTCAGCTTTG-3′;BpGLK-R:5′-AAGCATAAGAGGGTGGTATTTTCC-3′。PCR反应体系及扩增程序参照文献[11],PCR产物用1.0%的琼脂糖凝胶检测。
BpGLK1干扰表达株系的PCR检测:分别提取参试株系的总DNA,以其为模板,以p35S:: GLK-RNAi质粒为阳性对照,以WT白桦的总DNA为阴性对照,采用引物RNAi_Cis-F:5′-CATGCCATGGGCACAGAAGGTTTGTGCAAG -3′;RNAi_Cis-R:5′-TTGGCGCGCCCCATACATCTGCCTTCTCTGG -3′,扩增靶基因的正向序列及部分载体序列,扩增片段长度为414 bp。采用引物RNAi_Anti-F:5′-GCTCTAGAGCACAGAAGGTTTGTGCAAG -3′;RNAi_Anti-R:5′-CGCGGATCCCCATACATCTGCCTTC TCTGG -3′,扩增靶基因的互补序列及部分载体序列,扩增片段长度为429 bp。PCR反应体系及扩增程序参照文献[12],PCR产物用1.0%的琼脂糖凝胶检测。
qRT-PCR检测:分别提取参试株系的总RNA,经DNaseI(RNase free)消化后反转录为cDNA,将其稀释10倍后用作定量PCR的模板,上游引物为:5′-CACAACATAGCCAGCCACCTTC -3′,下游引物为:5′- GTCGGTGCTACCCAAGGACTC -3′。同时以18S rRNA为内参基因,对BpGLK1基因进行qRT-PCR分析,PCR反应体系及扩增程序参照文献[12]。
1.2.2 叶色调查 RHS比色:用RHS标准比色卡(英国皇家园艺学会)与参试株系叶片近轴面进行对比并记录。
色差仪比色:采用分光色差仪(KONICA MINOLTA CR-400)测定参试株系叶片的中部,每个株系测定30个不同单株,每株1个叶片,取其平均值。测定于5月15日开始,9月1日结束,每15 d测定1次,每次每个株系测定3株树叶片。测量结果用CIELAB表色系统进行色度分析[13],其中,L*代表从黑色到白色中间的位置,即明亮度(Lumination),范围是 0~100;a*代表红(+a)绿(-a)色轴饱和度;b*代表黄(+b)蓝(-b)色轴饱和度。
1.2.3 叶片叶绿素含量的测定 采用便携式叶绿素测定仪(SPAD-502 PLUS,KONICA MINOLTA)测定参试11个株系健康的第5叶片的SPAD值,测定于5月15日开始,9月1日结束,每15 d测定1次,每次每个株系测定30株树叶片。
1.2.4 生长性状调查 用米尺测量参试株系苗高,于5月15日开始,9月1日结束,每15 d调查1次,共调查8次(以1月1日为生长起始日,则5月15日为第136天,即最后调查的日期9月1日为第241天)。
1.2.5 数据处理 利用MATLAB Compiler Runtime 8.3对苗高的生长规律进行Logistic方程拟合[13]。
2 结果与分析
2.1 转BpGLK1基因白桦的分子检测
分别以BpGLK1过表达株系叶片的总DNA为模板,用BpGLK1基因ORF两端序列为引物进行PCR扩增,结果显示:阳性质粒及3个过表达株系在1200 bp处扩增出单一谱带,与BpGLK1基因序列1284 bp长度吻合;而WT株系未见扩增谱带(图1)。
分别以BpGLK1干扰表达株系叶片的总DNA为模板, PCR 扩增BpGLK1靶基因的正向序列及互补序列,结果显示:7个干扰表达株系插入的靶基因正向序列及反向互补序列均有扩增谱带,且分别与预期的414 bp及429 bp长度吻合(图2),PCR 扩增结果证明,目标基因仍然整合于2年生的转基因株系基因组中。
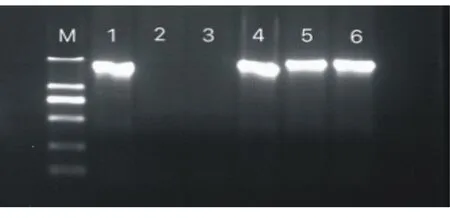
M.DL2000(2 kb、1 kb、0.75 kb、0.50 kb、0.20 kb、0.10 kb);1.阳性质粒;2.WT;3.去离子水;4~6.OE1、OE2、OE3株系 。

a.引物为RNAi_Cis-F、RNAi_Cis-R;1.阳性质粒;2.WT;3.去离子水;4~10.干扰表达株系。b.引物为RNAi_Anti-F、RNAi_Anti-R;1.阳性质粒;2.WT;3. 去离子水;4~10.干扰表达株系。M.DL2000(2 kb、1 kb、0.75 kb、0.50 kb、0.20 kb、0.10 kb)。
分别对参试株系进行qRT-PCR检测,结果显示:与WT株系比较,BpGLK1表达量在3个过表达株系中均呈现上调表达,差异达到显著水平(P<0.01),其中,OE1株系的BpGLK1相对表达量最高;BpGLK1在7个干扰表达株系中均显著下调表达,其中RE5株系的BpGLK1相对表达量最低,较WT株系低77.96%,而RE4株系的BpGLK1下调幅度最小,仅较WT株系低36.09%(图3)。
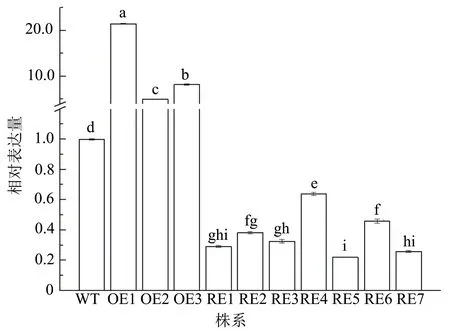
图3 转基因白桦BpGLK1基因的相对表达量
2.2 转BpGLK1白桦叶色变异
RHS比色卡测定结果显示,BpGLK1过表达株系(OE)在5月15日至9月1日植物生长发育季节,叶色以中度橄榄绿或灰色橄榄绿为主;而BpGLK1干扰表达株系(RE4例外)叶色在浓黄绿色和淡黄绿色之间变化;野生型白桦(WT)在6月前期叶色为中度橄榄绿,此后叶色呈现中度黄绿色(图4)。
在CIELAB颜色系统中,L*值是衡量叶色明暗程度的指标,该值越大表明叶片亮度越高;而黄蓝属性b*值由小变大,则表示蓝色的减退、黄色的增加。采用分光色差仪测定叶片,结果显示,在生长季的不同发育时期叶片L*值及b*值在株系间均差异显著(P<0.05),多数干扰表达株系(RE4例外)的L*值显著高于过表达株系及WT株系,其中RE1、RE7株系的L*值在各时期均居前2位(表1);同样,叶片的b*值也呈现干扰表达株系显著高于过表达株系及WT株系(RE4例外)(表2),其中RE1、RE7株系的b*值在各时期均显著高于其他株系。从L*值及b*值的时序变化发现,各株系叶片亮度(L*值)及黄色程度(b*值)在7月1日达到高峰。总之,BpGLK1干扰表达株系由于BpGLK1基因的低量表达,能够不同程度地提高叶色亮度及黄色程度。

a:BpGLK1过表达株系叶色比较。WT.中度黄绿(RHS 138A);OE.灰橄榄绿(RHS NN137C)。b:BpGLK1干扰表达株系叶色比较:WT.中度黄绿(RHS 138A);RE1、RE2、RE5、RE6、RE7浓黄绿(RHS 143C);RE3浓黄绿(RHS 143B);RE4中度黄绿(RHS 138A)。
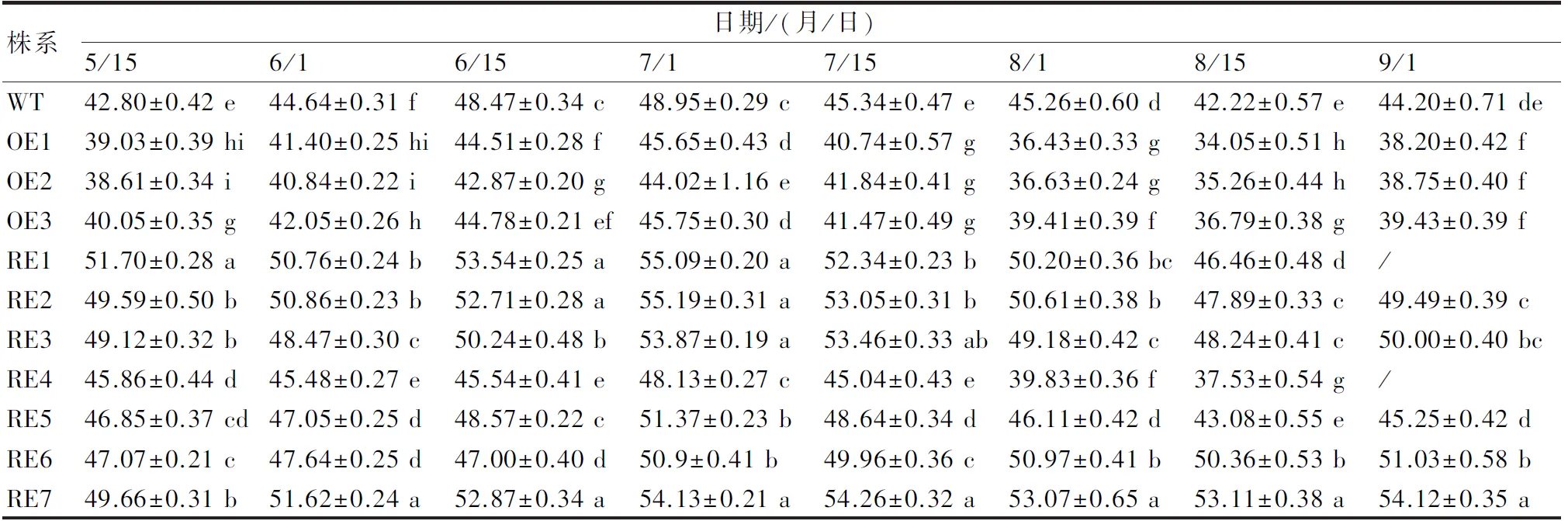
表1 不同发育时期各参试株系叶片L*值多重比较
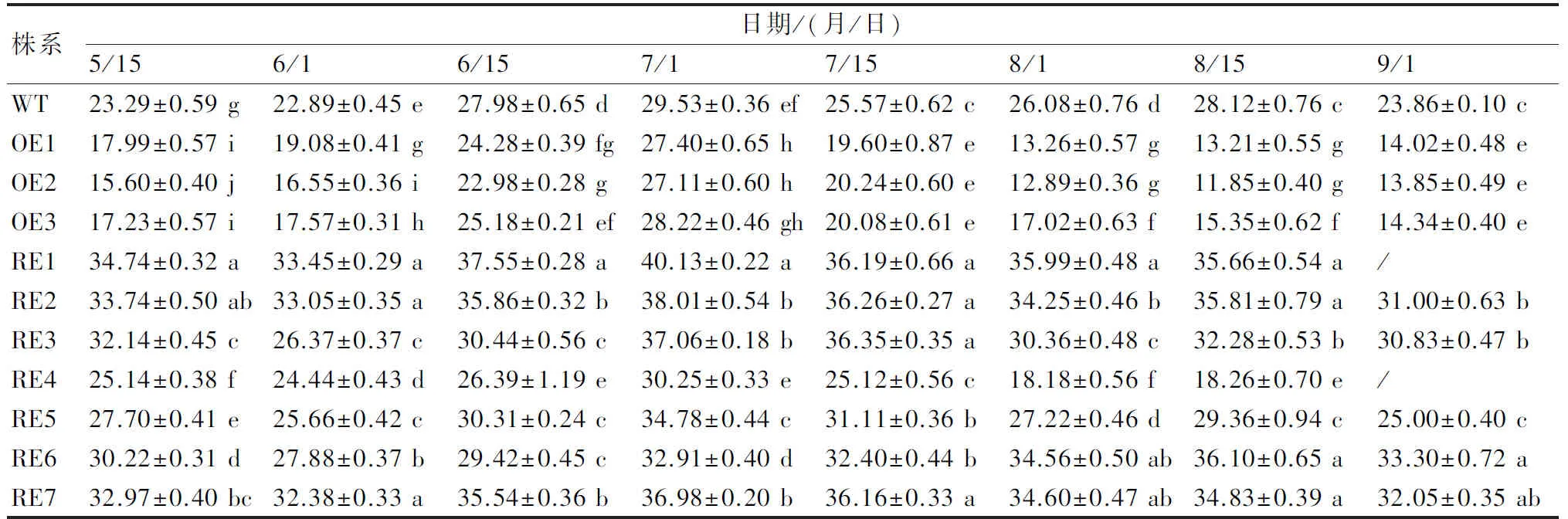
表2 不同发育时期各参试株系叶片b*值多重比较
2.3 转BpGLK1基因白桦叶绿素相对含量的时序变化
转基因株系叶绿素相对含量(SPAD值)测定显示,各株系在生长发育的不同时期叶片叶绿素SPAD值差异显著(P<0.05),在各时期均以BpGLK1过表达株系叶绿素相对含量最高,而BpGLK1干扰表达株系叶绿素相对含量较低(图5)。各株系SPAD值的时序变化趋势表现为7月1日最低,随后3个过表达株系显著高于WT株系,而干扰表达株系(RE4例外)均低于或显著低于WT株系。
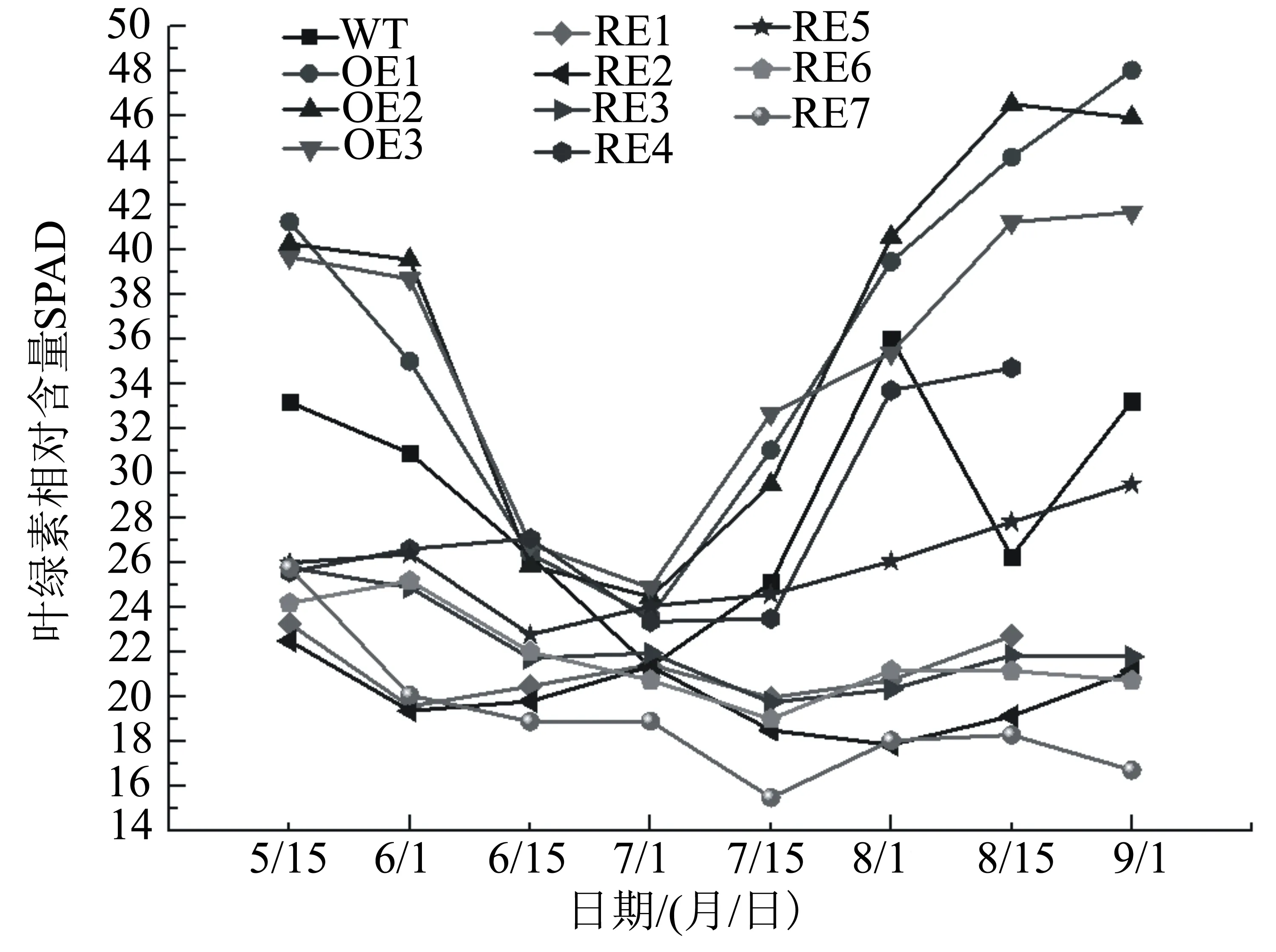
图5 参试株系叶片SPAD值时序变化
2.4 转BpGLK1基因白桦生长变异分析
2.4.1 苗高生长模型建立与拟合 用Logistic模型对11个株系共233个单株苗高生长进行拟合,其生长曲线的拟合度R2均在0.98以上,说明用该模型能够准确模拟参试株系生长曲线,可对参试株系的苗高生长进行分析与预测。
由表3可知,BpGLK1抑制表达株系和过表达株系的苗高(即停止生长后的实测值)均显著高于WT株系(RE1、RE6例外),其中OE3株系最高,为187.29 cm;其次为RE5株系,为167.13 cm;RE4株系的苗高峰值出现最早,t0= 157.46 d,较WT株系及其他参试株系(RE2例外)提前10 d左右;苗高速生点处的生长速度较高为RE1、RE4及OE2,平均为2.15 cm/d,显著高于WT株系及RE3、RE6、RE7,但与其他株系差异不显著(表3)。
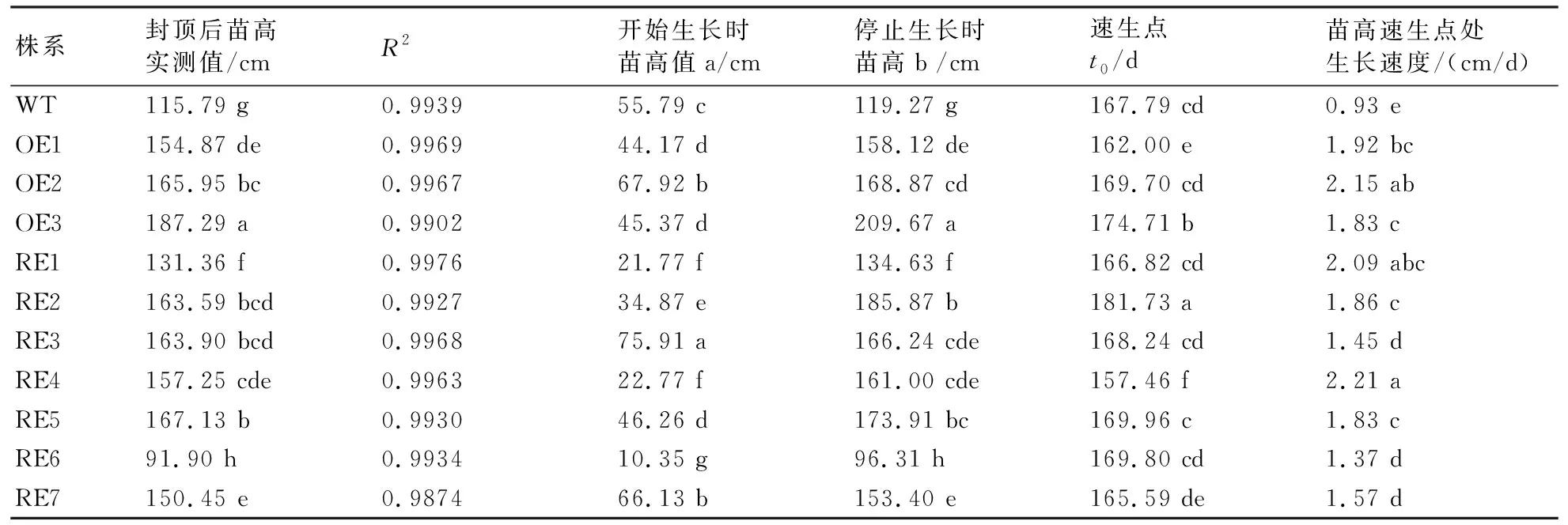
表3 参试株系苗高的Logistic模型
2.4.2 转BpGLK1基因白桦速生期生长参数变异 由表4可知,苗木速生期始期最早的是RE4株系(t1=136.33 d),该株系的速生期结束期也最早。速生期持续时间较长的株系为OE3和RE2,分别为60.88、58.50 d;在速生期内苗高平均生长量(GR)高于群体均值的株系为OE1、OE2、OE3、RE1、RE2、RE4和RE5,这些株系在速生期内苗高日生长量(GD)在1.5 cm/d以上,其中RE4最高,苗高日生长量达1.9330 cm/d(表4)。参试株系在速生期内的生长量占整个生物量的56%~58%,说明速生期内苗高生长在其整个生长期中起重要作用。

表4 参试株系速生期生长参数比较
3 结论与讨论
GLK转录因子在拟南芥、水稻(OryzasativaL.)、辣椒和番茄等植物中研究证明,GLK基因家族成员通过协调光合器官的表达从而调控植物叶绿体发育,对叶绿体发育至关重要,拟南芥AtGLK1和AtGLK2双突变体叶片表现为叶绿素含量降低,呈现黄色[4];在番茄中SIGLK1抑制表达株系叶色呈现黄绿色,其叶绿素含量较对照株系降低[4]。本研究团队获得的白桦转BpGLK1基因研究也证实了前人的研究结果,转基因白桦移栽当年叶色及光合色素测定发现,与野生型白桦比较,BpGLK1过表达株系表现为叶片绿色加深,叶绿素含量略高;而BpGLK1抑制表达株系(RE4例外)则叶色呈现黄绿色,叶绿素含量减少[14]。本试验是在团队研究基础上,对移栽2年的转BpGLK1白桦追踪调查,PCR及qRT-PCR检测表明,目标基因仍稳定整合于转基因白桦基因组中,并在mRNA水平上正常表达(图3)。叶色及叶绿素相对含量分析发现,与苗期呈现的规律一致,BpGLK1过表达株系的叶绿素含量较高,则叶片绿色程度也高,反之,BpGLK1干扰表达株系(RE4例外)的叶绿素含量较低,则叶色为黄绿色(图4)。通过连续2年的观测发现,RE4株系虽然是BpGLK1干扰表达株系,但其叶片叶绿素相对含量与WT差异不显著,叶色仍然为绿色,BpGLK1基因在叶片中的相对表达量下调程度较低。连续2年的qRT-PCR检测显示,该株系的BpGLK1的相对表达量在1年生时较WT株系低41.00%,在2年生时较WT株系低36.09%,而其他6个干扰表达株系BpGLK1的表达量均较WT株系低54.16%以上(图3)。众所周知,转基因植物外源基因的插入通常是随机的,T-DNA插入位点不同,可能导致目标基因表达程度不尽相同,我们推测由于RE4株系BpGLK1基因表达下调幅度小于其他转基因株系,因此,其叶绿素相对含量与WT差异不显著,叶色呈现绿色。后续将通过T-DNA插入位点的鉴定及侧翼序列分析探究影响RE4株系叶色的因素。
对于多年生的木本植物,在获得转基因株系后往往需要经过多年的选择,只有既表现出导入目标基因性状,其生长及产量又不受影响的转基因株系才能成为应用推广的转基因新品种[15,16]。对2年生转BpGLK1白桦干扰表达株系叶色及生长分析,初步选择RE1、 RE7株系为优良株系,这2个株系在叶片亮度及黄色程度方面均优于其他株系,其中RE7株系已获批植物新品种权(20190378),可作为城乡绿化彩叶树种,丰富东北地区的园林绿化树种组成。
研究发现,植物高叶绿素含量并不是高光合速率的必需条件。高叶绿素含量会加剧植物个体之间竞争,不利于群体的光合作用[17,18]。例如,水稻低叶绿素突变体研究发现,在高光强条件下,适当降低叶绿素含量,有利于缓解因过量光吸收而导致的活性氧的产生以及对光系统的破坏,缓解光抑制,提高光系统Ⅱ光电转化效率与光合电子传递效率,进而可以提高水稻的光合能力[19]。此外,有研究表明,番茄叶色黄化突变体与正常植株在生长势上没有显著差异,且果实产量与普通品种差异不显著,突变体果实内色素含量低于正常品种,耐贮性和硬度都优于正常品种[20,21]。陆地棉(GossypiumhirsutumL.)黄绿苗突变体浙12-12N幼苗期真叶叶绿素含量低于常绿苗泗棉2号, 其各叶位叶片的水分利用率、光能利用率及净光合速率却高于泗棉2号,子棉产量和皮棉产量也要高于常绿苗泗棉2号[22]。上述研究证明,适当地降低叶绿素含量,可以增强植物的光合能力,提高作物产量及品质。
转BpGLK1白桦苗高生长调查显示,移栽当年苗高生长与WT株系差异不显著[14];而在2年生时,无论是BpGLK1过表达株系,还是干扰表达株系,其苗高生长均显著高于WT株系(RE1、RE6例外),在速生期内苗高平均生长量(GR)高于群体均值的株系为OE1、OE3、RE1、RE2、RE4和RE5,其中RE4苗高日生长量高达1.9330 cm/d。试验表明,BpGLK1干扰表达株系的叶片叶绿素含量降低,并未影响干扰表达株系的高生长。白桦与上述作物同样,适当降低叶片叶绿素含量可有效促进白桦苗期株高生长。白桦为多年生高大乔木,我们后续还需追踪调查其生长特性,探讨转BpGLK1基因白桦的吸收光能分配特性、叶绿体荧光参数及光合作用速率等变化。
猜你喜欢
种子(2022年6期)2022-08-03
农业灾害研究(2022年2期)2022-05-31
南方农业·下旬(2022年4期)2022-05-24
语文世界(小学版)(2021年5期)2021-06-04
中国蔬菜(2021年3期)2021-03-29
上海师范大学学报·自然科学版(2020年5期)2020-12-18
林业与生态(2019年6期)2019-07-02
南方农业学报(2018年9期)2018-09-10
山东农业科学(2018年3期)2018-03-29
山东农业科学(2018年11期)2018-01-17
