
川西亚高山森林木质残体及其附生苔藓持水特性
2021-10-09 01:52王芝慧黎静妤侯建峰蒋雨芮李旭清郑冰倩杨万勤
生态学报 2021年16期
王芝慧,黎静妤,白 义,李 飞,侯建峰,蒋雨芮,2,李旭清,郑冰倩,杨万勤,*
1 台州学院生命科学学院,台州 318000 2 长江上游林业生态工程省级重点实验室,四川农业大学生态林业研究所,成都 611130 3 王朗国家级自然保护区,绵阳 622550
木质残体是绝大多数森林生态系统的基本结构[1]。其中,粗木质残体储量约占全球木质生物量的20%—30%[2],在水源涵养、水土保持、生物多样性保育、森林更新与演替、物质循环等方面发挥着重要作用[3-4]。木质残体可通过多种途径影响森林水文生态过程[5]。首先,不同分解阶段的木质残体通过对降水的截留,影响土壤入渗和林地蒸发等森林水文生态过程[6]。其次,木质残体能够增加地表糙率,改变林内微地形,影响森林流域的产流产沙[7]。然而,已有的森林水文生态研究多关注乔木层、林下苔藓层、枯落物层以及土壤层对降雨的截留作用[8- 10],有关木质残体在森林水文生态过程中的作用尚不清楚,这不能满足森林生态管理的需要。
不同分解阶段和不同径级大小的木质残体是苔藓等附生植物的良好基质[11-12],苔藓自身疏松多孔及较大的表面积特性,与海绵的弹力性状相似,使其持水率比森林地面上其他植被类别的持水率高两至五倍左右[13-14]。因此,附生苔藓对森林水文过程和物质循环具有潜在的重要影响,也是一个评判森林生态系统完整性的潜在指标[15]。森林苔藓主要来源两个方面:林下苔藓层和木质残体附生苔藓。首先,石生、树生或木质残体附生等不同来源的苔藓[16],其持水性能存在显著差异;其次,不同的苔藓种类更是影响苔藓持水能力的主要原因之一[17];此外海拔高度[18]、生态恢复模式[14]也能显著影响苔藓的水文效应。但是理论上,苔藓的储量和持水能力还受到林型、腐烂等级和径级大小等的影响。然而,已有的研究主要集中在森林地表林下苔藓层的持水能力研究[13- 20],有关木质残体附生苔藓水源涵养能力尚缺乏关注。
川西亚高山针叶林是我国第二大林区(西南林区)的主体,在水源涵养、水土保持和生物多样性保育等方面具有不可替代的战略地位[21]。不同腐烂等级的木质残体及其附生苔藓是亚高山针叶林生态系统的两个基本结构[22-23],其可能对森林水文生态过程产生不同程度的影响。然而,有关亚高山针叶林木质残体及其附生苔藓的持水特性随林型、腐烂等级和径级大小的变化特征尚缺乏必要的关注。因此,基于前期调查,以川西亚高山岷江冷杉原始林、紫果云杉原始林、方枝柏原始林等8个典型森林类型研究对象,调查研究了木质残体及其附生苔藓的饱和储水量和持水率随林型、腐烂等级和径级的变化特征,以期为深入理解亚高山森林水文生态过程和可持续经营管理提供科学依据。
1 材料和方法
1.1 研究区概况
本研究位于四川省平武县王朗国家级自然保护区(32°49′N—33°02′N,103°55′E—104°10′E,a.s.l. 2300—4983 m),属于丹巴—松潘半湿润气候,年平均气温2.9℃,7月平均气温12.7℃,1月平均气温-6.1℃,极端高温26.2℃,极端低温-17.8℃;年降水量859.9 mm,降水日数195 d,受季风影响,降雨主要集中在生长季5月—9月。如表1所示,本项研究涵盖了8种典型亚高山寒温性落叶阔叶林和寒温性常绿针叶林。森林乔木层主要有岷江冷杉(Abiesfaxoniana)、紫果云杉(Piceapurpurea)、方枝柏(Sabinasaltuaria)、糙皮桦(Betulautilis)、白桦(B.platyphylla)等。高灌树主要有皂柳(Salixwallichiana)、沙棘(Hippophaerhamnoides)、榆树(Ulmuspumila)等。林下灌木主要有高山杜鹃(Rhododendronlapponicum)、峨眉蔷薇(Rosaomeiensis)、长叶毛花忍冬(Loniceratrichosanthavar.xerocalyx)、石灰花楸(Sorbusfolgneri)、川滇小檗(Berberisjamesiana)、紫花卫矛(Euonymusporphyreus)等。草本植物主要有掌裂蟹甲草(Cacaliapalmatisecta)、莎草科(Cyperaceae)、草地早熟禾(Poapratensis)等。苔藓植物主要有异叶提灯藓(Mniumheterophyllum)、塔藓(Hylocomiumsplendens)、大羽藓(Thuidiumcymbifolium)、尖叶匍灯藓(Plagiomniumacutum)、美姿羽苔(Plagiochilapulcherrima)、偏叶管口苔(Solenostomacomatum)、中华耳叶苔(Frullaniasinensis)、黄角苔(Phaeoceroslaevis)、石地钱(Rebouliahemisphaerica)等。
1.2 研究方法
1.2.1野外调查与采样
2019年8月初(雨季),在前期对王朗国家自然保护区亚高山针叶林区进行踏查的基础上,以8种典型的亚高山森林类型为研究对象(表1),分别在8种典型林型设置3个大小为10 m×20 m的样方,总计共24个调查样方,开展木质残体及其附生苔藓持水能力调查研究。其中,木质残体腐烂等级划分参照Pyle和Brown[24]的方法分为5个等级(表2);径级大小划分根据Harmon等[1]的分类标准和Sefidi等[25]的分类方法,再根据本研究的具体目标,将木质残体根据直径划分为5个径级:2 cm≤d≤5 cm(D1)、5 cm
调查方法:首先,对每个样方内所有的木质残体(直径≥2 cm,长度或者高度>20 cm)的倒木、枯立木、树桩、大枯枝和小枯枝进行逐一测量和调查,针对完整的倒木,测量大头直径和长度;对于不完整的树桩和枯立木,测量两头直径和高度,所有指标均用卷尺测量3次,最后根据这些数据计算木质残体体积和表面积,再根据样方面积计算储量等指标。对木质残体的腐烂等级根据表2的标准判断并记录;对木质残体的树皮脱落、苔藓等附生植物分布情况进行记录。其次,根据王朗植物的记录识别附生苔藓的种类,根据测量和观察估计覆盖度。然后,采集不同类型木质残体,带回实验室测定含水量和密度等。最后,采集不同腐烂等级、不同径级大小和不同种类的木质残体表面的附生苔藓,带回实验室测定附生苔藓生物量和饱和储水量等。
采样方法:在每个样方中,每份相同的样品采集3个重复。对腐烂等级I到III级的粗木质残体,使用油锯截取大约5 cm左右厚的圆盘;对腐烂等级IV到V级的粗木质残体和不同腐烂等级的细木质残体,每个称取500 g左右的样品;对于不同树种和不同腐烂等级的木质残体上的附生苔藓,使用小刀割取20 cm×20 cm大小的苔藓样品,均标记分装好带回实验室做后续分析。
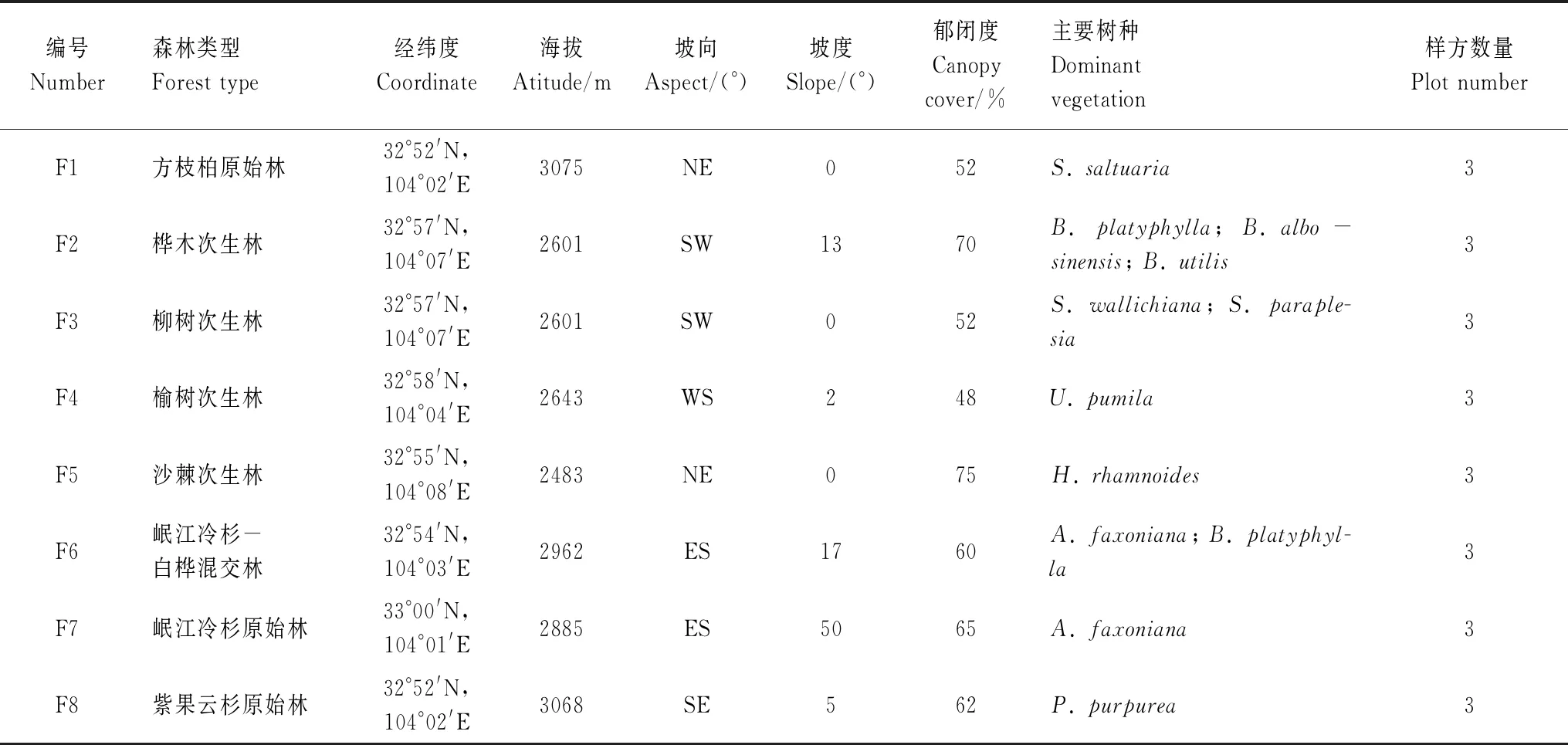
表1 典型亚高山森林样地概况

表2 木质残体腐烂等级分类
1.2.2木质残体及其附生苔藓持水特性测定
I到III级粗木质残体圆盘样品带回实验室后,测量其直径d1和厚度h,进一步计算其体积V;IV到V级的粗木质残体和细木质残体样品带回实验室后,直接采用排水法测定其体积V,然后将木质残体样品置于85℃干燥箱中烘干至恒量m1,体积与干重的比值即为密度ρ[26]。木质残体饱和储水量采用浸泡法测定:将烘干后的圆盘或细木质残体样品完全浸入水中过夜吸满水,拎起不再滴水后称重m2。附生苔藓样品带回实验室后,在85℃干燥箱中烘干至恒量m3。然后,附生苔藓的饱和储水量同样采用浸泡法测定:将烘干后的苔藓样品完全浸入水中过夜吸饱水,拎起后不再滴水,称其吸饱水后的重量m4。对于野外调查的完整的倒木按圆锥体计算体积和表面积,针对不完整的树桩和枯立木,按台体计算体积和表面积[27],木质残体的饱和储水量和饱和持水率根据刘一霖等[28]计算。
相关计算公式为:
RWD=(m2-m1)/m1×100%
WWD=RWD×GWD/10(mm)
RM=(m4-m3)/m3×100%
WM=RM×GM/10(mm)
式中,RWD和WWD分别为野外调查的木质残体的饱和持水率(%)和饱和储水量(mm);RM和WM分别为野外调查的木质残体附生苔藓的饱和持水率(%)和饱和储水量(mm);GWD为野外调查的木质残体的储量(t/hm2);GM为野外调查的木质残体附生苔藓的储量(t/hm2);m1和m2分别为采集的木质残体烘干状态质量和吸饱水状态质量(g);m3和m4分别为附生苔藓烘干状态质量和吸饱水状态质量(g);10为将饱和持水量单位t/hm2换算成mm的系数。
其中,野外调查的木质残体及其附生苔藓的储量计算公式为:
GWD=∑V×ρ/S0(t/hm2)
GM=∑m3×S×C/0.04S0(t/hm2)
式中,S0为每个样方的正投射面积(m2);V为野外调查的木质残体的体积(m3);ρ为根据采集的木质残体样品计算的密度(g/cm3);m3为附生苔藓烘干状态质量(g);S分别为野外调查的木质残体的侧面积(m2);C为野外调查的木质残体上附生苔藓的覆盖度(%);0.04为割取20 cm×20 cm样品的苔藓的面积(m2)。
其中,野外调查的木质残体的体积、密度和侧面积的计算公式为:
V1=πld22/12(m3)

V2=πld22/12+πld2d3/12+πld32/12(m3)

ρ=m1/V(g/m3)
式中,V1和S1分别为野外调查的完整倒木按圆锥体计算的体积和侧面积;V1和S1分别为不完整树桩和枯立木按台体计算的体积和侧面积;d2,d3和l分别为木质残体的两端直径和长度(m);V为采集的木质残体样品的体积;m1为木质残体烘干状态质量(g)。
1.3 数据处理与统计分析
采用单因素方差分析(One-way ANOVA)和Duncan法检验不同林型、不同腐烂等级和不同径级之间木质残体及其附生苔藓各类指标的差异显著性;采用多因素方差分析Multivariate analysis of variance(MANOVA)和效应量估计分析林型、腐烂等级和径级大小对木质残体及其附生苔藓各类指标的交互作用并分析影响的主要因素;采用Pearson相关性分析方法分析不同林型、不同腐烂等级和不同径级之间木质残体或附生苔藓的储量与饱和储水量之间的相关性;为了检测饱和储水量或饱和持水率与腐烂等级之间的响应趋势,采用二次项方程Q=ex2+fx+g拟合它们之间的回归关系;采用SigmaPlot12.5软件进行作图;采用Excel 2019和SPSS 20.0对数据进行统计分析处理。
2 结果与分析
2.1 木质残体储水量和持水性能随林型的变化
图1显示,8种亚高山森林类型的木质残体饱和储水量介于4.55—22.06 mm之间,不同林型之间差异显著(P<0.01)。研究还发现,木质残体的饱和储水量与其储量呈现显著正相关(r=0.938,P<0.01)(表3和表4),木质残体储量较高的原始林紫果云杉林和岷江冷杉林具有最大的饱和储水量,分别为22.06 mm和17.24 mm;但木质残体储量较低的次生林柳树林、沙棘林、榆树林和桦木林具有较低的木质残体饱和储水量,分别为4.55、5.55、5.79、6.62 mm;岷江冷杉-白桦混交林的饱和储水量水平中等,有11.25 mm;而针叶原始林方枝柏林,它的饱和储水量与次生林接近,仅有5.09 mm。另外,8种森林的木质残体吸水能力接近,差异不显著(P>0.05),平均能够吸取自身质量的2.28倍的水分,其中紫果云杉原始林和柳树次生林的饱和持水率相对较高,达到291.50%和290.37%,而方枝柏原始林和沙棘次生林的饱和持水率相对较低,为180.02%和176.34%。

图1 亚高山针叶林区木质残体储水能力随林型的变化Fig.1 Changes in water holding capacity of woody debris with forest types in the subalpine forest region图中数据为平均值±标准误差(n=3);图中不同小写字母表示不同森林类型木质残体之间的饱和储水量差异显著(P<0.05)

表3 不同森林类型、腐烂等级或径级大小下的木质残体及其附生苔藓的储量

表4 亚高山典型森林木质残体及其附生苔藓的储量和饱和储水量之间的相关性
2.2 木质残体附生苔藓的储水量和持水性能随林型的变化
木质残体附生苔藓的饱和储水量和持水率在不同林型间显著变化(P<0.001),饱和储水量的变化范围在0.21—7.01 mm之间(图2),饱和储水量与储量之间表现为正相关性(r=0.984,P<0.01)(表3和表4)。其中,附生苔藓储量最高的紫果云杉原始林具有最高的饱和储水量(7.01 mm),显著高于其他林型;岷江冷杉原始林(4.07 mm)和岷江冷杉-白桦混交林(4.48 mm)的木质残体附生苔藓饱和储水量水平中等;次生林沙棘林(2.73 mm)、榆树林(1.35 mm)、柳树林(1.19 mm)和白桦林(1.16 mm)的附生苔藓饱和储水量较低;而附生苔藓储量最低的方枝柏原始林具有最低的饱和储水量(0.21 mm),显著低于其他林型。另外,从图2我们可以看出,附生苔藓饱和持水率较高的森林是原始林紫果云杉林(576.41%),岷江冷杉林(575.61%)和方枝柏林(541.86%),能吸取自身质量将近6倍的水分;榆树次生林的木质残体附生苔藓饱和持水率最低(385.78%),显著低于其他林型(P<0.01)。附生苔藓的持水性能达到自身质量的3.9倍到5.8倍,显著高于木质残体。

图2 亚高山针叶林区木质残体附生苔藓储水能力随林型的变化Fig.2 Changes in water holding capacity of epiphytic moss on woody debris with forest types in the subalpine forest region图中数据为平均值±标准误差(n=3);图中不同小写字母表示不同森林类型木质残体附生苔藓之间的饱和储水量或饱和持水率差异显著(P<0.05);F1:方枝柏原始林;F2:桦木次生林;F3:柳树次生林;F4:榆树次生林;F5:沙棘次生林;F6:岷江冷杉-白桦混交林;F7:岷江冷杉原始林;F8:紫果云杉原始林
2.3 木质残体储水量和持水性能随腐解等级的变化
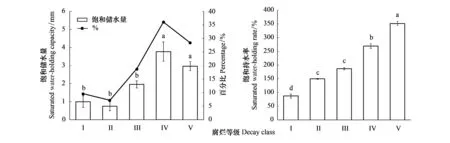
图3 亚高山针叶林区木质残体储水能力随腐烂等级的变化Fig.3 Changes in water holding capacity of woody debris with decay classes in the subalpine forest region图中数据为平均值±标准误差(n=3);图中不同小写字母表示不同腐烂等级木质残体之间的饱和储水量或饱和持水率差异显著(P<0.05)
如图3所示,5个腐烂等级木质残体的饱和储水量介于0.75—3.77 mm之间,不同腐烂等级之间差异显著(P<0.01),与其储量之间表现为正相关性(r=0.698,P<0.01)(表3和表4)。其中腐烂等级IV(3.77 mm)和V(2.97 mm)的木质残体的饱和储水量显著高于I(1.00 mm)和II(0.75 mm)腐烂等级。5个腐烂等级的木质残体对川西亚高山生态系统持水量的贡献为:IV(36.09%)>V(28.41%)>III(18.74%)>I(9.58%)>II(7.19%)。另外,腐烂程度还显著影响木质残体的饱和持水率(P<0.001),其饱和持水率随着腐烂等级的升高而显著增加,总体表现为I(86.81%) 图4 木质残体饱和持水率和附生苔藓饱和储水量与腐烂等级的回归分析Fig.4 The regression analysis between saturated water-holding rate of woody debris or saturated water-holding capacity of epiphytic moss and decay classes图中R2表示饱和持水率或饱和持水量随腐烂等级变化的测定系数,P<0.001表示此回归方程极其显著 图5 亚高山针叶林区木质残体附生苔藓储水能力随腐烂等级的变化Fig.5 Changes in water holding capacity of epiphytic moss on woody debris with decay classes in the subalpine forest region图中数据为平均值±标准误差(n=3);图中不同小写字母表示不同腐烂等级木质残体附生苔藓之间的饱和储水量差异显著(P<0.05) 腐解程度显著影响木质残体附生苔藓的饱和储水量,腐烂等级越高,其饱和储水量越大,具有显著正相关性(r=0.882,P<0.001),变化范围介于0.09—1.49 mm之间(图5),同时,附生苔藓的饱和储水量与其储量之间也表现显著正相关(r=0.997,P<0.01)(表3和表4)。不同腐烂等级的木质残体附生苔藓对川西亚高山生态系统持水量的贡献为:V(50.26%)>IV(27.73%)>III(12.85%)>II(6.23%)>I(2.93%)。研究还发现,其饱和储水量(Q′)与腐烂等级(x)之间的关系也符合二次方程Q′=ex2+fx+g的函数关系式(图4)。然而,进一步研究发现,不同腐烂等级附生苔藓之间的饱和持水率没有显著差异(P>0.05),其平均持水能力为539.72%,即能吸取自身质量5.40倍的水分。 如图6所示,不同径级大小的木质残体饱和储水量变化表现是:径级越大,木质残体饱和储水量越高,具有显著差异(P<0.001)。其变化范围介于0.09—8.31 mm之间,与各径级大小木质残体的储量显著正相关(r=0.991,P<0.01)(表3和表4)。储量最大的D5径级木质残体具有最高饱和储水量(8.31 mm),占比超过60%,显著高于其他径级;而储量最少的D1径级木质残体具有最低的饱和储水量(0.09 mm),显著低于其他径级。不同径级大小的木质残体对川西亚高山生态系统储水量的贡献为:D5(63.86%)>D4(17.48%)>D3(15.62%)>D2(2.31%)>D1(0.73%)。同时研究还发现,不同径级大小的木质残体饱和持水率之间也变化显著(P<0.05),D5径级木质残体的饱和持水率最高,为247.25%,D1径级木质残体的饱和持水率最低,为171.57%,D5径级显著高于D1径级。 图6 亚高山针叶林区木质残体储水能力随径级的变化Fig.6 Changes in water holding capacity of woody debris with diameter sizes in the subalpine forest region图中数据为平均值±标准误差(n=3);图中不同小写字母表示不同腐烂等级木质残体附生苔藓之间的饱和储水量或饱和持水率差异显著(P<0.05) 图7 亚高山针叶林区木质残体附生苔藓储水能力随径级的变化Fig.7 Changes in water holding capacity of woody debris epiphytic moss with diameter sizes in the subalpine forest region图中数据为平均值±标准误差(n=3);图中不同小写字母表示不同腐烂等级木质残体附生苔藓之间的饱和储水量差异显著(P<0.05) 不同径级大小的木质残体附生苔藓的饱和储水量差异显著(P<0.01),变化范围介于0.08—1.69 mm之间,与各径级大小木质残体附生苔藓的储量显著正相关(r=0.993,P<0.01)(表3和表4)。具体表现为D5径级的木质残体附生苔藓的饱和储水量最高,占比超过40%,显著高于其他径级大小的饱和储水量(图7)。其次,不同径级大小的木质残体附生苔藓对川西亚高山生态系统储水量的贡献为:D5(46.47%)>D3(24.35%)>D4(21.44%)>D2(5.47%)>D1(2.26%)。进一步研究发现,不同径级与不同腐烂等级下的木质残体附生苔藓饱和持水率表现相似,均没有显著差异(P>0.05)。 多因素方差分析结果表明,林型、腐烂等级和径级大小两两之间和三者之间对木质残体及其附生苔藓的饱和储水量均具有显著的交互影响(P<0.001;表5)。同时,效应量估计结果表明,在3个单因素中,径级大小是影响木质残体饱和储水量的主要因素,而对附生苔藓饱和储水量影响最大的因素则是腐烂等级。然而,林型、腐烂等级和径级大小三者的交互对木质残体及其附生苔藓的影响比两两交互和单个因素的影响要更大(表5)。 表5 不同森林类型、腐烂等级和径级大小对木质残体及其附生苔藓储水能力的多因素方差分析 木质残体和附生苔藓的水文和水土保育功能是森林水文研究的重要一环[13]。林分结构、腐解程度等显著影响着其水源涵养功能[29]。本研究结果表明,川西亚高山针叶林区不同类型森林木质残体及其附生苔藓的持水能力之间表现明显差异。其中,原始林紫果云杉林和岷江冷杉林木质残体的饱和储水量最高,柳树次生林木质残体的饱和储水量最低,主要原因是由于原始林木质残体储量较高,而柳树次生林的储量较低(表3),因为储量特征是影响持水能力最关键因素之一[30]。此外,木质残体的储量又与森林年龄密切相关[31],演替后期森林木质残体的累积要显著高于前期,因此,处于演替前期的柳树次生林的木质残体输入量要低于后期的原始林。最后,持水率也是反映储水量的一个重要指标,而本研究的不同林型之间的饱和持水率并没有显著差异,说明影响川西亚高山针叶林区木质残体饱和储水量的关键因素是其自身的储量和地上生物量的输入。 本研究还发现,方枝柏原始林木质残体附生苔藓的饱和储水量最低。储量的差异是最主要的影响因素,方枝柏林木质残体本身较低的储量决定了低的附生苔藓储量(表3)。其次,附生苔藓的覆盖度是影响其饱和储水量的另一个原因[32],而影响覆盖度的因素主要有温湿度变化和林分结构。其中温湿度会显著影响苔藓等附生植物的生长[33],因为林地温湿度的改变会影响光合作用的生产[34]和苔藓种群的结构;其次,林分结构中的郁闭度也会严重影响苔藓的生长和分布[13, 33]。方枝柏林内较低的郁闭度导致了充足的光照和较大的蒸发量,进而使得林内苔藓较难生长,而干旱的雏形土土壤特质和干燥的林分微气候导致木质残体上苔藓附生较少,进一步造成其储水能力较低。 不同地区各林分木质残体及其附生苔藓(或林下苔藓)间水源涵养能力也有差异(表6)。本研究的川西亚高山针叶林区森林类型跨度较大,其木质残体饱和储水量范围在50.88—220.62 t/hm2之间,同为川西亚高山的毕棚沟、西藏色季拉山和美国的草原河红杉州立公园的结果在本研究范围之内,但本研究结果高于天宝岩各类混交林和大兴安岭火烧迹地木质残体储水量,又显著低于白龙江高海拔的杜鹃纯林木质残体储水能力(表6)。地理位置是影响持水性能的重要因素[35],这些地区从南到北,跨越多个经纬度的空间距离,因此会导致气候的显著变化,其中最为明显的就是降雨量、温湿度的差异,进而导致不同水热条件下森林小气候和郁闭度等也存在显著差异;其次,海拔也是主要的影响因素之一[36],海拔越高的地区气候条件相对恶劣,这减缓了木质残体的腐解速率,导致高海拔地区木质残体储量越大,其持水能力也越强。然而,相对于饱和持水量而言,本研究的木质残体饱和持水率范围在176.34%—291.50%之间,与相同地区和其他地区如色季拉山、天宝岩、白龙江和大兴安岭相差不大,分别为184.98%、168.82%、126.59%—211.57%、199.92%和172.34%—195.25%(表6)。 表6 不同地区森林类型木质残体或苔藓的持水能力 本研究中,各林型木质残体附生苔藓的饱和储水量范围在2.15—70.09 t/hm2,饱和持水率范围在385.78%—576.41%之间,其中,饱和储水量与川西亚高山米亚罗林区、西藏色季拉山和长白山的林下苔藓层饱和储水量相近,但显著低于川西亚高山毕棚沟原始冷杉林的林下苔藓饱和储水量(表6);而饱和持水率与川西亚高山毕棚沟、米亚罗林区、西藏色季拉山和长白山的林下苔藓层饱和持水率相差不多,只有米亚罗林区的原始冷杉林林下苔藓饱和持水率超高,达到1195.85%。苔藓本身的种类是重要的影响因子[17],因为不同种类的苔藓形态特征和生理生态特征不同。刘艳等[17]研究表明桧叶白发藓(Leucobryumjuniperoideum),由于植物个体间存在大量毛孔间隙,并且鞘状的叶片基部增加了表面积,进而有助于吸收更多水分,而叶片有类似角质层结构的东亚小金发藓(Pogonatuminflexum)则较难吸收水分,因此持水能力较差。但即使苔藓种类相同,其饱和储水量会存在较大的差异,因为还基于单位面积上苔藓的生物量,所以木质残体本身的储量也是附生苔藓饱和储水量的一大限制因子。此外,生长基质[16]、海拔[18]、林龄[44]等可能都是影响苔藓持水性能的几个关键因素。因此,保护原始冷杉林和云杉林,促进次生林更新,增加森林木质残体及其附生苔藓储量对于水源涵养等具有重要作用。 分解程度是影响木质残体及其附生苔藓持水能力的关键因子[29],本研究中,较高腐烂等级(IV和V)的木质残体的饱和储水量要显著高于低腐烂等级(I、II和III),此结果与色季拉山长孢冷杉林一致[38]。主要原因是分解程度越高,饱和持水率越高。因为腐解程度越高的木质残体密度越小,结构疏松,孔隙度更大,水分能快速渗透进木材的组织和细胞中,因此其持水能力也越高,并且高腐烂的木质残体表面会滋生更多的附生苔藓,而苔藓反过来可增加水汽在木质残体上凝结;另外,本研究的保护区内的高度腐烂等级的木质残体本身储量较高也是一个原因。本研究木质残体饱和持水率随着腐解等级升高而增强,符合Q=ex2+fx+g多项式方程的函数关系式,与目前报道的腐烂等级与持水率的关系均一致[35, 38, 45]。此外,木质残体的腐烂等级越高,其附生苔藓饱和储水量越大,它们的函数关系也符合Q′=ex2+fx+g多项式方程式。首先,高腐解程度的木质残体维管组织结构疏松,微生物种类丰富,较低腐烂等级为苔藓等植物提供了更丰富的营养物质和生长基质[46],因此覆盖的苔藓植物也越多。其次,附生苔藓饱和储水量还基于其单位面积的生物量大小,因为本研究高腐烂等级的木质残体本身蓄积量更高,因而其附生苔藓储水量也较高。因此,保留较高腐烂等级的木质残体及其附生苔藓,对于维持亚高山针叶林水源涵养功能具有重要意义。 本研究中,木质残体及其附生苔藓的饱和储水量均随着径级的增大而升高,超过60%和40%的水量都保存在D5(d>40 cm)大径级木质残体和附生苔藓上。首先,最主要的原因是大径级木质残体及其附生苔藓的储量更高,其次可能与木质残体比表面积有关。已有研究表明,径级大小与比表面积呈现显著负相关关系[37],而比表面积与饱和储水量也是负相关,因此,大径级木质残体具有更高的饱和储水量和饱和持水率[41]。而有相关研究表明,土壤储水量与比表面积是显著正相关性[47],这可能由于土壤和木质残体(附生苔藓)特性不同导致的。因此,在森林经营与管理实践中,保留大径级的倒木等木质残体对于维持森林生态系统结构和功能具有重要意义。 木质残体由于本身的吸水特性以及对降水的截流、增加地表粗糙率等功能[6-7],其附生苔藓由于海绵质地赋予的强大的吸水能力[13],成为了森林水文生态研究中重要的组成。以往研究多关注木质残体本身的水源涵养功能,但往往忽略具有强大吸水能力的附生苔藓。本文研究表明川西亚高山原始针叶林内高腐烂等级和大径级的木质残体及其附生苔藓具有强大的持水性能和蓄水潜能,说明其水源涵养能力在整个川西亚高山森林水文生态系统中占据重要的位置,为森林生态健康持续稳定发展提供基础。同时,林型、腐烂等级和径级大小三者的相互作用对其储水能力的影响显著高于单个因素,说明在森林经营与管理中,同时保留原始林、高腐烂等级和大径级的木质残体及其附生苔藓对于维持森林水文生态稳定性具有重要意义。不过,本研究对于木质残体及其附生苔藓的持水能力的动态变化如吸水速率等还缺乏持续关注。因此,想要更全面了解亚高山森林木质残体及其附生苔藓的水土保育功能,其水源涵养能力随时间、季节、海拔、土壤等的动态变化,还需要进一步的调查研究。
2.4 木质残体附生苔藓储水量和持水性能随腐烂等级的变化

2.5 木质残体储水量和持水性能随径级的变化


2.6 木质残体附生苔藓储水量和持水性能随径级的变化

3 讨论
3.1 林型对木质残体及其附生苔藓持水能力的影响

3.2 腐烂等级对木质残体及其附生苔藓持水能力的影响
3.3 径级大小对木质残体及其附生苔藓持水能力的影响
4 结论
猜你喜欢
南方农业(2022年16期)2022-11-22
广西林业科学(2022年4期)2022-11-07
林业调查规划(2022年3期)2022-06-13
测井技术(2022年2期)2022-06-06
水利科技与经济(2022年3期)2022-03-25
土壤学报(2021年4期)2021-09-10
园艺与种苗(2021年3期)2021-05-26
灌溉排水学报(2021年2期)2021-03-17
南方农业·中旬(2018年11期)2018-05-14
中南林业科技大学学报(2015年8期)2015-05-22
