
珙桐幼苗生理生化指标对重金属铅、镉胁迫的响应
2021-10-16 12:48刘朝荣张柳青杨艳黄兴黎云祥权秋梅朱晓华
广西植物 2021年9期
关键词:抗氧化酶
刘朝荣 张柳青 杨艳 黄兴 黎云祥 权秋梅 朱晓华

摘 要:铅和镉在土壤中表现出很强的毒性,严重危害植物的生长发育。该研究着眼于中国特有濒危珍稀植物珙桐,探究盆栽实验条件下其抗氧化酶活性、丙二醛(MDA)以及游离脯氨酸、可溶性蛋白对不同浓度重金属铅、镉胁迫的响应规律。结果表明:(1)不同浓度铅处理下,珙桐幼苗MDA浓度均显著高于对照组,而镉胁迫条件下除10 mg·kg-1浓度外,其余处理组MDA浓度也显著高于对照组,表明了铅和镉污染加剧了珙桐膜脂过氧化进程。(2)游离脯氨酸随着铅、镉浓度的增加,表现出先增加后降低的现象,分别在铅浓度≥800 mg·kg-1和镉浓度≥20 mg·kg-1处理下显著低于对照组。可溶性蛋白浓度随着铅浓度的增加也表现出先增后减的规律,其浓度在镉胁迫下均显著高于对照组。可溶性蛋白和游离脯氨酸的增加可以提高珙桐抵抗低浓度重金属危害的能力,高浓度重金属对珙桐则产生抑制作用。(3)随着铅和镉浓度的增加,珙桐抗氧化酶活性也表现出先增加后降低的特征,表明了低浓度重金属(铅浓度≤600 mg·kg-1,鎘浓度≤5 mg·kg-1)容易激活珙桐抗氧化应激反应,有效地减少重金属的危害,高浓度的重金属则会抑制抗氧化酶活性。(4)通过相关性和主成分分析表明,珙桐幼苗中抗氧化酶、游离脯氨酸可以较好地反映珙桐对两种重金属胁迫的响应规律。
关键词:珙桐,铅,镉,抗氧化酶,膜脂过氧化
中图分类号:Q945.78
文献标识码:A
文章编号:1000-3142(2021)09-1401-10
Abstract:Lead (Pb)and cadmium (Cd),which are very toxic heavy metals found in soil,severely impair plant growth. This study focuses on Davidia involucrata,a rare relic plant in China,exploring the responses of the antioxidant enzymes,malondialdehyde (MDA)and free proline,soluble protein to different concentrations Pb and Cd by pot experiment. The results were as follows:(1)The concentrations of MDA in D. involucrata seedlings were higher in different concentrations of Pb than that in the control group. The MDA concentrations of the different concentrations of Cd,except for the concentration was 10 mg·kg-1,was significantly higher than that in the control group,indicating the increased degree of cell membrane lipid peroxidation of D. involucrata. (2)Free proline concentration of osmoregulation substances initially increased and then decreased with the increase concentrations of Pb and Cd,which was lower than that in the control group under treatment of Pb concentration ≥800 mg·kg-1 and Cd concentration ≥20 mg·kg-1,respectively. With the increase of Pb concentration,the soluble protein concentration also showed an initial increase and then decrease trend. The concentrations of soluble protein were higher in the treatment groups of Cd than that in the control group. The increase of soluble protein and free proline enhanced the stress resistance of D. involucrata to resist the damage. In addition,the high concentration of heavy metals inhibited the growth of D. involucrata. (3)Antioxidant enzyme activities also initially increased and then decreased with the increase of Pb and Cd concentrations,indicating that low concentrations of heavy metals(Pb concentration≤600 mg·kg-1 and Cd concentration≤5 mg·kg-1)could easily activate the antioxidant capacity of D. involucrata and it could deal with the harm,while high concentrations of heavy metals could inhibit the activities of antioxidant enzymes. (4)The correlation and principal component analysis showed that antioxidant enzymes and free proline in D. involucrata seedlings had important indicative effects on the stresses of the two heavy metals.
Key words:Davidia involucrata,Pb,Cd,antioxidant enzymes,membrane lipid peroxidation
随着城镇化进程的加快,人类活动强度加剧,加速了污染物向大气、水和土壤的释放,进而降低自然资源质量,危害动植物和人类的健康。土壤是重要的物质和能量交换媒介,也是陆生植物生长繁殖的物質基础,由于人为的干扰导致了土壤污染的加剧。其中,土壤重金属污染具有难降解、易迁移、潜伏期长等污染特征,我国受污染最严重的两种重金属污染物是铅(Pb)和镉(Cd)。对于植物而言,铅、镉并非植物生长所需要的元素,但它们会通过植物根系吸附作用迁移到植物体内,过量的铅、镉会抑制植物细胞酶的活性和功能,干扰植物的呼吸和光合作用,进而阻碍植物的生长发育,造成累积毒害现象。植物抗氧化酶通常作为植物抵抗重金属胁迫的重要指示指标,重金属刺激植物产生应激反应,生成较多的超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)等自主清除体内过多的氧自由基。因此,其活性增加表明植物抵御不利环境的能力也增强。另外,游离脯氨酸的积累也是植物在受到不利环境条件下的一种重要解毒机制。可溶性蛋白受到重金属毒害作用后主要表现出变性、降解等现象,从而使酶参与的代谢活动发生功能紊乱(杨惠等,2013;唐探等,2015)。然而,MDA的积累则指示植物受到逆境伤害的程度较高,尤其是细胞膜结构被破坏的程度。
珙桐(Davidia involucrata),俗称鸽子树,属于珙桐科(Davidiaceae)落叶乔木,是第三纪古热带植物区的遗迹种,我国特有属种,已被列为重点保护野生植物。目前,我国现有的野生种仅分布在四川省西南部、湖北省中部及周边地区。由于珙桐具有较高的观赏价值、经济价值和药用价值,已被广泛地引种在自然保护区、风景名胜区等地区(王宁宁等,2011)。许多学者对珙桐展开了研究,主要集中在人工繁殖和引种、种群生态学、细胞学等领域(朱利君等,2007)。贺金生等(1995)研究了珙桐及珙桐林的天然分布特征和分布区的自然环境状况;Tang et al.(2017)运用物种分布模型预测了珙桐气候适宜区域的未来分布状况;刘周莉等(2009)在42 ℃热处理下对珙桐幼苗进行了基于RNA-Seq的基因表达分析,对珙桐的研究有利于提高其存活率以及较好地规划保护措施,具有重要的意义。由于重金属污染物质会通过大气干湿沉降、污水灌溉等进入到土壤中,一部分有害重金属被截留在土壤中,通过植物富集和迁移转化进入植物体内阻碍其正常生长发育、降低生物量等(韩琳等,2020)。珙桐对其生长环境有较高的要求,土壤质量的优劣对珙桐的生长具有重要的影响作用。然而,关于土壤重金属污染对珙桐幼苗的生理生化指标影响的研究较少,相关资料比较缺乏。因此,本研究分析了珙桐幼苗的抗氧化酶、MDA、可溶性蛋白以及游离脯氨酸在不同浓度土壤重金属铅、镉污染条件下的活性或浓度变化规律,以期探讨珙桐对重金属污染的响应机制和潜在影响因素,为进一步研究珙桐在逆境下的生存能力提供了研究资料,也为保护珙桐、提高成活率及制定合理的保护规划提供了参考依据。
1 材料与方法
1.1 实验材料
珙桐幼苗(三年生)均来自四川省什邡市,实验所使用的土壤pH为7.67±0.07,全氮、全磷含量分别为513.47、472.5 mg·kg-1,其中Pb、Cd的背景值含量分别为5.71、0.09 mg·kg-1。
1.2 栽培和处理
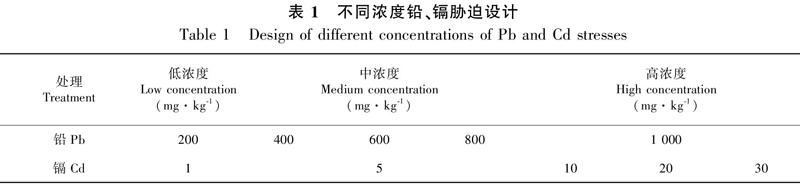
根据土壤环境质量标准(GB15618-1995),使用Pb(NO3)2和CdCl2·2.5H2O制备重金属溶液,分别模拟低浓度、中浓度和高浓度铅和镉胁迫环境(表1)。配制的溶液添加到土壤中并混合均匀,用盆栽土培的实验方法于西华师范大学实验基地培养90 d(2018年6月至2018年8月),每组处理均重复3次。
1.3 生理生化指标测定
1.3.1 株高 重金属处理前,用卷尺测量每株珙桐幼苗的株高,3次重复。经90 d重金属处理后,再次测量每株幼苗的株高,均3次重复。计算株高增量,即处理后株高减去处理前株高。
1.3.2 MDA、游离脯氨酸和可溶性蛋白 摘取同一功能区的叶片3~5片,置于冰盒中,立即送回实验室进行各项指标的检测。MDA采用硫代巴比妥方法测定(AbdElgawad et al.,2019),游离脯氨酸使用磺基水杨酸法测定(Jin et al.,2019),可溶性蛋白采用考马斯亮蓝法测定(Cheng et al.,2016)。
1.3.3 抗氧化酶活性 SOD、POD和CAT采用分光光度计法测定。称取叶片0.2 g于研磨钵中,添加2 mL磷酸盐缓冲液并置于冰浴中快速研磨,然后在4 ℃下10 000 r·min-1离心15 min,收集上清液作为待测样品。根据Donahue et al.(1997)的方法,波长560 nm处的OD值引起50%硝基蓝四氮唑(NBT)还原速率抑制所需的酶量来测量SOD活性。POD活性通过愈创木酚氧化法测定(Merey et al.,2018),测量波长470 nm处吸光度的增加值。CAT活性被定量为底物H2O2的消耗而导致波长240 nm处的吸光度的减少值(Jing et al.,2018)。
1.4 统计分析
采用SPSS 23.0(IBM Inc.,美国)对实验数据进行统计分析,包括均值和标准差、不同处理组间的显著性差异分析以及皮尔逊相关性分析,采用Origin 2018进行主成分分析。
2 结果与分析
2.1 珙桐幼苗株高变化特征
由图1可知,经重金属铅、镉胁迫90 d后,珙桐幼苗的株高均有所增加,随着铅浓度的增加,株高的增量先变大后减小。其中铅浓度在200~800 mg·kg-1范围内对珙桐生长的抑制作用较低,而1 000 mg·kg-1具有较强抑制作用。且600和800 mg·kg-1处理下株高增量显著高于对照组。随着镉浓度的升高,珙桐幼苗株高的增量也出现先增后减的规律,镉浓度为5 mg·kg-1时,株高的增量最明显,但整体上无显著性差异。
2.2 MDA、游离脯氨酸和可溶性蛋白的特征
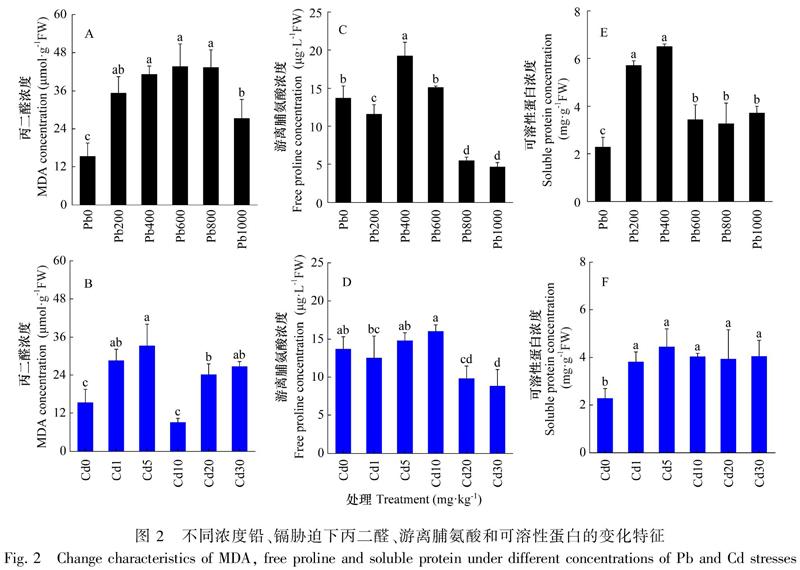
由图2:A,C,E可知,MDA、游离脯氨酸和可溶性蛋白浓度随铅浓度的增加先升高后下降。对照组MDA的浓度显著低于其他处理组(P<0.05),铅浓度在400~800 mg·kg-1范围内,MDA浓度显著高于其他处理组。铅浓度为400 mg·kg-1时,游离脯氨酸的浓度最高(P<0.05);800和1 000 mg·kg-1浓度的铅胁迫下,游离脯氨酸浓度显著降低(P<0.05)。可溶性蛋白浓度在铅浓度为200和400 mg·kg-1时显著高于其他处理组(P<0.05);铅浓度超过400 mg·kg-1时,对可溶性蛋白的影响无显著差异。
镉胁迫对MDA、游离脯氨酸和可溶性蛋白的影响与铅胁迫存在一定的差异(图2:B,D,F)。镉浓度为10 mg·kg-1浓度时,MDA浓度最低,其次是对照组,均显著低于其他处理组(P<0.05)。镉浓度为20和30 mg·kg-1时,游离脯氨酸的浓度显著低于对照组(P<0.05)。不同浓度镉胁迫下可溶性蛋白的浓度均显著高于对照组(P<0.05),但各处理组之间无显著差异。
2.3 SOD、POD和CAT活性特征
随铅浓度的增加,抗氧化酶SOD、POD和CAT的活性先升高后降低。铅浓度为400 mg·kg-1时,SOD活性最高;而铅浓度高于600 mg·kg-1时,SOD活性降低显著低于对照组(P<0.05)。对照组POD活性显著低于其他处理组(P<0.05),铅浓度为200、400和800 mg·kg-1时,POD活性较高。除铅浓度800 mg·kg-1外,各處理组CAT活性均显著高于对照组(P<0.05),其中铅浓度为200 mg·kg-1时CAT活性较高。
镉浓度分别为1、5和20 mg·kg-1时,SOD活性显著高于其他处理组(P<0.05)。POD和CAT活性在镉浓度分别为5和1 mg·kg-1时最高。CAT活性随镉浓度的增加逐渐降低,且镉浓度在10~30 mg·kg-1时显著低于对照组(P<0.05)。
2.4 抗氧化酶、MDA以及渗透调节物质相关性分析
由表2可知,在铅胁迫下,SOD活性与游离脯氨酸、可溶性蛋白显著正相关(P<0.01),POD活性与MDA浓度也显著正相关(P<0.01),CAT活性与可溶性蛋白显著相关(P<0.01)。由表3可知,SOD活性与CAT活性、MDA浓度显著正相关(P<0.01,P<0.05),此外POD活性与MDA、可溶性蛋白也有显著性正相关性(P<0.05)。不同种类重金属对珙桐各项生理生化指标的影响存在差异,铅、镉胁迫下SOD和POD的产生与可溶性蛋白浓度的增加相关,铅胁迫环境下SOD活性的增强与游离脯氨酸浓度增加是相关的。此外,镉胁迫下SOD活性的提升也伴随着CAT活性的增强。
2.5 抗氧化酶、MDA以及渗透调节物质的主成分分析
由图4:A可知,两个主成分共同解释了75.49%的信息,其中PC1解释了45.63%,PC2解释了29.86%。抗氧化酶、MDA、游离脯氨酸和可溶性蛋白为主成分PC1的主要指标,可溶性蛋白与PC1相关性最高。可见3种抗氧化酶、MDA、游离脯氨酸和可溶性蛋白都能在一定程度上反映珙桐对铅胁迫的响应规律。由图4:B可知,PC1和PC2共同解释了64.98%的信息,其中,PC1为39.98%、PC2为25%。除游离脯氨酸外,其余指标均与PC1有较强的正相关性。游离脯氨酸、CAT和SOD与PC2有较强的相关性,MDA和可溶性蛋白与PC2有较强的负相关性。3种抗氧化酶、MDA浓度和可溶性蛋白能较好地指示珙桐对镉的响应特征。
3 讨论
3.1 铅、镉对珙桐幼苗膜脂化、游离脯氨酸和可溶性蛋白的影响
研究发现,膜脂质和蛋白质也是植物在环境胁迫下活性氧的作用目标,因此认为它们是氧化应激可控调节的指示指标(Liu et al.,2015)。MDA是植物中自由基和脂质相互作用的产物,胁迫环境会引起MDA累积,进而造成蛋白质、核酸等大分子的交联聚合,降低植物细胞中蛋白质代谢效果,也会对细胞膜系统、光合作用等产生较大的危害(杨国远等,2014)。研究发现,不同浓度的铅胁迫均会引起珙桐幼苗MDA浓度的显著增加,且铅的浓度在低于800 mg·kg-1范围内,其浓度越高,珙桐幼苗的膜脂过氧化程度也越强。这与王吉秀等(2011)的研究相同,意味着较高浓度的重金属加剧珙桐膜脂化程度。游离脯氨酸和可溶性蛋白是植物体内重要的代谢物质(张亚玲等,2016),游离脯氨酸在保护细胞膜结构、生物大分子结构的稳定性、清除自由基等方面也发挥重要作用。可溶性蛋白也能提高功能蛋白的数量,以维持细胞正常的生理代谢活动,从而提高植物的抗逆性。没有添加重金属条件下,游离脯氨酸较高,但随着铅、镉浓度的增加,游离脯氨酸浓度先升高后降低。研究结果表明,植物叶片中游离脯氨酸浓度会随着胁迫程度的增加而增加(陆海燕等,2013)。这与本研究的结果存在差异,表明在一定范围内,珙桐能通过增加游离脯氨酸和可溶性蛋白来抵抗重金属铅、镉胁迫,但铅、镉浓度过高可能会进一步破坏珙桐的细胞膜系统以及代谢平衡,导致其应激能力降低,因此游离脯氨酸浓度有降低的现象(覃光球等,2006),在不同浓度铅、镉(除10 mg·kg-1外)胁迫下,MDA在各处理组均显著高于对照组,且先增加后降低可以进一步印证,与此同时也说明游离脯氨酸对铅、镉的胁迫更为敏感。镉浓度为10 mg·kg-1时,膜质化程度最低,但珙桐幼苗株高增量也最低,且其他组没有显著差异,这可能与其抗氧化酶活性在高浓度镉胁迫下降低,其活性受到抑制,植物抵抗重金属伤害能力下降有关。铅胁迫下,可溶性蛋白浓度先增加后降低,但均显著高于对照组,这与游离脯氨酸的变化规律相似,表明较低浓度的铅能诱导珙桐产生可溶性蛋白以减轻铅胁迫对珙桐的伤害,而较高的铅浓度会产生抑制作用,这与珙桐膜脂过氧化加剧降低蛋白质功能,导致蛋白质合成体失活有关,也进一步印证了随着重金属浓度的增加,珙桐幼苗生长受到抑制作用增强。除对照组可溶性蛋白浓度最低外,不同浓度的镉对可溶性蛋白的影响无显著性差异,这与珙桐对不同种类重金属胁迫的应激机制存在差异有关,因此可溶性蛋白的响应程度也存在差异,这与唐探等(2015)的研究相似。
3.2 铅、镉对珙桐幼苗抗氧化酶活性的影响
重金属刺激植物产生较多的活性氧自由基,这些氧自由基与脂质、蛋白质、核酸等物质发生反应,引起脂质过氧化、膜损伤和酶失活,从而影响细胞的性能和生存能力,一旦超过植物自我清除的能力,还会导致植物死亡。SOD是一种金属酶,能够催化体内氧化活性很强的超氧自由基(O2-)的歧化反应(陆海燕等,2013),生成氧化活性较弱的H2O2,进而由POD、CAT参与催化细胞内H2O2的分解以防止过氧化反应。本研究发现,铅胁迫环境下,SOD活性存在“高促低抑”的现象,POD和CAT(除800 mg·kg-1外)活性均顯著高于对照组,但在较高浓度铅处理下其活性均出现降低的趋势,这表明了低浓度铅激发了珙桐抗氧化反应,刺激抗氧化酶的产生以去除重金属铅胁迫产生的氧基自由基,但高浓度的铅(800~1 000 mg·kg-1)会严重破坏植物细胞功能,降低了抗氧化系统的应激反应能力,其中SOD活性对铅胁迫最敏感。李铮铮等(2007)的研究也发现鱼腥草在不同浓度铅胁迫下SOD、POD、CAT活性先增加后降低,此外POD和CAT活性的降低也与SOD活性的降低有关。镉浓度为1、5和20 mg·kg-1,POD活性随镉浓度的增加也有先增加后降低的变化规律,较高浓度的镉(10~30 mg·kg-1)胁迫也抑制了CAT的活性,Liu et al.(2010)对镉影响高粱的抗氧化调节作用的研究也有相似的发现。本研究还发现,珙桐幼苗的抗氧化系统能够应对较低浓度的镉(Cd≤5 mg·kg-1)胁迫,清除O2-、H2O2等有害物质以维持植物体内自由基的正常代谢,同时镉的毒性增强会抑制抗氧化系统的反应能力(Yang et al.,2018)。珙桐幼苗株高变化量对铅胁迫更为敏感,不同浓度镉对珙桐幼苗的生长影响不显著。
3.3 铅、镉胁迫下珙桐幼苗的主要指示指标
铅胁迫下,SOD活性与游离脯氨酸、可溶性蛋白均显著正相关,CAT也与可溶性蛋白有显著正相关性,主成分分析中SOD、可溶性蛋白与第一主成分有强烈的相关性,POD活性和游离脯氨酸与第二主成分相关性较强,这意味着抗氧化酶SOD、POD与游离脯氨酸可以作为珙桐幼苗对铅的响应规律的主要参考指标。与铅胁迫不同,镉胁迫下,SOD活性可能会影响CAT活性,SOD、POD活性与膜脂过氧化产物MDA的正相关性表明了珙桐抗氧化性的提高,其膜脂过氧化程度有增加的趋势。SOD、POD、MDA与第一主成分有较强的正相关性,CAT和游离脯氨酸与第二主成分也有强烈的正相关性,也进一步说明了镉胁迫环境下其抗氧化酶、MDA和游离脯氨酸对镉胁迫有重要的指示作用。
4 结论
(1)在低浓度铅、镉(Pb≤600 mg·kg-1,Cd≤5 mg·kg-1)胁迫环境下,珙桐的抗氧化反应的程度较强,且株高增加量较高,意味着珙桐能较好地通过自我防御减轻重金属毒性。
(2)铅、镉胁迫导致MDA有较高的累积,表明2种重金属很容易引起珙桐幼苗细胞膜脂过氧化并损害细胞正常代谢功能。游离脯氨酸浓度随着铅、镉浓度的增加先升高后降低,且可溶性蛋白在铅的胁迫下表现出与游离脯氨酸相同的变化规律。然而,在高浓度铅、镉胁迫下,珙桐幼苗对铅、镉的应激反应能力降低。
(3)珙桐幼苗抗氧化酶在低浓度的铅、镉胁迫下表现出较强的活性,增强了珙桐幼苗对氧自由基等有害物质的清除能力。此外,抗氧化酶活性、游离脯氨酸可以较好地指示珙桐幼苗对重金属铅、镉胁迫的响应规律。我们的发现丰富了珙桐相关领域的研究资料,也为探究珙桐对重金属胁迫环境适应能力的研究提供了参考依据。
参考文献:
ABDELGAWAD H,ZINTA G,BADRELDIN AH,et al.,2019. Maize roots and shoots show distinct profiles of oxidative stress and antioxidant defense under heavy metal toxicity[J]. Environ Pollut,113705.
CHEN K,JIANG XZ,MA SY,et al.,2019. Physiological response and cold resistance evaluation of the leaves of Parashorea chinensis seedlings to low temperature stress[J]. J NW For Univ,34(3):67-73.[陈凯,江秀章,马松亚,等,2019. 望天树苗期叶片对低温胁迫的生理响应及抗寒性评价[ J]. 西北林学院学报,34(3):67-73.]
CHENG YF,WEI HM,SUN R,et al.,2016. Rapid method for protein quantitation by Bradford assay after elimination of the interference of polysorbate 80[J]. Anal Biochem,494:37-39.
DING HD,QI NM,ZHU WM,et al.,2006. Effects of Cd,Zn stress on the growth,contents of proline and GSH of tomato seedlings[J]. Chin J Eco-Agric,14(2):53-55.[丁海东,齐乃敏,朱为民,等,2006. 镉、锌胁迫对番茄幼苗生长及其脯氨酸与谷胱甘肽含量的影响[J]. 中国生态农业学报,14(2):53-55.]
DONAHUE JL,OKPODU CM,CRAMER CL,et al.,1997. Responses of antioxidants to paraquat in pea leaves (relationships to resistance)[J]. Plant Physiol,113(1):249-257.
EWA SP,DRKIEWICZ M,KRUPA Z,2009. Lipid peroxidation andantioxidative response in Arabidopsis thaliana exposed to cadmium and copper[J]. Acta Physiol Plant,32:169-175.
GUO YL,TAI PD,HAN YP,2009. Effects of cadmium on the growth and physiological characteristics of sunflower seedlings[J]. Chin J Environ Eng,3(12):2291-2296.[郭艷丽,台培东,韩艳萍,2009. 镉胁迫对向日葵幼苗生长和生理特性的影响[J]. 环境工程学报,3(12):2291-2296.]
HAN L,XU XB,2020. Quantitative evaluation of human health risk of heavy metals in soils based on positive matrix factorization model and geostatistics[J/OL]. Environ Sci:1-15[2020-06-22].https://doi.org/10.13227/j.hjkx.202004081.[韩琳,徐夕博,2020. 基于PMF模型及地统计的土壤重金属健康风险定量评价[J/OL]. 环境科学:1-15[2020-06-22].https://doi.org/10.13227/j.hjkx.202004081.]
HE JS,LIN J,CHEN WL,1995. The current status of endemic and endangered species Davidia involucrata and the preserving strategies[J]. Chin Biodivers,3(4):213-221.[贺金生,林洁,陈伟烈,1995. 我国珍稀特有植物珙桐的现状及其保护[J]. 生物多样性,3(4):213-221.]
HU J,MENG DL,LIU XD,et al.,2018. Response of soil fungal community to long-term chromium contamination[J]. Trans Nonferrous Metal Soc Chin,28(9):1838-1846.
JIA X,ZHOU CJ,DONG SM,2011. Progress of research on the effects of Cd2+ stress on wheat and the response of wheat to Cd2+[J]. J Triticeae Crops,31(4):786-792.[贾夏,周春娟,董岁明,2011. 镉胁迫对小麦的影响及小麦对镉毒害响应的研究进展[J]. 麦类作物学报,31(4):786-792.]
JIANG MY,GUO SC,ZHANG XM,1997. Proline accumulation and antioxidant activity in rice seedlings under -OH stress[J]. Chin Sci Bull,42(6):647-649.[蒋明义,郭绍川,张学明,1997. -OH 胁迫下稻苗体内脯氨酸积累及其抗氧化作用[J]. 科学通报,42(6):647-649.]
JIN Y,QU JJ,LI Y,2013. Isolation,identification and Pb (Ⅱ)biosorption characterization of a lead-resistant strain[J]. Acta Sci Circum,33(8):2248-2255.[金羽,曲娟娟,李影,2013. 一株耐铅细菌的分离鉴定及其吸附特性研究[J]. 环境科学学报,33(8):2248-2255.]
JIN YQ,ZENG CM,LV QF,et al.,2019. Efficient adsorption of methylene blue and lead ions in aqueous solutions by 5-sulfosalicylic acid modified lignin[J]. Int J Biol Macromol,123:50-58.
LI ZZ,WU J,TANG Y,et al.,2007. Effect of Pb,Zn and their interactions on the chlorophyll content and antioxidant enzyme systems of Houttuynia cordata Thunb[J]. Acta Agron Sin,(12):5441-5446.[李铮铮,伍钧,唐亚,等,2007. 铅、锌及其交互作用对鱼腥草 (Houttuynia cordata)叶绿素含量及抗氧化酶系统的影响[J]. 生态学报,(12):5441-5446.]
LIN L,ZHOU WH,DAI HX,et al.,2012. Selenium reduces cadmium uptake and mitigates cadmium toxicity in rice[J]. J Hazardous Mat,235-236:343-351.
LIU B,LIANG CJ,2005. Recent advances of catalase in organism[J]. Chin Agric Sci Bull,21(5):223-232.[刘冰,梁婵娟,2005. 生物过氧化氢酶研究进展[J]. 中国农学通报,21(5):223-232.]
LIU DL,ZHANG SP,CHEN Z,et al.,2010. Soil cadmium regulates antioxidases in sorghum[J]. Agric Sci Chin,9(10): 1475-1480.
LIU QS,VETUKURI RR,XU X,et al.,2019. Transcriptomic responses of dove tree (Davidia involucrata Baill.)to heat stress at the seedling stage[J]. Forest,10(8): 656.
LIU XF,HOU F,LI GK,et al.,2015 Effects of nitrogen dioxide and its acid mist on reactive oxygen species production and antioxidant enzyme activity in Arabidopsis plants[J]. J Environ Sci,34(8):93-99.
LIU ZL,HE XY,CHEN W,2009. Effects of cadmium stress on the growth and physiological characteristics of Lonicera japonica[J]. J Appl Ecol,20(1):40-44.[刘周莉,何兴元,陈玮,2009. 镉胁迫对金银花生理生态特征的影响[J]. 应用生态学报,20 (1):40-44.]
LU HY,LIU ZH,L GH,2013. Reaction of MDA,proline,and SOD under Cd stress in mixture of Phragmites australiss stems and leaves[J]. J Arid Land Res Environ,27(8):171-175.[陆海燕,刘志辉,吕光辉,2013. 镉污染下芦苇叶片丙二醛、脯氨酸及SOD保护酶反应研究[J]. 干旱区资源与环境,27(8):171-175.]
MALEKI M,MANSOUR G,KHALIL K,2017. Physiological and antioxidative responses of medicinal plants exposed to heavy metals stress[J]. Plant Gene,11:247-254.
MEREY HA,RAMADAN NK,DIAB SS,et al.,2018. Validated UPLC method for the determination of guaiphenesin,oxeladin citrate,diphenhydramine,and sodium benzoate in their quaternary mixture used in treatment of cough,in the presence of guaiphenesin-related substance (guaiacol)[J]. Chem Pap,72(9):2247-2254.
QI BZ,1989. The effects of sulfur nutrition on some physiological parameters in relation to carbon and nitrogen metabolism in wheat and maize[J]. Acta Agr Sin,15(1):31-35.[祁葆滋,1989. 硫營养对小麦、玉米碳、氮代谢中几项生理参数的影响[J]. 作物学报,15(1):31-35.]
QIN GQ,YAN CL,WEI LL,2006. Effect of cadmium stress on the contents of tannin,soluble sugar andproline in Kandelia candel (L.)Druce seedlings[J]. Acta Ecol Sin,26(10):3366-3371.[覃光球,严重玲,韦莉莉,2006. 秋茄幼苗叶片单宁、可溶性糖和脯氨酸含量对Cd胁迫的响应[J]. 生态学报,26(10):3366-3371.]
SHI J,SUN JJ,FANG BJ,et al.,2018. Photoluminescence performance ofEr/Yb co-doped NBT ceramics prepared via hydrothermal method[J]. J Physics Chem Solids,121:228-235.
SPONGBERG SA,1993. Exploration and introduction of ornamental and landscape plants from Eastern Asia[J]. N Crop N Y:Wiley 140-147.
TANG CQ,DONG YF,HERRANDO-MORAIRA S,et al.,2017. Potential effects of climate change on geographic distribution of the tertiary relict tree species Davidia involucrata in China[J]. Sci Rep,7:43822.
TANG T,JIANG YL,ZHANG Y,et al.,2015. Physiological characteristics of Cinnamomum glanduliferum seedlings under lead and cadmium stress[J]. Jiangsu Agric Sci,43(1):199-201.[唐探,姜永雷,张瑛,等,2015. 铅、镉胁迫下云南樟幼苗的生理特性变化[J]. 江苏农业科学,43(1): 199-201.]
TIAN GZ,LI HF,QIU WF,2001. Advances on research of plant peroxidases[J]. J Wuhan Bot Res,19(4):332-344.
WANG JX,ZU YQ,LI Y,2011. Effects of maize and vegetable intercropping system on accumulation of Pb,Cu and Cd in plants[J]. J Agro-Environ Sci,30(11):2168-2173.[王吉秀,祖艳群,李元,2011. 玉米和不同蔬菜间套模式对重金属 Pb、Cu、Cd 累积的影响研究[J]. 农业环境科学学报,30(11):2168-2173.]
WANG NN,HU ZH,SHEN YB,2011. Photosynthetic characteristics of Davidia involucrata Baill. seedlings under soil drought stress[J]. Acta Bot Boreal-Occident Sin,31(1):101-108.[王宁宁,胡增辉,沈应柏,2011. 珙桐苗木叶片光合特性对土壤干旱胁迫的响应[J]. 西北植物学报,31(1):101-108.]
WANG X,XU HJ,LIU T,2010. Comparison of the physiological response of Chinese cabbage between cerium and lead stress[J]. J Nucl Agric Sci,24(3):634-638.[王学,徐恒戬,刘涛,2010. 白菜对铈、铅胁迫响应的比较研究[J]. 核农学报,24(3):634-638.]
WANG XJ,WANG WB,YANG L,et al.,2015. Transport pathways of cadmium (Cd)and its regulatory mechanisms in plant[J]. Acta Ecol Sin,35(23):7921-7929.[王晓娟,王文斌,杨龙,等,2015. 重金属镉(Cd)在植物体内的转运途径及其调控机制[J]. 生态学报,35(23):7921-7929.]
WU G,XIAO HD,LI J,et al.,2000. Relationship between human activities and survival of rare and endangered species Davidia involucrata[J]. J Appl Ecol,11(4):493-496.
XU JJ,LIU NN,QIN L,et al.,2016. Effects of Cd stress on antioxidant enzymes activity of Sonchus asper L. Hill. and Zea mays L. in intercropping system[J]. J Yunnan Agric Univ(Nat Sci Ed),30(2):348-355.[徐佳佳,刘宁宁,秦丽,等,2016. 镉胁迫对续断菊与玉米间作体系中植物叶片抗氧化酶活性的影响[J]. 云南农业大学学报(自然科学版),31(2):348-355.]
YANG GY,WAN LL,LEI XQ,et al.,2014. Effects of lead and chromium on the growth,photosynthetic performance,and antioxidant activity of Scenedesmus obliquus[J]. Acta Sci Circum,34(6):1606-1614.[楊国远,万凌琳,雷学青,等,2014. 重金属铅、铬胁迫对斜生栅藻的生长、光合性能及抗氧化系统的影响[J]. 环境科学学报,34(6):1606-1614.]
YANG H,ZHAI MZ,LI L,et al.,2013. Cold-tolerance andosmoregulation mechanism of walnut cultivars[J]. Acta Bot Boreal-Occident Sin,33(10):2003-2009.[杨惠,翟梅枝,李丽,等,2013. 不同核桃品种的耐寒性及其渗透调节机制[J]. 西北植物学报,33(10):2003-2009.]
YANG LP,ZHU J,WANG P,et al.,2018. Effect of Cd on growth,physiological response,Cd subcellular distribution and chemical forms of Koelreuteria paniculata[J]. Ecotox Environ Safe,160:10-18.
YANG Y,LI YX,XU X,et al.,2016. The effect of short time treatment of two kinds of heavy metal ions on photosynthesis in Davidia involucrata seedlings[J]. J Chin West Norm Univ(Nat Sci Ed),37(2):163-168.[杨艳,黎云祥,胥晓,等,2016. 两种重金属离子短时处理对珙桐幼苗光合作用的效应[J]. 西华师范大学学报(自然科学版),37(2):163-168.]
ZENG QY,LING QP,YANG ZD,et al.,2019. Effects of copper stress on growth and antioxidant enzymes in sugarcane[J]. Guihaia,39(7):951-958.[曾巧英,凌秋平,楊湛端,等,2019. 铜胁迫对甘蔗生长及抗氧化酶活性的影响[J]. 广西植物,39(7):951-958.]
ZHANG JX,LI JQ,ZHOU BS,et al.,1995. Natural distribution of Davidia involucrata and introduction analysis[J]. J Beijing For Univ,17(1):25-30.[张家勋,李俊清,周宝顺,等,1995. 珙桐的天然分布和人工引种分析[J]. 北京林业大学学报,17(1):25-30.]
ZHANG SY,ZHANG Q,LI L,2019. On tolerance of Ficus virens Aiton in soil contaminated by Pb and Cd[J]. J SW Chin Norm Univ(Nat Sci Ed),44(1):79-83.[张思悦,张晴,李凌,2019. 黄葛树对土壤铅、镉污染耐受性的研究[J]. 西南师范大学学报(自然科学版),44(1):79-83.]
ZHANG YL,XING HW,GUO N,et al.,2016. The physiological characteristics of ornamental kale for cold resistance[J]. Acta Agric Boreal-Sin,31(4):168-176.[张亚玲,幸宏伟,郭宁,等,2016. 羽衣甘蓝抗寒生理特性研究[J]. 华北农学报,31(4):168-176.]
ZHAO ZQ,XI MZ,2007. Advances in research on mechanisms of Cd induced oxidative stress in plants[J]. J Agro-Environ Sci,26:47-51.[赵中秋,席梅竹,2007. Cd 对植物的氧化胁迫机理研究进展[J]. 农业环境科学学报,26:47-51.]
ZHOU JJ,ZHOU J,FENG RG,2014. Status of Chinas heavy metal contamination in soil and its remediation strategy[J]. Bull Chin Acad Sci,29(3):315-320.[周建军,周桔,冯仁国,2014. 我国土壤重金属污染现状及治理战略[J]. 中国科学院院刊,29(3):315-320.]
ZHU LJ,SU ZX,HU JY,et al.,2007. SOD activity of rare plant Davidia involucrata[J]. Chin J Ecol,26(11):1766-1770.[朱利君,苏智先,胡进耀,等,2007. 珍稀濒危植物珙桐超氧化物歧化酶活性[J]. 生态学杂志,26(11):1766-1770.]
ZHUANG GT,2015. Current situation of national soil pollution and strategies on prevention and control[J]. Bull Chin Acad Sci,30(4):477-483.[庄国泰,2015. 我国土壤污染现状与防控策略[J]. 中国科学院院刊,30(4):477-483.]
(责任编辑 周翠鸣)
猜你喜欢
中国医药导报(2016年33期)2017-03-06
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
安徽农学通报(2016年24期)2017-01-12
江苏农业科学(2015年11期)2016-01-27
江苏农业科学(2015年11期)2016-01-27
河北渔业(2015年10期)2015-10-15
江苏农业科学(2015年8期)2015-09-10
湖南师范大学学报·自然科学版(2015年3期)2015-07-17
