
稻瘟病菌效应蛋白MoIEEPl功能初探
2021-11-12 19:29张茜茜刘文德李智强
植物保护 2021年4期
张茜茜 刘文德 李智强

中图分类号: S435.111.41 文献标识码: A DOI: 10.16688/j.zwbh.2020118
水稻是最重要的粮食作物之一,供养着全世界50%以上的人口。然而,由稻瘟病菌引起的水稻全生育期病害极其严重且难以控制。研究显示,稻瘟病造成的水稻年均减产量可大约供给6000万人口的稻米需求。稻瘟病菌无性态为灰梨孢犘狔狉犻犮狌犾犪狉犻犪狅狉狔狕犪犲Sacc,属半知菌亚门,丛梗孢纲,丛梗孢目,丛梗孢科,梨形孢属;有性态为犕犪犵狀犪狆狅狉狋犺犲狅狉狔狕犪犲BarrYaegash,子囊菌亚门。其侵染阶段分为:首先,在适宜温湿度条件下,稻瘟病菌分生孢子梗产生分生孢子,作为侵染循环的接种体;其次,借助空气和风雨的传播,分生孢子利用尖端的黏液附着在植物组织表面,2h后萌发形成芽管,8h后特异性分化产生附着胞;成熟的附着胞随即形成侵染钉穿透植物表皮细胞,侵入植物体内;最后,稻瘟病菌菌丝在寄主细胞内和组织间生长,并在侵染4~5d后形成病斑,寄主细胞死亡。空气湿度较高的情况下,稻瘟病菌侵染菌丝再次产生分生孢子梗和分生孢子,形成新一轮侵染循环,造成病害的不断产生与扩散。自稻瘟病菌全基因组测序完成以来,己有上百个基因被预测并证明可能与稻瘟病菌的致病性相关。如敲除稻瘟病菌MoDuol基因后,发现其突变体分生孢子发育异常,营养菌丝生长减缓,致病力明显减弱。有些基因则与稻瘟病菌附着胞和侵染钉的形成有关,如Pmk1、MoMSB2、MoSHO1、Com1等,进而影响其致病性。因此,稻瘟病菌致病相关基因及其与水稻的互作研究是寻找新的防病途径的关键,具有重要的科学意义和实践价值。
为了顺利侵入寄主植物,病原菌需要经历一系列形态和生理变化,其中包括抑制或克服植物免疫系统,扰乱寄主新陈代谢和信号传导,促进其自身生长。相应地,植物体内进化出一套相对完善的防御系统。该系统包括两个层面。第一是植物细胞表面模式识别受体识别保守的病原菌相关分子模式(PAMP)激发植物基础防卫反应PTI(PAMPtriggeredimmunity);第二是植物抗病蛋白直接或间接识别病原菌分泌的效应蛋白产生的更强烈的免疫防卫反应ETI(Effectortriggeredimmunity),通常在侵染位点引起寄主细胞死亡。在植物和病原菌激烈的“军备竞赛”中,PTI是寄主植物先天免疫的基础防线。寄主植物模式识别受体识别到鞭毛蛋白、碳水化合物、脂质和小分子等病原菌相关分子模式即可抵御病原菌侵染。随之病原菌协同进化出多种途径直接或间接抑制植物PTI反应,其中最重要的是通过分泌效应蛋白干扰免疫反应。大多数真菌效应蛋白N端具有信号肽,利用内质网-高尔基体蛋白分泌系统分泌到细胞外基质,或被转运至植物细胞特定空间发挥作用。基因组、转录组和蛋白质组等相关生物技术的不断发展,使越来越多与水稻互作的稻瘟病菌效应蛋白先后被鉴定。稻瘟病菌Slp1基因编码一个具有分泌功能的LysM 效应蛋白,通过与水稻模式识别受体几丁质激发子结合蛋白(CEBIP)竞争结合几丁质,抑制植物免疫反应。AVRPITA 和PWL2含有分泌信号肽且在真菌侵入、定殖和晚期感染过程中特异表达。犪犮犲1是已克隆无毒基因中唯一编码非分泌蛋白的基因,控制合成一种次生代谢物,侵染水稻时特异性积累在附着胞细胞质中。
鉴定更多的稻瘟病菌效应蛋白并明确其在寄主细胞中的可能作用位置及其致病性不仅是当前研究“水稻稻瘟病菌”分子互作机制的热点内容,而且对稻瘟病防治及培育抗性品种等工作具有极其重要的意义。前期研究中,本实验室应用RLSAGE、MPSS与SBS转录组高通量分析技术,检测到MoIeep1分泌蛋白基因在稻瘟病菌侵染寄主植物时诱导表达。为进一步了解该基因的功能,本文对MoIEEP1(MGG_00083)蛋白的预测信号肽进行了分析与功能验证,亚细胞定位分析与验证了该蛋白的定位,并构建该基因的敲除突变体,解析其生长表型与致病性,为进一步研究该效应蛋白的功能奠定了基础。
1 材料与方法
1.1 试验材料水稻品种为‘日本晴和‘CO39。稻瘟病菌菌株为Guy11,农杆菌Agrobacterium tumefaciens菌株为EHA105,克隆菌株为大肠杆菌Escherichia coli菌株DH5α和JM109,酵母突变体菌株为YTK12。试验用载体为用于信号肽验证的pSUC2和用于水稻亚细胞定位的pYBA1132,以上试验材料均由本实验室保存。用于酵母菌培养的SD/T培养基、蔗糖培养基、YPD培养基的配制参考文献。
1.2 试验方法
1.2.1 效应蛋白基因克隆与载体构建
根据真菌数据库(https:∥fungidb.org/fungidb/)中MoIeep1(GeneID:MGG_00083)基因編码区(CDS)序列信息,采用PrimerPremier5.0软件设计包含全长开放阅读框(fulllength,FL)和不含预测信号肽的截短编码区(nosignalpeptide,NS)目的基因引物,正反向引物前端加上合适的酶切位点及同源序列,用于载体构建;以稻瘟病菌菌株Guy11菌丝cDNA为模板,PCR扩增目的基因并进行琼脂糖凝胶检测和纯化回收;对pSUC2和pYBA1132载体相应酶切位点进行双酶切,胶回收后分别与MoIeep1基因不同片段重组连接并转化至大肠杆菌感受态细胞中,挑取适量单克隆菌落进行PCR 筛选和测序验证;确认序列准确的单菌落经扩大培养后提取质粒,-20℃保存备用。引物合成和测序由北京擎科生物技术公司完成。
1.2.2 效应蛋白转录分析
参照吴立叶等的稻瘟病菌菌丝和孢子样品收集方法,获得Guy11新鲜菌丝和孢子样品,液氮速冻,-80℃保存备用。喷雾接种法将Guy11分生孢子接种于生长1周的大麦叶片背面,分别在接种后2h(芽管形成阶段)、8h(附着胞形成阶段)、18、24、42h撕取叶片下表皮,液氮速冻,-80℃保存备用。
TRIzol法提取稻瘟病菌Guy11菌丝、孢子及不同侵染时期大麦样品总RNA,获得的总RNA 用全式金生物公司RNA 反转录试剂盒合成cDNA 作为荧光定量PCR(qRTPCR)模板。本试验使用引物见表1。内参基因MoActin(MGG_03982)作为对照,基因表达量用相对阈值法(2-ΔΔCT)计算,软件GraphPadPrism5分析并作图。
1.2.3 效应蛋白生物信息学分析
利用SignalP(http:∥www.cbs.dtu.dk/services/SignalP5.0/)在线平台预测MoIEEP1氨基酸序列信号肽的有无和具体位置;WoLFPSORT(https:∥wolfpsort.hgc.jp/)在线平台预测MoIEEP1在植物中的亚细胞定位。网站预测过程中,均需首先输入MoI
EEP1氨基酸序列Fasta格式,信号肽分析选择“Organismgroup”为“Eukarya”,亚细胞定位分析选择相应物种类型为“Plant”,然后直接提交即可获得预测结果。
1.2.4 效应蛋白信号肽验证
LiAc法制备酵母突变体菌株YTK12感受态细胞,并将构建好的含有目的基因MoIeep1的pSUC2载体质粒进行转化。挑取转化成功的酵母单克隆于SD/T液体培养基中,28℃,180r/min过夜培养。当OD600达到0.8时,菌液稀释1000倍划线于蔗糖筛选培养基上,观察酵母菌株在培养基上的生长情况。
1.2.5 效应蛋白亚细胞定位验证
高渗透压下,利用以纤维素酶和果胶酶为主要成分配制的酶解液消化生长10d的‘日本晴水稻黄化苗细胞壁,使之结构松散破坏;随后移除酶解液,用普通渗透压,浓度为0.6mol/L的缓冲液使细胞吸水,原生质体冲破细胞壁释放出来,离心收集原生质体。再将构建好的含有目的基因MoIeep1的pYBA1132载体质粒转化到水稻原生质体中,光照培养箱28℃,L∥D=12h∥12h条件下孵育48h后,激光共聚焦显微镜观察。
1.2.6 MoIeep1基因敲除突变体的构建
依据改进的SplitPCR方法进行目的基因敲除。首先根据稻瘟病菌犕狅犐犲犲狆1基因组DNA 序列信息,设计两轮PCR 扩增引物;然后以稻瘟病菌Guy11基因组DNA 为模板,LBCK/LBR和RBF/RBCK两对引物,分别扩增目的基因上游LB 片段和下游RB片段,并以pCX62载体为模板,HYGF/HYGR扩增完整潮霉素(HYG)基因;再分别以片段LB和HYG共同作为模板,LBF和HYGR1为引物,扩增出片段LB+HY,以片段RB和HYG 共同作为模板,HYGF1和RBR为引物,扩增出片段YG+RB;最后将两个DNA 融合片段共同转化至稻瘟病菌Guy11原生质体;用含有潮霉素抗性的培养基筛选转化子并进行DNA 和RNA 水平的验证,最终获得阳性敲除菌株。
1.2.7 MoIeep1突变体生长表型分析
用孔径6mm 打孔器从生长一周左右的稻瘟病菌野生型菌株Guy11和2个MoIeep1敲除突变体菌落边缘取同等大小的菌块,同时接种于完全培养基(CM)正中央,28℃黑暗培养7d后观察菌落生长形态并测量菌落直径。
1.2.8 MoIeep1突变体致病力检测
稻瘟病菌野生型菌株Guy11和突变体菌株于25℃养菌室暗培养3d后恢复光照,7~10d后产生大量孢子。用0.05%的Tween20水溶液洗下分生孢子,并调整孢子悬浮液浓度为1.5×105个/mL,再均匀喷施在三叶一心期‘CO39水稻叶片上,避免孢子悬浮液液滴凝聚,影响接种效果。接种后于植物光照培养箱中26~28℃,覆膜黑暗保湿培养24h,然后恢复正常光照,温度不变,光照和黑暗各12h,湿度为80%~90%,7d后调查病情并统计病斑密度。即统计叶片上一定面积的病斑数,计算不同病原菌接种后的平均病斑数(个/cm2)。
2 结果与分析
2.1 稻瘟病菌效应蛋白基因MoIeep1转录分析
稻瘟病菌侵染水稻过程中,黏附在水稻表面的分生孢子先产生芽管,再逐渐分化出一种特殊的圆顶状休眠结构———附着胞,附着胞通过积累高浓度的相容性溶质产生巨大的膨压,细胞膨压再转换成机械动力,促使侵染钉穿透叶角质和表皮细胞壁。稻瘟病菌侵染早期表达的效应蛋白更有可能与致病性相关。我们利用qRTPCR 技术,验证了稻瘟病菌MoIeep1 在不同生长和侵染时期的表达水平。如图1所示,MoIeep1在侵染初期8h(附着胞形成阶段)表达量最高,而在菌丝、孢子等营养生長阶段表达量较低,侵染寄主8h后的表达量呈逐渐降低趋势。推测该基因可能在稻瘟病菌被寄主植物识别或进行活体营养侵染阶段发挥重要作用。
2.2 稻瘟病菌效应蛋白犕MoIEEP1信号肽分析与验证
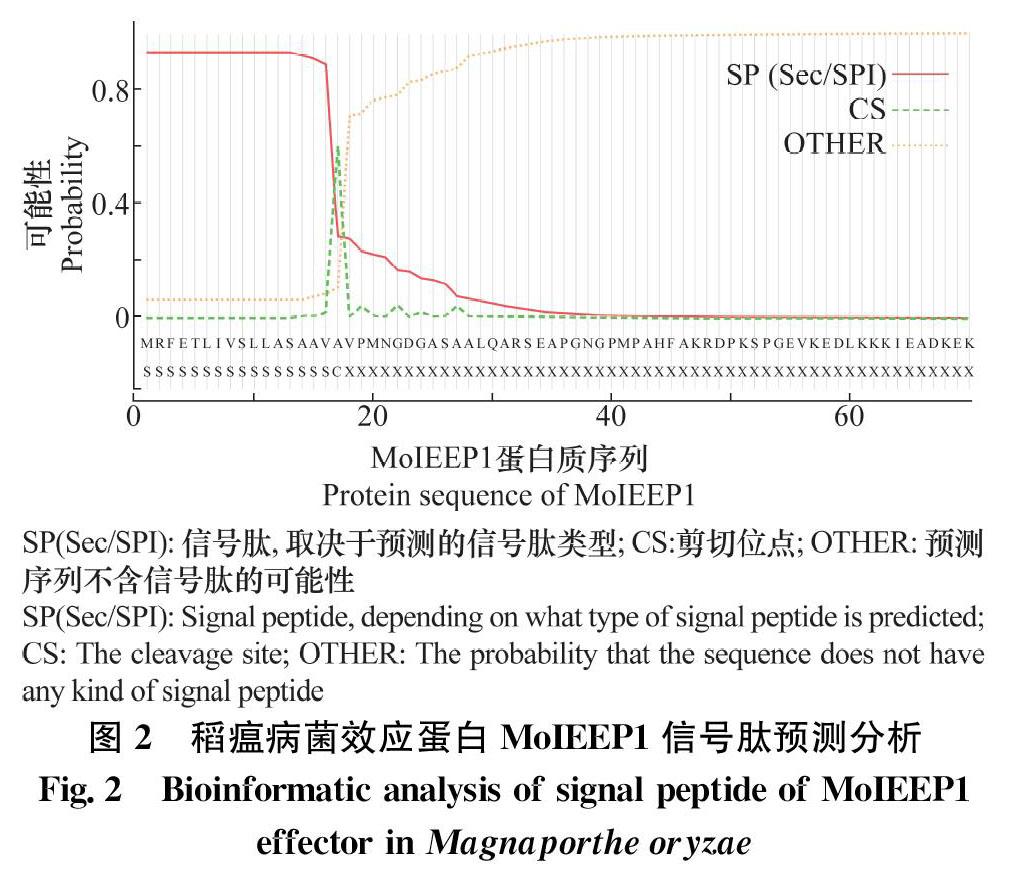
信号肽是真菌效应蛋白N端的一段氨基酸序列,使效应蛋白在病原菌入侵寄主植物时具有分泌性。首先利用SignalP在线分析软件预测了MoIEEP1蛋白的信号肽,结果表明,该蛋白N端氨基酸序列具有信号肽,且其信号肽切割位点为17/18(图2)。随后构建了2个MoIeep1基因的pSUC2(含蔗糖转化酶基因)转化质粒并分别转化到酵母突变体菌株YTK12中。YTK12菌株不含蔗糖转化酶基因,仅能在YPD培养基上生长,而不能在以蔗糖为唯一碳源的筛选培养基上生长。试验结果表明(图3),转化Avr1b_pSUC2(阳性对照)的YTK12菌株能在蔗糖筛选培养基上正常生长,但含有mg87_pSUC2(阴性对照)的YTK12菌株因mg87不具有分泌性,不能在蔗糖筛选培养基上生长。含MoIEEP1_FL_pSUC2序列的菌株能在蔗糖筛选培养基上生长,而含MoIEEP1_NS_pSUC2序列的菌株不能在蔗糖筛选培养基上生长,说明该效应蛋白含有信号肽序列,为分泌蛋白。
2.3稻瘟病菌效应蛋白犕狅犐犈犈犘1亚细胞定位分析与验证
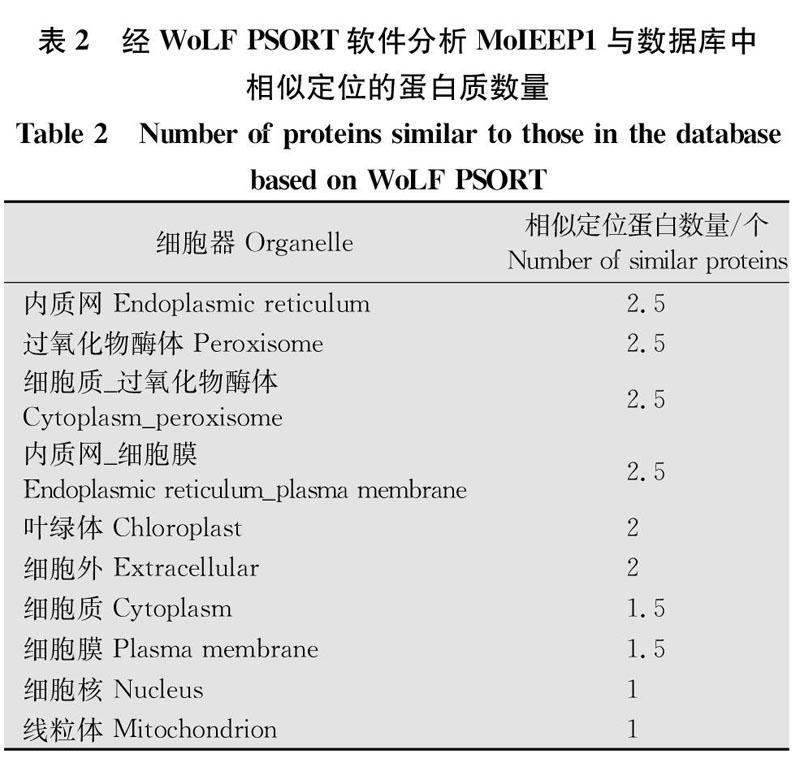
病原菌效应蛋白在水稻细胞中的亚细胞定位与其功能的发挥密切相关。采用WoLFPSORT软件对MoIEEP1蛋白进行亚细胞定位分析。该软件数据库来源于UniProt网站,分为真菌、植物和动物3个方面,各包含2113、2333和12771个蛋白质。基于分类信号、氨基酸组成和功能修饰(如DNA结合基序),WoLFPSORT将蛋白质氨基酸序列转换为数字定位特征,然后使用k近邻分类法预测其亚细胞定位。预测结果以两种方式显示:Ⅰ)与查询序列最相似定位特征的已知定位蛋白质个数;Ⅱ)关于各个已知定位蛋白的详细特征信息。如表2、表3所示,该软件共筛选到14个MoIEEP1最相似定位蛋白。其中与定位在内质网、内质网和质膜、过氧化物酶体、细胞质和过氧化物酶体中的相似蛋白质个数相等且最多;其次是叶绿体和细胞外基质。所以MoIEEP1蛋白极有可能定位在内质网或过氧化物酶体等细胞器。激光共聚焦观察亚细胞定位结果显示:MoIEEP1蛋白質全长或去信号肽均在水稻原生质体中呈明亮点状的定位信号,排除内质网定位,初步推测该定位信号为过氧化物酶体或线粒体等细胞器(图4)。该结果与软件预测内容相互辅正,可信度较高。
2.4 MoIeep1基因敲除突变体的获得
本研究利用SplitPCR方法构建MoIeep1基因敲除突变体。通过两轮PCR 扩增待敲除基因两侧的同源片段分别融合部分潮霉素基因,再将两个DNA片段直接转化至稻瘟病菌Guy11原生质体,两个片段经过同源重组构成完整的潮霉素基因,进而替换目的基因,获得阳性敲除菌株(图5a)。整个试验过程操作方便,转化效率高。潮霉素抗性培养基上筛选的阳性转化子,需分别提取菌丝DNA 和RNA进一步验证。DNA水平上,目的基因内部引物MoIEEP1NF/NR和基因上下游1.5kb处的正反向引物UA/DB 与潮霉素基因的正反向引物H708/H853同时验证(图5b);RNA水平上,仅用基因内部正反向引物鉴定即可(图5c)。结果如图所示。本试验成功获得2个目的基因敲除突变体,后续试验中分别命名为MoIEEP1KO1、MoIEEP1KO2。
2.5 MoIeep1敲除突变体表型鉴定
生长表型方面,稻瘟病菌野生型菌株Guy11和MoIeep1敲除突变体在CM 培养基生长一周的表型如图6所示,突变体和野生型在菌落形态和生长速率方面无明显差异,和图7中菌落生长直径测量结果一致,表明MoIeep1基因可能不直接参与稻瘟病菌的营养生长过程;致病性方面,水稻叶片发病情况和病斑密度统计结果见图8和图9,相比于野生型菌株,MoIeep1敲除突变体致病性轻微减弱。因此,该基因可能对稻瘟病菌的致病性产生一定的影响。
3 讨论
稻瘟病是最具破坏性的植物病害之一,每年可造成10%~30%的产量损失,严重时甚至造成颗粒无收,俗称水稻的癌症。目前,施用化学农药和培育抗性品种是防治稻瘟病的主要方法。随着生态环境建设思想的贯彻执行和人民综合素质的不断提高,化学农药的使用逐渐减少,水稻抗性品种的选育和种植成为最经济有效的措施。深入了解水稻和稻瘟病菌的分子互作机制,有助于培育更加优质高效的水稻抗病品种。稻瘟病菌是最早完成全基因组测序的病原真菌。Dean等解析了稻瘟病菌实验室菌株70-15总长度38.8Mb的序列,分析表明其含有11109个预测基因,其中包含1032个编码分泌蛋白的基因。事实上,稻瘟病菌侵染水稻是一个复杂的过程,大量分泌蛋白参与其中。例如,稻瘟病菌犕犆96基因编码一个分泌蛋白。研究显示,该基因突变后,不仅会显著降低其致病性,而且会影响稻瘟病菌侵染菌丝的扩展。Mosquera等研究表明,真菌侵染早期分泌的效应蛋白具有较高的研究价值。已发表的稻瘟病菌犌犃犛1和犌犃犛2基因在附着胞形成过程中特异表达且编码蛋白定位于细胞质,预测有信号肽,表示它们可能从附着胞移动到侵染菌丝,或运送到寄主植物细胞内发挥作用;Catanzariti等还通过筛选亚麻锈菌吸器cDNA文库,鉴定了21个特异表达的分泌蛋白。因此,本研究对在稻瘟病菌侵染初期8h诱导表达的MoIeep1基因进行了初步的功能鉴定和分析,期望为进一步开展该基因在后续水稻稻瘟病菌分子互作机制研究中提供借鉴。
信号肽位于分泌蛋白的N 端,引导分泌蛋白离开其合成所在的细胞到其他组织细胞。侵染寄主植物期间,病原真菌分泌蛋白干扰寄主免疫防卫反应。本研究采用酵母系统验证目的基因的信号肽。酵母是一种高效真核表达体系,本身含有较少的分泌蛋白,且酵母suc2基因编码蔗糖转换酶,可在以蔗糖为唯一碳源的筛选培养基上生长,而酵母突变体菌株YTK12因缺乏suc2基因,不能将蔗糖或棉籽糖转化为可直接吸收利用的单糖,因此不能在蔗糖或棉籽糖培养基上生长。利用该特性,我们分别将含有全长和不含信号肽的MoIeep1基因的pSUC2载体(含有suc2基因)转化至YTK12酵母突
变体菌株感受态细胞并挑取单菌落涂板,若该基因含有信号肽,则只有含有该基因全长的pSUC2载体(MoIEEP1_FL_pSUC2)可将蔗糖转换酶分泌至胞外,并把培养基中的蔗糖转化为葡萄糖或果糖,从而在蔗糖培养基上正常生长,而不含信号肽的pSUC2载体(MoIEEP1_NS_pSUC2)则不能在蔗糖培养基上生长。本文研究结果与该假设一致,表MoIeep1基因的信号肽具有分泌活性,可能通过分泌到胞外执行生物学功能。
蛋白质的合成在核糖体上进行,信号肽是蛋白质的一个片段。“信号假说”认为信号肽引导核糖体定位并附着于内质网上,并使不断伸长的蛋白质链通过水溶性通道,随后信号肽被切割,合成的蛋白质被释放进内质网腔,随蛋白分泌系统转运到胞外。因此,信号肽不仅与蛋白质的分泌特性相关,也与蛋白质在细胞内的定位有关。近年来,原生质体瞬时表达系统广泛应用于水稻功能基因组学研究。为进一步探究MoIEEP1效应蛋白在水稻细胞中的作用位点,结合在线平台WoLFPROST 的亚细胞定位预测结果,将目的基因构建到含有绿色荧光蛋白基因的表达载体pYBA1132上并转化至水稻原生质体细胞,激光共聚焦观察。该方法可以保持效应蛋白的天然特性,灵敏度高,易于操作,试验周期相对较短。试验结果表明,该蛋白定位于水稻细胞的过氧化物酶体或线粒体上,初步证明MoIEEP1可能通过影响寄主植物过氧化物酶体或线粒体上的蛋白受体或其他抗性相关蛋白,进而调控植物抗病反应。
4 结论
本研究首先通过转录分析MoIeep1基因在稻瘟病菌野生型菌株Guy11菌丝、孢子和不同侵染时期的表达量,发现其在侵染初期8h表达量最高;信号肽和亚细胞定位分析结果表明MoIEEP1N 端含有17个氨基酸的信号肽,为分泌蛋白且定位于水稻细胞过氧化物酶体或线粒体上;通过构建MoIeep1基因敲除突变体,证明其与稻瘟病菌野生型Guy11相比在生长表型方面无明显差异,但影响其致病性。因此推测该基因可能参与调控稻瘟病菌的致病性。
猜你喜欢
热带作物学报(2020年7期)2020-09-02
农民致富之友(2020年15期)2020-05-25
中国瓜菜(2019年8期)2019-09-19
江苏农业学报(2019年1期)2019-09-10
农民致富之友(2019年24期)2019-08-20
农民致富之友(2018年20期)2018-11-25
山东农业科学(2017年12期)2018-01-09
湖北农业科学(2017年4期)2017-03-28
江苏农业科学(2016年11期)2017-03-21
江苏农业科学(2017年1期)2017-02-27
