
热应激对蛋鸡血清外泌体miRNA表达谱的影响
2021-12-07 01:43黄宇浩崔艳峰黄安琪
中国兽医杂志 2021年7期
黄宇浩 , 高 文 , 崔艳峰 , 黄安琪 , 吴 江 , 康 恺
(广东海洋大学滨海农业学院 , 广东 湛江 524088)
蛋鸡的集约化饲养环境面临严峻的热应激挑战,尤其是我国华南地区,每年高温持续时间长,湿度大。持续高温导致蛋鸡免疫力、采食量、产蛋率和蛋品质下降,严重时还会引起蛋鸡死亡。外泌体是一种可由细胞分泌至细胞外,直径在30~150 nm的膜性囊泡,其组成成分主要有蛋白质(受体、转录因子、酶)、脂质、核酸,核酸成分主要有信使RNA(mRNA)、微小RNA(miRNA)、长链非编码RNA(lncRNA)和环状RNA(circRNA)。目前对外泌体的研究报告比较多,常见于外泌体内含物成分的分析、机体功能异常、病毒感染、各种疾病的分子标记,也作为多种信号的传递体,介导细胞与细胞之间信息交流[1-2],国内对蛋鸡外泌体尤其是蛋鸡在热应激条件下血清外泌体的浓度成分变化等报道少见。
动物在处于热应激状态时,除了相关的应激蛋白、应激激素及细胞因子会发生变化外,miRNA的表达也会出现改变,参与应激反应引起机体生理功能改变[2-3]。Cho等发现,热应激处理后的小鼠结肠癌细胞分泌的外泌体中热休克蛋白70(HSP70)的含量比未经处理的细胞高,而且外泌体能激活树突状细胞和巨噬细胞,触发协同免疫反应,并能增强辅助性T 细胞 1 型免疫应答,清除肿瘤细胞[4]。利用外泌体与miRNA (miR-140)分子联合,通过诱导膜融合,将miRNA释放到骨髓干细胞质中,促进干细胞向软骨细胞的分化,实现外泌体作为生物介质用于疾病治疗[5]。郑月等利用miRNA高通量测序技术在热应激奶牛血清中,发现52个显著差异表达的miRNA,其中通过分析发现miR-181a与免疫功能的调控密切相关,miR-19a和miR-19b与调控热应激的发生有关,这些miRNA可视为诊断热应激的候选生物标记物[6]。Guo等报道热应激通过miRNA 调节Fas/FasL信号通路从而参与热应激诱导的细胞凋亡[7]。另有研究表明,热应激的畜禽通过miRNA 作用,对畜禽生产性能进行调控。Attia等采用基因芯片技术对能量处于负平衡状态奶牛肝脏 miRNA 表达进行检测时发现,miR-17-5p、miR-31、miR-140、miR-1281和miR-2885这5个miRNA表达上调[8]。
本试验通过建立蛋鸡的热应激模型,分别对常温对照组(Normal temperature control group,NC组)及高温控制组(High temperature control group,HC组)进行血清外泌体分离鉴定,并测定血清外泌体浓度变化。利用miRNA 高通量测序技术和荧光实时定量PCR技术,分析差异表达的miRNA,并分析其在热应激和抗热应激过程中所涉及的通路,为研究外泌体miRNA调控蛋鸡热应激反应的机制提供科学依据。
1 材料与方法
1.1 主要试剂 TRIzol,购自天根生化科技(北京)有限公司;外泌体提取试剂盒(76064),购自凯杰生物工程(深圳)有限公司;PrimeScriptTMRT 反转录试剂盒、SYBGREEN Mix,均购自宝生物工程(大连)有限公司。
1.2 主要仪器 冷冻离心机,Sigma公司产品;透射电子显微镜,日本JEM产品;荧光定量PCR仪,美国伯乐产品。
1.3 试验动物 罗曼粉壳蛋鸡鸡苗,购自广东喜迎珍禽有限公司,饲养于广东海洋大学动物医院动物房。蛋鸡育成及饲养管理参照Lohmann Tieraucht Gmb H手册(https://lohmann-breeders.com/)。
1.4 试验方法
1.4.1 试验动物分组 选取40周龄健康状况良好、生产性能相近的罗曼粉壳蛋鸡40只,随机分为两组,常温对照组(NC组)保持鸡舍处于(25±3)℃,高温控制组(HC组)环境温度控制在(32±3)℃,采用空调、除湿机等严格控制环境温湿度,进行饲养试验。3层阶梯笼养,每笼3只鸡。每天投料2次, 上午8:00和下午18:00各1次,自由饮水和采食。光照设置:采用自然光照和人工补光结合的方法,控制16 h光照。饲养持续1个月。
1.4.2 血清的制备与外泌体的提取 NC组和HC组分别取10只蛋鸡,进行采血,每只5 mL,凝血后离心分离血清,混合血清提取外泌体。按照Qiagen外泌体提取试剂盒(76064)说明书步骤进行操作。
1.4.3 透射电镜鉴定外泌体形态 将提取的外泌体用250 μL PBS稀释,用移液器枪头吹打混匀后,取稀释液10 μL滴于载样铜网上,室温静置5 min, 用滤纸将多余液体吸去。用3%磷钨酸钠溶液负染3 min,室温晾干,在透射电镜下观察并拍照(于广东医科大学透射电镜室完成)。
1.4.4 外泌体miRNA 表达谱及粒径检测 将提取的外泌体送杭州联川生物科技有限公司进行miRNA 表达谱检测及粒径分析(Nanoparticle tracking analysis,NTA)。
1.4.5 荧光定量PCR TRIzol裂解外泌体,常规方法提取总RNA,利用PrimeScriptTMRT 试剂盒反转录成 cDNA,并利用 SYBGREEN Mix试剂盒进行RT-PCR 反应,其中5S rRNA基因作为参照(引物信息见表1)。定量PCR体系(20 μL):ddH2O 8 μL、SYBR Premix ExTaqⅡ10 μL、cDNA 1 μL、上下游引物(10 μmol/L)(序列信息见表1)各0.5 μL。反应程序:95 ℃ 5 min,95 ℃ 30 s;退火:50~60 ℃ 30 s;延伸:72 ℃ 30 s,40个循环,72 ℃ 10 min。每个样品3个重复。根据2-ΔΔCt法计算基因在各组中miRNA 的相对表达量。
1.4.6 引物设计与合成 对两组样品中差异表达的8个miRNA,设计引物,由生工生物工程(上海)股份有限公司合成,见表1。

表1 引物序列Table 1 Primer sequences
1.4.7 统计分析 定量PCR数据依据公式2-△△Ct计算,以NC组相对均值为1作图。采用t检验对试验数据进行统计学分析,计量资料以平均值±标准差表示,以P<0.05表示差异显著,P<0.01表示差异极显著。
2 结果
2.1 血清外泌体形态鉴定 在对NC组及HC组蛋鸡进行血清的分离及外泌体的提取后,利用透射电镜对所提取的外泌体进行形态学鉴定。在透射显微镜下观察HC组及NC组样品,视野清晰,可见数个形态呈类球形,直径约为70 nm,外部有双层膜结构,内部有低密度物质的微小囊泡,形态、直径符合外泌体的形态描述。两组样品外泌体形态相似,无明显差别,多数外泌体呈单个分布,见图1。
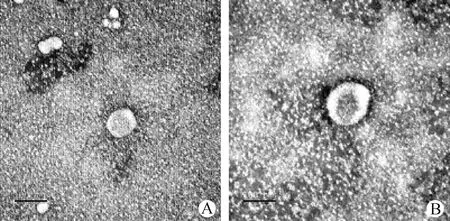
图1 透射电镜下血清外泌体形态Fig.1 Morphology of serum exosomes under transmission electron microscopeA: 常温组(NC组); B:高温控制组(HC组); 标尺:100 nmA: Normal temperature control group (NC group); B: High temperature control group (HC group); Bar:100 nm
2.2 血清外泌体浓度粒径分析 经对血清外泌体NTA分析检测后,发现NC组和HC组大多数外泌体的粒径集中在(70±5)nm,HC组粒径为70 nm的外泌体数量明显多于NC组,HC组外泌体粒径大小范围与NC组外泌体粒径大小范围都在30~150 nm,见图2。对血清外泌体浓度分析时,发现NC组血清外泌体浓度约为1.0×1010个/mL,HC组约为5.5×1010个/mL,见图3。可见单位体积HC组血清外泌体浓度极显著大于NC组(P<0.01)。

图2 血清外泌体富集组分的粒径分布Fig.2 Particle size distribution of serum exosome enriched components

图3 血清外泌体的浓度Fig.3 Concentration of serum exosomes与NC组比较,**:P<0.01Compare with NC group,**:P<0.01
2.3 miRNA表达谱分析及差异表达谱分析 对两组样品miRNA表达谱分析,有287个miRNA在两组样品中均有表达,18个miRNA在NC组中特异表达,39个miRNA在HC组中特异表达,见封二图4。在两组样品中共有的miRNA差异表达分析,表达差异极其显著(P<0.01)的数目为83个,其中HC组较NC组有51个miRNA上调表达、32个miRNA下调表达,见封二图5。上调表达的前5个miRNA:gga-miR-214、gga-miR-1434-3p、gga-miR-1416-5p、gga-miR-1416-5p和gga-miR-29b-2-5p,下调表达的前5个miRNA: gga-miR-6544-5p、gga-miR-19a-3p、gga-miR-6582-3p、gga-miR-142-3p和gga-miR-122-5p,其对应的靶基因主要参与氧化磷酸化、蛋白质转运、丝氨酸和蛋氨酸代谢、脂肪酸代谢。见表2。

图4 常温对照组和高温控制组miRNA测序数据的韦恩图Fig.4 Venn diagrams of miRNA sequencing data in NC group and HC group
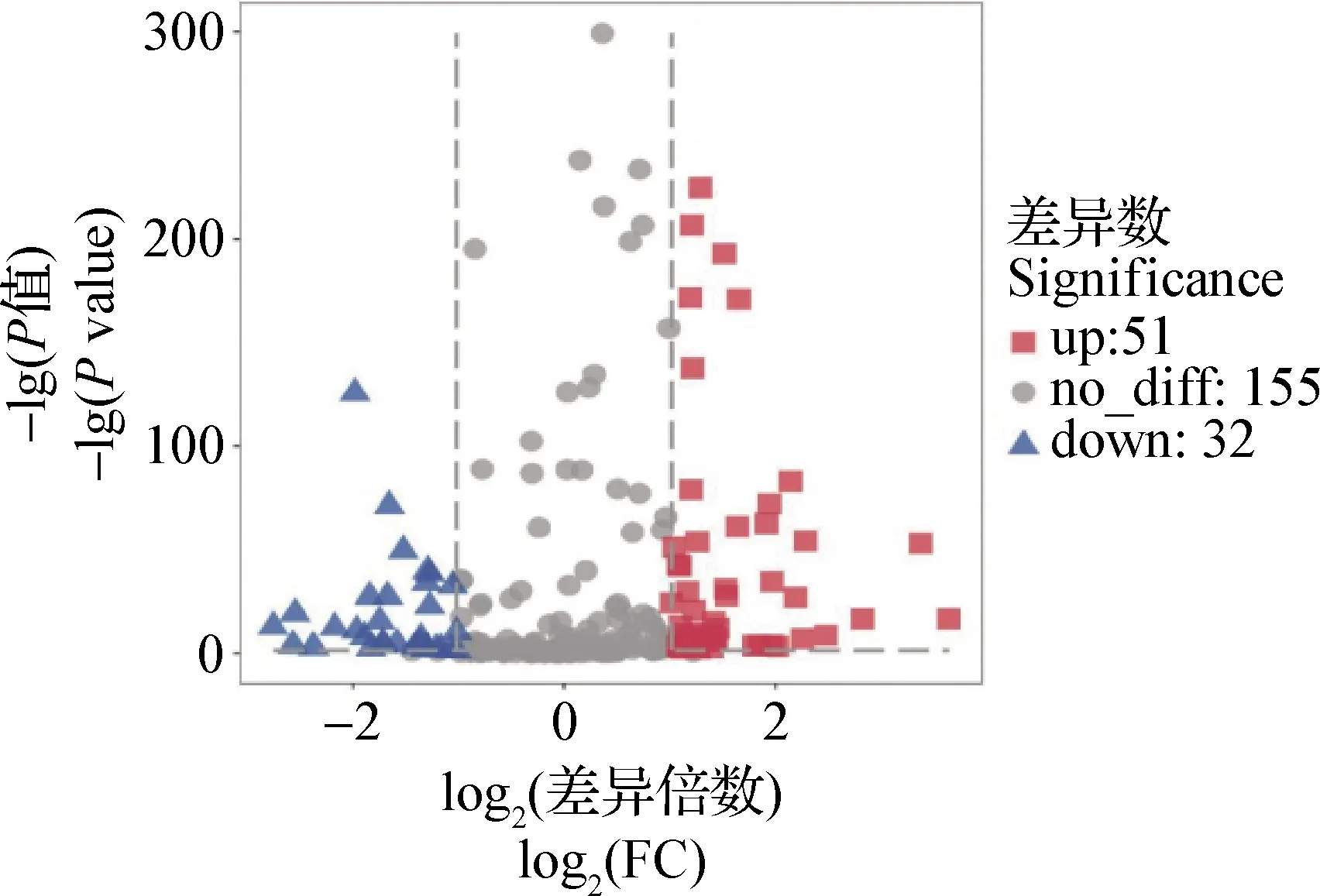
图5 高温控制组对比常温对照组间miRNA差异表达Fig.5 Differential expression of miRNA of HC group compared with NC group

表2 部分差异表达miRNA列表Table 2 List of partial differentially expressed miRNA
2.4 差异表达miRNA靶基因功能预测与分析 对两组样品差异表达miRNA靶基因功能进行预测与分析,在GO数据库中可极显著(P<0.01)富集的生物学过程群(Biological process,BP)有信号转导、氧化还原过程、RNA聚合酶Ⅱ对转录的正调控、转录调控DNA模板、G蛋白偶联受体信号通路、蛋白质磷酸化、蛋白质水解、转录阳性调控DNA模板、RNA聚合酶Ⅱ对转录的负调控、细胞内蛋白质转运、跨膜转运、磷酸化、蛋白质泛素化、RNA聚合酶Ⅱ对转录的调控、细胞增殖的正向调节、凋亡过程的负调控、基因表达的正调控、转录负调控DNA模板、细胞内信号转导、蛋白质转运、离子输运、信号受体活性调控、翻译、囊泡介导的转运、细胞增殖负调控,细胞组分群(Cellular component,CC)有细胞膜、细胞膜组成成分、细胞质、细胞核、胞质、质膜、核质、线粒体、细胞外间隙、内质网、高尔基体、胞外区、质膜整体组分、细胞内膜结合细胞器、核仁,分子功能群(Molecular function,MF)有蛋白质结合、金属离子结合、ATP结合、核苷酸结合、DNA结合、转移酶活性、相同蛋白结合、水解酶活性、蛋白质同聚活性、核酸结合,见封二图6;在KEGG数据库中极显著(P<0.01)富集的通路有泛素介导的蛋白质水解、泛醌等萜类醌生物合成、酪氨酸代谢、蛋白质输出、吞噬体、P53信号通路、氧化磷酸化、鞘糖脂生物合成、叶酸生物合成、铁死亡、脂肪酸降解、内吞作用、半胱氨酸和蛋氨酸代谢、心肌收缩、丁酸代谢、生物素代谢、自噬动物、细胞凋亡、Apelin信号通路、脂肪细胞因子信号通路。见封二图7。

图6 差异表达外泌体miRNA靶基因GO功能聚类分析Fig.6 Gene ontology classifications of target genes of differentially loaded miRNA analyzed in exosomes

图7 差异表达外泌体miRNA靶基因KEGG通道聚类分析Fig.7 Kyoto encyclopedia of genes and genomes classifications of target genes of differentially loaded miRNA analyzed in exosomes
2.5 差异表达miRNA的定量PCR检测 挑选测序结果中显著差异表达的9个miRNA进行定量检测验证,分别是HC组4个miRNA表达上调的gga-miR-221-3p、gga-miR-30a-5p、gga-miR-199-5p和gga-miR-214,HC组5个miRNA表达下调的gga-miR-122-5p、gga-miR-100-5p、gga-miR-125b-5p、gga-miR-142-3p和gga-miR-19b-3p,见图8A。结果表明,4个miRNA上调表达,其中表达差异极显著(P<0.01)有3个(gga-miR-221-3p、gga-miR-30a-5p和gga-miR-214),表达差异显著(P<0.05)有1个(gga-miR-199-5p);5个miRNA下调表达,其中表达差异极显著(P<0.01)有3个(gga-miR-122-5p、gga-miR-142-3p、gga-miR-19b-3p),表达差异显著(P<0.05) 有2个(gga-miR-100-5p和gga-miR-125b-5p),见图8B。结果显示,定量PCR表达差异趋势与miRNA测序结果一致。
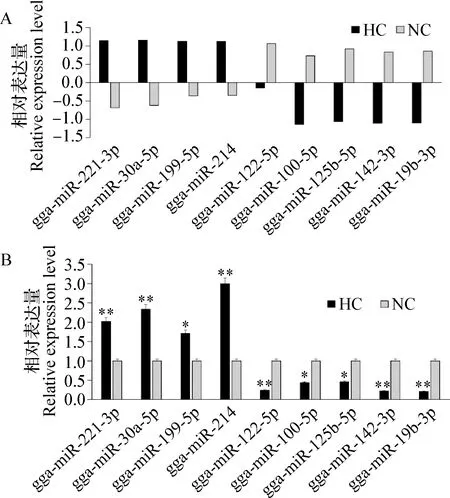
图8 差异表达miRNA的定量PCR检测Fig.8 Detection of differentially expressed miRNA by quantitative PCRA:测序数据分析结果; B:定量PCR检测结果与NC组比较,*:P<0.05, **:P<0.01A:Results of sequencing data analysis; B:Results of qRT-PCR detectionCompare with NC group,*:P<0.05, **:P<0.01
3 讨论
热应激作为最常见的应激因素之一,通过使蛋白质变性进而影响细胞活性及功能,通常体现为蛋鸡生长发育、自身免疫功能、产蛋量及产蛋品质受到影响,对广大蛋鸡养殖产业产生了巨大的影响。血液中存在外泌体,其内涵物会随着机体功能的改变而发生变化,当机体处于热应激状态时,外泌体miRNA的表达也会出现改变。有研究表明,当机体受到应激作用时,会通过不同的途径促进外泌体的释放。大鼠肾近端肾小管细胞处于缺氧状态时,可以显著促进机体细胞产生和分泌外泌体,而这些缺氧状态下产生的外泌体对于大鼠肾小管细胞的损伤具有保护作用[9]。同时,也有研究发现,低氧诱导因子1可以诱导处于缺氧状态的肾小管上皮细胞产生外泌体,通过调节外泌体的含量来调控受体细胞的生理功能[10-11]。祝鑫桃对热应激肝细胞外泌体蛋白质谱分析及其促进程序性死亡作用初探的研究表明,经热应激处理的肝细胞可以产生释放大量的外泌体,引起肝损伤[12]。在本试验中,对外泌体形态及粒径大小分析的结果与上述研究结果相似,形态结构符合外泌体的形态描述,外泌体粒径大小在30~150 nm,说明本试验分离出血清外泌体。对两组样本进行外泌体浓度的测定,发现HC组的血清外泌体浓度(5.5×1010个/mL)大于NC组外泌体浓度(1.0×1010个/mL),说明当机体受到热应激时,会促进外泌体的产生和分泌。
当动物机体受到热应激时,机体的各项生理生化指标都会发生改变,如:免疫指标、抗氧化指标、蛋白质、酶和激素等,从而使机体的免疫功能和抗氧化功能减弱。有研究表明,miRNA对处于热应激动物的细胞增殖、分化、凋亡、免疫功能、抗氧化功能及繁殖性能有重要的调节作用[13]。赵健等阐明NGR 1对动脉粥样硬化的作用的研究中,经过双荧光素酶报告试验证实miR-221-3p是HUVECs中XIST的靶点,miR-221-3p与TRAF 6相互作用,NGR1通过核因子Kappa B(NF-κB)途径调节ox-LDL作用于内皮细胞的增殖、凋亡、炎症反应和氧化应激[14]。傅晓东等证明,miR-30a-5p的表达上调可抑制MAPK/ERK信号从而抑制炎症反应,同时增加SEPN1、TXNL1和GPX1的表达,增强了氧自由基的清除作用,抑制氧化应激[15]。王艳等对miR-214靶向抑制CaMKⅡ调节缺氧复氧诱导H9c2心肌细胞氧化应激的作用的研究中,过度表达的miR-214可抑制CaMKⅡ转录后翻译过程,从而改善H9c2心肌细胞氧化应激水平及凋亡状态[16]。Antonini等研究表明,miR-34a、miR-34b可通过调控凋亡基因(p53)的表达来调控DNA损伤修复、细胞周期阻滞、细胞凋亡[17]。本试验通过对蛋鸡血清外泌体miRNA进行高通量测序以及利用qRT-PCR技术检测验证,发现高温控制组相比于常温对照组,血清中外泌体miRNA 共有83个差异表达,根据上述已发现的研究,说明了本试验中差异表达的miRNA在热应激状态下,受到了激活或抑制,使miRNA表达量上调或下调,对细胞增殖、分化、凋亡、免疫功能有调节作用,产生了一系列抗热应激的生理过程,使蛋鸡适应热应激对机体所产生的影响。
当对两组样品中差异表达miRNA靶基因功能分析时,发现在GO数据库中可极显著富集的生物学过程群有信号转导、氧化还原过程、G蛋白偶联受体信号通路、蛋白质磷酸化、蛋白质水解、磷酸化、蛋白质泛素化、凋亡过程的负调控、信号受体活性调控、翻译、囊泡介导的转运、细胞增殖负调控等;在KEGG数据库中可以显著富集的通路有泛素介导的蛋白质水解、吞噬体、P53信号通路、氧化磷酸化、叶酸生物合成、铁死亡、脂肪酸降解、内吞作用、自噬、细胞凋亡、Apelin信号通路、脂肪细胞因子信号通路。高温环境造成的热应激会引起动物的氧化应激,氧化还原的稳态受到了破坏,使活性氧自由基(ROS)产生过量,导致机体的抗氧化能力下降。与此同时,热应激使活性氧自由基(ROS)过量产生激活了细胞凋亡的内源性途径(即线粒体外膜细胞色素C释放的线粒体途径),当细胞色素C进入细胞质后,激活了起始因子和效应器凋亡蛋白(即半胱氨酸天冬氨酸蛋白酶),使细胞程序性死亡,这证实了热应激能诱导细胞坏死[18]。氧化磷酸化是ATP产生的途径之一,有研究表明,热应激会影响氧化磷酸化过程中的酶活性,从而影响机体对碳水化合物的代谢,葡萄糖是动物主要的功能物质,处于热应激状态的动物血糖水平较正常动物有所下降,处于负能量状态,不能满足动物生长需要[19],热应激会减少脂肪的分解,降低脂肪分解酶的活性,降低游离脂肪酸的水平[20]。有研究表明,热应激能影响细胞膜的流动性和稳定性,从而抑制了细胞内蛋白质运输和蛋白质转运。上述说明热应激动物在热应激与抗热应激过程中,涉及氧化还原过程、半胱氨酸和蛋氨酸代谢、氧化磷酸化、脂肪酸降解、细胞内蛋白质运输、蛋白质转运、蛋白质输出、细胞凋亡等生物学过程,本试验所检测到信息与报道基本一致。
综上所述,热应激会使机体各项生理生化指标发生改变,血清外泌体浓度增加,83个血清外泌体miRNA差异表达,从而使机体的免疫功能和抗氧化功能减弱,且与热应激与抗热应激密切相关的调控通路有氧化还原过程、半胱氨酸和蛋氨酸代谢、氧化磷酸化、脂肪酸降解、细胞内蛋白质运输、蛋白质转运、蛋白质输出、细胞凋亡。本试验的外泌体测序数据为研究热应激对蛋鸡生产性能的影响提供了基础数据,也为外泌体在蛋鸡热应激及抗热应激的研究提供了参考。
猜你喜欢
今日农业(2022年2期)2022-11-16
昆明医科大学学报(2022年2期)2022-03-29
今日农业(2021年7期)2021-11-27
昆明医科大学学报(2021年5期)2021-07-22
老年医学研究(2021年6期)2021-03-09
天津医科大学学报(2019年6期)2019-08-13
国际呼吸杂志(2019年8期)2019-04-29
分析化学(2017年12期)2017-12-25
少儿科学周刊·少年版(2015年11期)2015-12-17
少儿科学周刊·儿童版(2015年11期)2015-12-17
