
放射虫硅质壳腔体微纳米硅质—有机质聚合体赋存特征、成因及油气地质意义
——以四川盆地五峰—龙马溪组为例
2022-01-10 06:57周晓峰李熙喆张晓伟梁萍萍于均民
东北石油大学学报 2021年6期
周晓峰, 郭 伟, 李熙喆, 张晓伟, 梁萍萍, 于均民
( 1. 中国石油大学(北京) 石油工程学院,北京 102249; 2. 中国石油大学(北京) 油气资源与探测国家重点实验室,北京 102249; 3. 中国石油勘探开发研究院,北京 100083 )
0 引言
四川盆地上奥陶统五峰组—下志留统龙马溪组是中国页岩气勘探开发的主力层系,富生物成因硅质页岩是主产气储层,纳米有机孔是页岩储层的主要储集空间[1-2]。页岩的生物成因硅质中,放射虫硅质骨骼化石极为丰富,镜下可以观察到保存较完整的近圆形放射虫硅质壳,指示海洋深水沉积环境和丰富的有机质保存[3-6]。在高分辨率扫描电镜下,观察四川盆地五峰—龙马溪组页岩储层,在放射虫硅质壳腔体内发现交互共生的微米—纳米硅质聚合体(微纳米硅质聚合体)和微米—纳米有机质聚合体(微纳米有机质聚合体),微纳米硅质聚合体以纳米硅质为主,微纳米有机质聚合体以微米有机质居多,二者交互共生聚合形式有多种,有机质发育丰富的纳米孔隙。
岩石中的微纳米硅质聚合体通常称为纳米硅质球粒聚合体。纳米硅质球粒聚合体实际上是微纳米硅质聚合体,硅质微粒不仅具有球形形貌,还常见长条形、不规则多边形等复杂形貌[7-8],采用更具内涵的“微纳米硅质聚合体”代替“纳米硅质球粒聚合体”。微纳米硅质聚合体主要发育于矿物颗粒之间[7,9-11],未见富集于放射虫硅质壳腔体的相关报道。硅质壳为放射虫的硅质骨骼化石,硅质壳腔体由放射虫的腔体(囊体)转化形成[12-14]。硅质壳腔体内交互共生的微纳米硅质聚合体(Micro-nano silica aggregate)和微纳米有机质聚合体(Micro-nano organic matter aggregate)简称为MNSA-MNOMA。
以普通薄片的光学显微镜观察寻找和发现放射虫硅质壳富集的页岩纹层,以高分辨率扫描电镜观察富放射虫硅质壳页岩纹层的氩离子抛光片,分析放射虫硅质壳腔体MNSA-MNOMA赋存特征、成因及油气地质意义,为类似页岩储层及有机孔研究提供指导。
1 地质概况
四川盆地是扬子板块内最富油气的多旋回叠合沉积盆地之一。奥陶纪早期,扬子板块为广阔的陆表海,以碳酸盐沉积为主。至晚奥陶世,受广西运动影响,盆地周缘古陆和隆起抬升,形成“三隆夹一凹”的局限海域深水陆棚沉积体系,产生大面积低能、欠补偿及缺氧的沉积环境,沉积上奥陶统五峰组—下志留统龙马溪组下部约60 m含放射虫和笔石的黑色页岩(见图1),具备形成页岩气的基本地质条件[3-6,15-16]。早志留世中晚期,海平面快速下降,深水陆棚转化为浅水陆棚,陆源黏土质和碎屑颗粒增加,形成黏土质页岩,构成下伏页岩气的优质盖层[17]。
四川盆地五峰—龙马溪组经历晚志留世—石炭纪、晚白垩世—现今的两次大规模抬升和二叠纪—早白垩世的大幅度沉降[18]。晚志留世—石炭纪构造抬升使五峰—龙马溪组下部黑色页岩长期处于浅埋藏状态,二叠纪—早白垩世持续强烈沉降使五峰—龙马溪组的最大埋深超过6.5 km,有机质达到过成熟阶段,页岩气藏形成。晚白垩世以来多阶段隆升过程普遍具有1.0~4.0 km的地表剥蚀厚度,现今五峰—龙马溪组页岩气埋深一般为3.0~5.5 km。
2 样品选择与研究方法
2.1 样品选择
四川盆地五峰—龙马溪组发育黑色富生物硅质页岩,放射虫硅质壳是生物硅质的主要赋存方式之一[3-6]。硅质壳的粒径普遍小于150 μm,岩心观察很难被发现。多口页岩气探井取心段密集采样,样品分为两部分:一部分样品用于制作普通薄片寻找放射虫硅质壳;另一部分样品备用制作氩离子抛光片,观察硅质壳腔体内MNSA-MNOMA赋存特征。
偏光显微镜观察普通薄片,放射虫硅质壳通常富集成纹层,约为8层/m,厚度普遍不足5 mm;壳体为黑色背景下格外醒目的亮色环带,宽度一般为10~25 μm;壳的腔体显示黑色、黄色和污浊的白色等,直径分布在40~120 μm之间(见图2)。确定放射虫硅质壳纹层样品后,通过放大镜在备用样品中圈定硅质壳纹层的具体位置(黑色背景下密集的白色小斑点)。

图2 四川盆地五峰—龙马溪组放射虫硅质壳纹层典型偏光显微镜照片Fig.2 Typical polarizing microscope photographs of radiolarian siliceous shell laminae from Wufeng-Longmaxi Formation in Sichuan Basin
选取4块典型硅质壳纹层样品制作氩离子抛光片,利用高分辨率扫描电镜观察和分析MNSA-MNOMA赋存特征。4块样品选自四川盆地南部深层页岩气开发试验区,其中2块样品取自Z205井五峰组顶部,样品编号及深度分别为Z205-26、4 103.37 m和Z205-27、4 103.96 m(见图1(b));另外2块样品取自L205井龙马溪组底部,样品编号及深度分别为L205-8、4 031.23 m和L205-9、4 033.40 m(见图1(c))。偏光显微镜观察普通薄片在中国石油大学油气资源与探测国家重点实验室完成,偏光显微镜型号:Leica DM 4500。氩离子抛光页岩样品和扫描电镜观察在中国科学院地质与地球物理研究所微纳结构和岩心分析实验室完成,氩离子抛光仪型号:Fischione Model 1060,扫描电镜型号:Zeiss Crossbeam 550。
2.2 研究方法
采用MAPS(Modular Automated Processing System)扫描技术,获取高精度、大视域微观图像,研究MNSA-MNOMA赋存特征。将氩离子抛光样品划分为一系列规整网格,对每一网格进行扫描成像,获得二次电子扫描图像,对所有网格的图像进行拼接,得到一幅二维大视域扫描图像[19]。首先,采用500 nm(放大500倍)较低分辨率快速对整个氩离子抛光片(2 mm×2 mm)进行扫描成像,获取放射虫硅质壳纹层的扫描图像数据体。然后,选取典型放射虫硅质壳获取10 nm(放大2.64×104倍)或4 nm(放大6.55×104倍)高分辨率扫描图像数据体。最后,采用离线版图像编辑器(ATLASTM BROWSER-BASED VIEWER)对不同分辨率图像数据体进行处理,获取更多、更加具有代表性的图像,如宽×高为5 mm×5 mm、分辨率为500 nm的图像数据体能够获取分辨率为7.5 μm~500 nm的图像;宽×高为150 μm×130 μm、分辨率为4 nm的图像数据体能够获取分辨率为157~4 nm的图像,不同分辨率的图像数据体能够获取MNSA-MNOMA微域的连续缩放倍数图像(见图3)。

图3 四川盆地五峰—龙马溪组放射虫硅质壳纹层典型扫描电镜照片Fig.3 Typical scanning electron microscope photographs of radiolarian siliceous shell laminae from Wufeng-Longmaxi Formation in Sichuan Basin
氩离子抛光岩面垂直于岩心样品的层理面。镜下观察氩离子抛光岩样时,以层理面或笔石条带在视域中显示水平状态作为基本条件,尽管难以确定样品的顶、底面是否为岩石地下状态的顶、底面,但是大致可以保证样品保持地下岩层的水平状态,能够判断岩石对上覆地层压实作用响应的微观信息。
3 MNSA-MNOMA赋存特征
放射虫硅质壳腔体内普遍发育MNSA-MNOMA,交互共生的MNSA和MNOMA呈现此消彼长的现象。若MNSA含量低,则MNOMA含量高,MNSA镶嵌在连片分布的MNOMA内(见图4);若MNSA含量高,则MNOMA含量低,MNOMA点缀在MNSA内(见图3(c))。MNOMA发育纳米孔隙,形状相似,孔径相近,分布均匀(见图3-7)。
根据MNSA和MNOMA的赋存状态,可以把MNSA-MNOMA划分为5种类型,Ⅰ类为MNSA漂浮在MNOMA内,Ⅱ类为MNSA均匀混合MNOMA,Ⅲ类为MNOMA漂浮在MNSA内,Ⅳ类为MNOMA充填镶嵌状MNSA缝隙,Ⅴ类为MNSA-MNOMA弧形带。通常情况下,Ⅰ~Ⅳ类聚合体可以分别独自占据一个硅质壳腔体,Ⅴ类聚合体和Ⅰ~Ⅳ类聚合体共处于一个硅质壳腔体(见图4-7)。
在Ⅰ和Ⅴ类聚合体共同出现的硅质壳腔体内,Ⅴ类聚合体通常将Ⅰ类聚合体分割成数个更加微小的区域(见图4(a),黄色虚线指示Ⅴ类聚合体)。各个Ⅰ类聚合体微区中,硅质微粒星点状散落在连片分布的有机质内,硅质微粒普遍小于600 nm,偶尔可见粒径不足2 μm的微米级颗粒(见图4(b-e))。Ⅴ类聚合体弧形带微区中,多个纳米硅质微粒首尾连接成微米硅质条带,长度一般不足2 μm,延伸方向大致垂直于弧形带切线方向,有机质也呈条带状并与硅质条带平行(见图4(f))。硅质壳顶部有一个约为56.86 μm的缺口,缺口处充填云母蚀变形成的黏土矿物条带(见图4(a、g))。紧邻缺口有556.10 nm~4.10 μm宽度的Ⅰ类聚合体微区异常带,具体表现为MNOMA中纳米孔隙不发育(见图4(g)黄色虚线与缺口共同限定的范围),其他微区MNOMA中普遍发育纳米孔隙(见图4(e)黄色虚线以下微区和图4(b-f))。根据这种异常现象推断,缺口处抗压实能力弱导致有机质中缺乏纳米孔隙,其他微区富有机孔隙归结为硅质壳具有较强的抗压实能力。
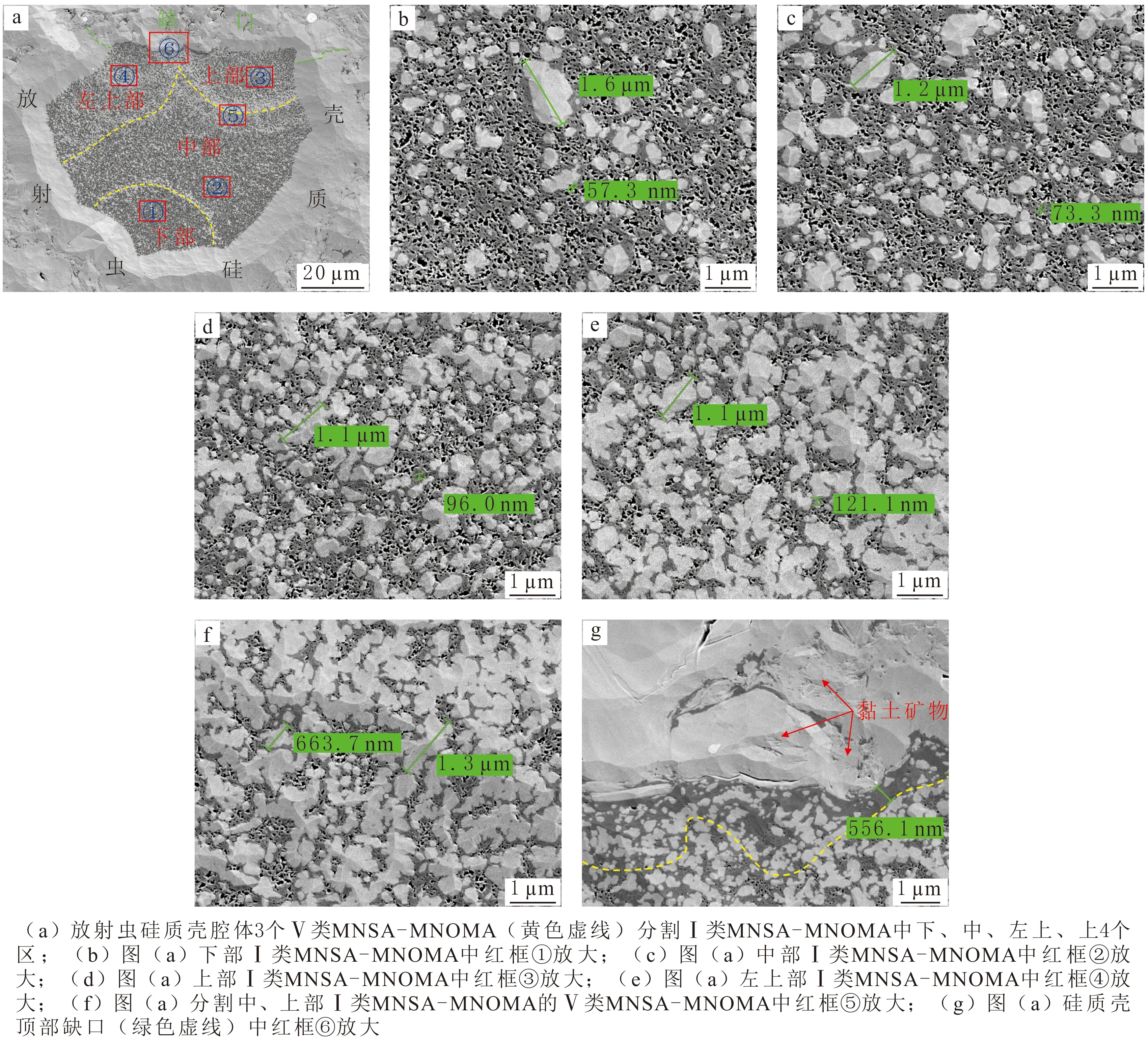
图4 Ⅰ和Ⅴ类MNSA-MNOMA典型微观照片Fig.4 Classic microscopic images of type Ⅰ and type Ⅴ MNSA-MNOMA
Ⅱ类聚合体可与黄铁矿晶体共处于硅质壳腔体(见图5(a))。微纳米硅质和有机质混合均匀,二者占据大致等体积的硅质壳腔体空间,硅质以纳米级微粒为主,有机质中纳米孔丰富(见图5(b))。黄铁矿晶体长度为44.5 μm、宽度为33.5 μm,晶形好(五边形),晶棱平直,晶面光滑,除局部与硅质壳接触,总体被Ⅱ类聚合体包围,表明黄铁矿晶体的形成时间早于Ⅱ类聚合体。

图5 Ⅱ类MNSA-MNOMA典型微观照片Fig.5 Classic microscopic images of type Ⅱ MNSA-MNOMA
Ⅲ和Ⅴ类聚合体可以共存在一个硅质壳腔体内(见图6(a))。Ⅲ类聚合体发育区内,硅质微粒相互接触组成“硅质网”,有机质分布在“网眼”中;小于2 μm的长条状微米硅质较多,延伸方向多变(见图6(b)红色虚线)。Ⅴ类聚合体发育区内,有5个相互平行的弧形带(见图6(a、c)黄色虚线),每个弧形带主要由2 μm左右的长条状微米硅质组成,微米硅质有一定的优势延伸方向,以垂直圆弧切线方向居多(见图6(a、c)红色虚线),各弧形带之间有机质含量增加。

图6 Ⅲ和Ⅴ类MNSA-MNOMA典型微观照片Fig.6 Classic microscopic images of type Ⅲ and type Ⅴ MNSA-MNOMA
Ⅳ类聚合体中,MNSA具有镶嵌结构,单个微粒体难以识别,有机质“塞牙缝式”充填硅质微粒之间的孔隙(见图7)。在没有充填有机质的纳米孔中,总能看到硅质微粒的自由表面呈尖锥状(见图7(b)红色箭头所指),预示硅质微粒向石英晶粒发展。五峰—龙马溪组曾经埋深至6.0 km以下,上覆地层压力巨大,致密的镶嵌结构可能与硅质微粒的压溶作用密切相关,纳米孔也可能是压溶作用的产物。

图7 Ⅳ类MNSA-MNOMA典型微观照片Fig.7 Classic microscopic images of type Ⅳ MNSA-MNOMA
4 MNSA-MNOMA赋存成因
4.1 形成时序
放射虫是浮游微体古生物的一个重要类群,从寒武纪至今,在海洋环境中繁盛。现代放射虫在生物学分类上包含多囊虫类、稀孔虫类和等辐骨虫类,其中多囊虫类的骨骼由硅质组成,稀孔虫类为有机质与硅质的混合物或碳酸盐,等辐骨虫类是以一种与几丁质有关的特殊有机质(硫酸锶)为组分[13-14]。放射虫死亡后,能够沉降到海底并埋藏保存为化石的仅为多囊虫类,难溶的硅质骨骼演化为硅质壳,囊内的细胞质分解后转化为硅质壳的体腔。五峰—龙马溪组硅质壳应为多囊虫类放射虫的硅质骨骼石化而来,硅质壳腔体为细胞质分解后的囊体。这说明MNSA-MNOMA并不是放射虫的生物组构,而是后期进入硅质壳腔体内的微纳米硅质和有机质。
Ⅰ类聚合体中硅质和有机质微粒的赋存状态表明,有机质的形成时间不可能晚于硅质微粒的;若有机质的形成时间晚于硅质微粒的,则硅质微粒在重力作用下降落到硅质壳腔体底部聚集起来,与硅质微粒悬浮在有机质中的实际情况不符。Ⅳ类聚合体中硅质和有机质微粒的赋存状态指示,硅质微粒的形成时间不可能晚于有机质的;若硅质微粒的形成时间晚于有机质的,则分散有机质微粒在重力作用下降落至硅质壳底部而形成絮凝状有机质团块,与有机质微粒散落在镶嵌状硅质中的实际情况不符。由Ⅰ和Ⅳ类MNSA-MNOMA赋存状态的分析结果可知,MNSA和MNOMA同时出现在放射虫腔体内是对这些微观现象的最合理解释。
Ⅰ~Ⅳ类聚合体的差异仅能指示不同硅质壳腔体内微环境的差异,微环境中有机质含量高形成Ⅰ和Ⅱ类聚合体,硅质含量高形成Ⅲ和Ⅳ类聚合体。Ⅴ类聚合体可能是Ⅰ~Ⅳ类聚合体在埋藏成岩过程中发生差异压实作用的产物,承载上覆压力大的区域,硅质微粒发生旋转或垂直应力方向压溶,长轴调整至垂直于压应力的方向而达到稳定状态保持下来,有机质发生塑性变形被挤压至硅质颗粒密集弧形带的两侧,体现在Ⅴ类聚合体中长条状硅质微粒的延伸方向垂直于弧形带切线(见图4(d)、图6(c));承载上覆压力小的区域,硅质微粒密度小,长条状硅质微粒也无优势排列方位(见图4(b-c)、图6(b))。
4.2 形成时间
放射虫是生活在海洋表层的浮游生物,从死亡至保存为目前的硅质壳大致经历5个成岩阶段,依次为水柱中沉降—成岩阶段、同生成岩阶段、埋藏早期成岩阶段、埋藏中期成岩阶段和埋藏晚期成岩阶段。同生成岩阶段是指沉积物沉降至水底到与海水隔绝阶段,经典成岩现象为富有机质沉积物中莓状黄铁矿大量出现[20-22]。埋藏早期成岩阶段是指同生成岩阶段之后、有机质生油之前,显著成岩特征为压实作用。埋藏中期和晚期成岩阶段的时间分别对应页岩油生成阶段和页岩气生成阶段。
硅质壳缺口处黏土矿物化云母碎屑未能进入硅质壳腔体,说明MNSA-MNOMA的形成时间早于埋藏早期成岩阶段的压实作用过程。MNSA-MNOMA早于压实作用充填硅质壳腔体,增加硅质壳的抗压实强度,硅质壳才能在后来的压实过程中不被压垮和破碎。
硅质壳腔体内没有发现莓状黄铁矿,可以解释为同生成岩阶段之前MNSA-MNOMA已经完全占据硅质壳腔体。同生成岩阶段为适宜莓状黄铁矿沉淀时期,硅质壳之外的空间沉淀大量莓状黄铁矿,硅质壳腔体全部充填MNSA-MNOMA而不能为莓状黄铁矿沉淀提供空间,因此无法沉淀莓状黄铁矿。
Ⅰ~Ⅳ类聚合体形成的成岩阶段为水柱中沉降—成岩阶段,可以解释镜下观察的各种微观现象,放射虫硅质壳腔体内形成各类MNSA-MNOMA,同生成岩阶段MNSA-MNOMA阻止莓状黄铁矿在硅质壳腔体内沉淀,埋藏早期成岩阶段MNSA-MNOMA抵抗云母、碳酸盐矿物等从硅质壳缺口处进入腔体。
4.3 物质来源
深水页岩中微纳米硅质可能有3个物质来源:生物硅的溶解、微生物(细菌)活动、黏土矿物转化,微纳米硅质聚合体有机质可能来自微生物活动[7-14]。
硅藻、放射虫、海绵骨针具有硅质壳(骨骼),是海水中的潜在硅源。在矿物学上,这些生物的硅质骨骼称为蛋白石A,是由胶态SiO2、硅石和少量水组成的非晶质矿物[12-14]。活体生物的硅质骨骼外部包裹有机膜,有效降低蛋白石A在海水中的溶解度。生物死亡后,有机膜在几天内分解[23],硅质壳裸露在海水中,溶解速率增加[24]。生物硅的溶解性能与海水温度密切相关,溶解速率随水温上升而急剧上升,溶解度可从3 ℃时800~1 000 μM Si增加至23 ℃时1 500~1 700 μM Si[25-26]。生物硅的溶解性能还与环境压力、硅酸盐浓度、生物硅基质中痕量元素,以及生物硅比表面积有关。表层海水中产出的生物蛋白石的50%~60%可以在100 m水深内溶解,生物硅在沉降过程中发生的最显著变化是溶解[27]。晚奥陶世五峰组沉积期—早志留世龙马溪组沉积期,四川盆地位于赤道附近[28],海水温度高,水深约为200 m[29],有利条件促进生物硅质溶解并为MNSA-MNOMA提供硅源。
微生物(细菌)活动为微纳米硅质沉淀提供物质来源的研究成果较多,研究对象除南阿曼盐盆地前寒武纪晚期—早寒武世深海沉积的“无机”硅质岩外,鲜有文献提及微生物对显生宙深海沉积的富有机质岩石的影响[7]。下寒武统硅质岩的形成时间早于放射虫的出现时间,前寒武纪晚期—早寒武世的深海沉积微纳米硅质是微生物活动的结果;在早寒武世晚期的页岩中发现放射虫和海绵骨针,富含蛋白石A的微体古生物为硅质沉淀提供一个明显的硅源。STOLPER D A等[30]认为,南阿曼盐盆地古老硅质岩中的微纳米硅质和有机质起源于海水,微生物与微纳米硅质共生共存。这预示细菌活动可能为四川盆地五峰—龙马溪组MNSA-MNOMA提供主要物质来源。
蒙皂石向伊利石转化释放硅质是第三个硅源,主要发生在地层埋深大于2.0 km、温度超过60 ℃的埋藏中期成岩阶段[31-32]。研究区MNSA-MNOMA的形成时间为水柱中沉降—成岩阶段,早于蒙皂石伊利石化释放硅质的成岩时间,可以忽略蒙皂石伊利石化给MNSA-MNOMA的形成提供硅质源。
生物硅溶解和微生物(细菌)活动是微纳米硅质的物质来源,但微生物活动是MNOMA的唯一物质来源。研究区页岩中放射虫硅质壳腔体内MNSA-MNOMA的主要物质来源于微生物(细菌)活动,生物硅的溶解提供部分硅源。
MNSA-MNOMA通过微生物(细菌)活动普遍出现在从海水表层向海底降落的放射虫硅质壳腔体内,在硅质壳外的海水中没有形成MNSA-MNOMA,是由硅质壳腔体的内外环境差异引起的。硅质壳腔体可以看作相对封闭系统,水体清洁为微生物繁盛提供适宜的环境,有利于MNSA-MNOMA沉淀和聚集。硅质壳腔体外的海水中,陆源黏土矿物含量较高[33],抑制微生物活动和/或提高硅质沉淀的浓度,导致缺乏MNSA-MNOMA,进而在页岩粒间孔隙中难以观察到MNSA-MNOMA。即使是很低的黏土矿物含量,也能极大延缓蛋白石微粒的形成[34]。硅质沉积物中,在黏土含量极低的条件下可以见到丰富的微纳米硅质[35-36]。
5 油气地质意义
5.1 有机孔归属
有机质内的蜂窝状纳米孔隙是页岩油气的重要储集空间,是干酪根孔(原地有机质孔)还是石油固体沥青化或焦沥青化孔(迁移有机质孔)存在分歧,分析放射虫硅质壳腔体内MNSA-MNOMA成因,可以解决有机孔的归属问题。
LOUCKS R G等[37]利用扫描电镜观察氩离子抛光页岩的有机孔产状,有机孔发育于有机质成熟度达到成熟—过成熟阶段的干酪根内部,未成熟有机质内部缺乏孔隙,有机孔是干酪根热降解形成油气的伴生产物。BERNARD S等[38]利用同步扫描透射X线显微镜,结合氩离子抛光技术进行页岩孔隙研究,认为有机孔是石油热裂解的产物,分布于石油固体沥青化或焦沥青化微粒,干酪根中孔隙少见,有机孔发育在迁移有机质中。根据有机质的赋存特征及与其他矿物的接触关系,LOUCKS R G等[39]将有机质划分为原地有机质和迁移有机质,认为有机孔发育于迁移有机质内部。关于四川盆地五峰—龙马溪组页岩有机孔的归属认识也存在分歧,采用有机质热模拟方法研究页岩油气的成果资料倾向于有机孔为干酪根孔[40-41],采用氩离子抛光和扫描电镜方法研究页岩岩石学特征的成果资料,倾向于有机孔为石油固体沥青化或焦沥青化微粒内孔[42-43]。
根据放射虫硅质壳腔体内MNSA-MNOMA赋存状态特征及成因,微纳米硅质和有机质形成于硅质壳从海水表层沉降至海底的过程,有机质来源于微生物活动,MNOMA是原地有机质聚合体,有机孔属于干酪根孔。
5.2 成岩作用研究
LOUCKS R G等[37,39]建立页岩储层成岩作用序列的概念模型,将页岩储层的成岩阶段划分为同生成岩阶段、埋藏早期成岩阶段、埋藏中期成岩阶段、埋藏晚期成岩阶段。同生成岩阶段,原始沉积碎屑颗粒之间孔隙发育,生物硅质壳腔体内无填隙物,莓状黄铁矿大量沉淀。埋藏早期成岩阶段,压实作用强,原始粒间孔隙的体积剧烈减小和数量骤然减少,生物硅质壳腔体部分充填自生矿物。埋藏中期成岩阶段,石油充填原始粒间孔隙和生物硅质壳腔体内孔隙并形成有机孔。埋藏晚期成岩阶段,石油裂解为天然气并生成有机孔。因此,四川盆地五峰—龙马溪组放射虫硅质壳腔体内的MNSA形成于埋藏早期成岩阶段,MNOMA是埋藏中期成岩阶段进入MNSA内的石油,在埋藏晚期成岩阶段固体沥青化和焦沥青化的伴生产物,MNOMA的纳米孔隙是迁移有机质孔。
王秀平等[44]将四川盆地及邻区五峰—龙马溪组页岩储层成岩作用阶段划分为同生成岩阶段、埋藏早期成岩阶段、埋藏中期成岩阶段、埋藏晚期成岩阶段;赵迪斐等[45]将成岩作用阶段划分为埋藏早期成岩阶段、埋藏中期成岩阶段、埋藏晚期成岩阶段。无论是四分还是三分成岩阶段强调成岩作用对孔隙发育的影响,指出压实和胶结作用是破坏性成岩作用,溶蚀作用是建设性成岩作用,没有采用典型岩石学特征开展成岩作用序列研究。
成岩作用序列研究是开展储层孔隙类型、微观孔隙结构演化研究的前提条件。对于页岩油气,更加客观的成岩作用序列能够确定有机孔是原地有机质孔还是迁移有机质孔,有机孔归属不同,油气勘探方向存在差异,有机孔为原地有机质孔的勘探方向是寻找原生有机质发育的页岩,有机孔为迁移有机质孔的勘探方向是发现原生粒间孔发育的页岩。页岩成岩作用序列研究[37-39,44-45]缺乏典型岩石学证据作为第一手资料,导致有机孔归属多解性,进而影响油气勘探效果。
考虑放射虫硅质壳腔体MNSA-MNOMA典型赋存特征,应将岩石学证据作为研究页岩储层的第一手资料,挖掘岩石学特征蕴含的成岩信息,同时关注水柱中沉降—成岩阶段。以往认为埋藏早期成岩阶段硅质壳腔体内才开始部分填充自生矿物[3,16],导致成岩作用序列划分产生分歧:
(1)同生成岩阶段,大量沉淀在粒间孔隙中的莓状黄铁矿没有出现在硅质壳腔体内,甚至是残缺不全硅质壳体腔内;
(2)局部残缺不全硅质壳的体腔和海水是连通的,没有陆源石英、长石、云母及黏土矿物沉淀其中;
(3)硅质壳没有在上覆地层压力下破碎;
(4)局部残缺不全硅质壳体腔内没有挤入泥质沉积物。
如果MNSA-MNOMA形成于硅质壳水柱中沉降—成岩阶段可以解释分歧,MNSA-MNOMA早已全充填放射虫硅质壳腔体,不再有陆源碎屑、莓状黄铁矿沉淀及泥质沉积物进入的空间,尤其是MNSA的支撑增强硅质壳抵抗上覆地层压力。
6 结论
(1)四川盆地五峰—龙马溪组放射虫硅质壳富集成纹层,硅质壳腔体内发育5种类型(Ⅰ~Ⅴ类)交互共生的MNSA-MNOMA,MNOMA有丰富的纳米孔。
(2)放射虫硅质壳腔体内MNSA-MNOMA同生共存,形成时间为水柱中沉降—成岩阶段,所需硅质和有机质主要由微生物供给,生物硅溶解贡献部分硅源;Ⅰ~Ⅳ类MNSA-MNOMA赋存状态指示不同硅质壳腔体内微环境差异,Ⅴ类MNSA-MNOMA是Ⅰ~Ⅳ类MNSA-MNOMA经历差异压实作用的产物。
(3)MNSA-MNOMA赋存状态的典型图像是研究赋存成因及有机孔归属的基础,应将典型岩石学特征作为研究页岩储层的第一手资料,重视沉积物在水柱中沉降—成岩阶段的变化。
猜你喜欢
盐科学与化工(2022年9期)2022-10-10
宝藏(2022年1期)2022-08-01
城市轨道交通研究(2022年6期)2022-07-15
环球时报(2022-04-18)2022-04-18
中国新通信(2020年3期)2020-07-06
中学生数理化·高三版(2016年11期)2017-03-02
海外文摘·艺术(2017年15期)2017-02-01
中学生数理化·高三版(2016年2期)2016-09-10
中学生数理化·高一版(2008年5期)2008-11-15
幼儿时代·故事妈妈(2004年5期)2004-08-25
