
虾肝肠胞虫孢壁蛋白SWP7的多克隆抗体制备和定位分析
2022-02-11 13:05包杰宁梓健丰程程姜宏波
中国渔业质量与标准 2022年6期
包杰,宁梓健,丰程程,姜宏波,2*
(1. 沈阳农业大学动物科学与医学学院,辽宁 沈阳 110866;2.东北畜禽疫病研究教育部重点实验室,辽宁 沈阳 110866)
微孢子虫是一类高度进化的、专性营细胞内寄生的单细胞真核生物,普遍存在于自然环境中,宿主类型多样,可感染包括人在内的所有动物[1-3]。目前为止,已有约200个属,1 500余种微孢子虫被报道[4-6]。经过对微孢子虫150多年的探究,科研人员发现其特征具有独特性:裂殖增殖期过渡至孢子形成期,孢子形态产生差异;在细胞水平上,缺乏过氧化物酶体和典型的高尔基体结构[7-8];已进化出极管和孢壁参与的独特侵染机制[3]。微孢子虫游离于胞外环境时,其孢壁蛋白可能与宿主细胞上的信号分子识别互作,触发极管弹出以刺破宿主细胞膜,随之迅速将含有遗传物质的孢原质注入宿主细胞,开始胞内寄生阶段[8-10]。鉴于孢壁在入侵装置中的重要作用,研究其组成对阐明宿主与寄生虫相互作用的机制具有重要意义。
微孢子虫的孢壁由3部分组成,分别为致密的外壁、较厚的电子透明内壁和包围孢原质的质膜。多项研究表明,孢壁中包含许多孢壁蛋白(spore wall protein,SWP),其为孢壁提供结构支撑,保护孢原质免受环境压迫的同时,也参与到信号传递、粘附等重要的生理过程中。在兔脑炎微孢子虫(Encephalitozooncuniculi)、肠脑炎微孢子虫(Encephalitozoonintestinealis)的内壁和外壁,已成功地定位到几种孢壁蛋白[11-12]。此外,一些研究人员还发现孢壁蛋白可通过宿主细胞表面GAG粘附在宿主细胞上[13-14]。
虾肝肠胞虫(Enterocytozoonhepatopenaei,EHP)2001年首次发现于日本囊对虾(Penaeusjaponicus)的肝胰腺上皮细胞内[15]。直到2009年,Tourtip等[16]通过显微观察和分子检测等技术手段将其命名。EHP极具传染性,目前世界上主要的凡纳滨对虾(Litopenaeusvannamei)养殖地区都有虾肝肠胞虫病的报道,给对虾养殖业造成巨大损失。因此,探究EHP感染机制对该病的预防与治疗具有指导性意义。微孢子虫的所有成员都有一种独特的、高度专业化的入侵机制,涉及极管和孢壁,但关于EHP孢壁结构的研究较少。因此,本研究以EHP全基因组中鉴定到的孢壁蛋白SWP7为研究对象,首先通过生物信息学方法对其序列基本特征进行分析,然后合成该基因序列、构建原核表达载体和表达纯化蛋白,最后制备多克隆抗体。通过Western blotting 验证该抗体的特异性后,利用间接免疫荧光技术观察和分析该蛋白在成熟孢子上的定位情况,为深入研究EHP孢壁结构和入侵机制提供基础。
1 材料与方法
1.1 实验材料
虾肝肠胞虫样品由本实验室保存,新西兰白兔购自重庆市第三军医大学。实验所需试剂有:Acr、Bis、Tris和弗氏佐剂(Sigma);6xHis标签蛋白纯化和BCA蛋白浓度测定试剂盒(ProbeGene);Percoll细胞分离液(鼎国生物);Alexa Fluor 488标记的山羊抗兔IgG(生工生物)。
1.2 SWP7基因序列的生物信息学分析
从Genbank下载获得编码为OQS55031.1_1的SWP7蛋白氨基酸序列;利用ExPASy预测和分析SWP7氨基酸组成、分子式、等电点等理化特征;SignalP-4.0用于分析SWP的信号肽;KohGPI筛选锚定位点;使用TMHMM Server v.2.0预测跨膜域;用NetOGlyc3.1Server和NetNGlyc 1.0 Server预测和分析O-糖基化和N-糖基化位点;利用NetPhos 2.0 Server预测磷酸化位点;通过ProtScale分析SWP7的疏水性;应用SMART预测SWP7的功能结构域。
1.3 SWP7基因克隆
根据虾肝肠胞虫SWP7序列信息、His标签以及酶切位点位置,设计载体构建方案见图1,基因合成这一序列。通过双酶切胶回收PCR扩增片段以获得目的片段;pET28a载体采用NcoI/XhoI 双酶切线性化后通过胶回收获得载体片段;再将上述目的DNA片段连接至载体中。连接反应条件为22.0 ℃,1 h;体系为载体片段4.0 μL,目的基因片段8.0 μL,10×T4 Ligase buffer 2.0 μL,T4 DNA Ligase 1.0 μL,加ddH2O至20.0 μL。将连接反应产物转化至TOP10感受态菌株中,涂布于含有抗生素的LB平板进行培养。挑取单克隆菌落稀释100倍后作为DNA模板,采用pET载体通用引物(T7)进行PCR扩增,选取阳性菌落扩培后,提取其质粒进行测序验证。
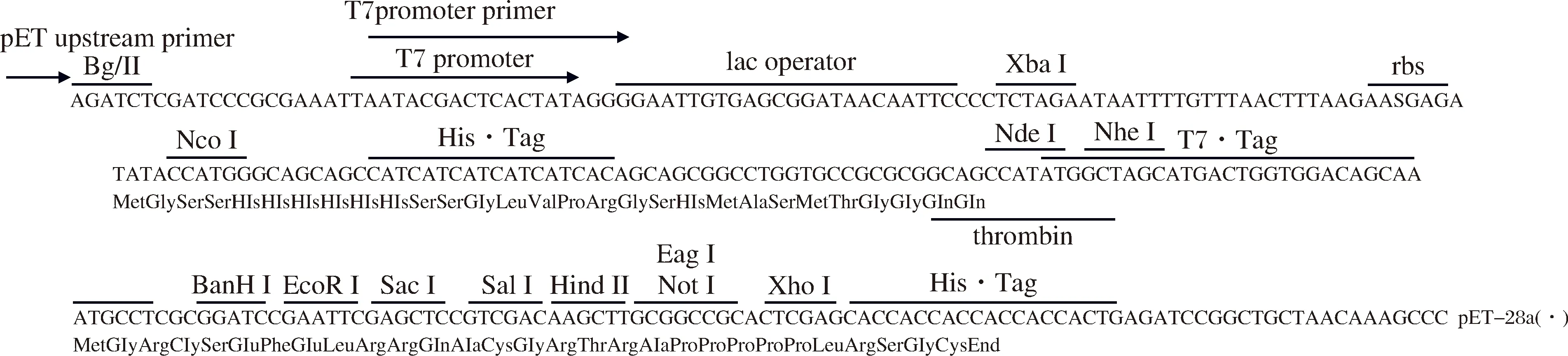
图1 SWP7序列载体构建方案Fig.1The vector construction scheme of SWP7 sequence
1.4 SWP7蛋白的诱导表达与纯化
将阳性单克隆菌落接种至无菌的LB液体培养基(含50 mg/L的卡纳霉素),37.0 ℃培养至OD600约为0.6时,加入终浓度为0.5 mmol/L的IPTG诱导pET28a-SWP7表达,2 h后将培养基离心获得菌体,利用Western Blot检测蛋白表达情况。用Ni-IDA柱纯化以上收集到的菌体蛋白。
1.5 SWP7多克隆抗体的制备和Western Blot分析
将纯化后的SWP7重组蛋白与弗氏佐剂按1∶1混合均匀,至呈乳化状态,通过皮下免疫对2只新西兰白兔(2.0~2.5 kg)注射4次,每次400.0 μg。第1次免疫注射液为重组蛋白与完全弗氏佐剂的混合液。第1次免疫后每隔2~3周通过重组蛋白与不完全弗氏佐剂混合进行强化免疫。免疫过程中,通过间接ELISA方法监测抗血清针对SWP7的效价,当效价大于1∶50 000时收集抗体。
取适量处理后的蛋白样品在12% SDS-PAGE凝胶中进行电泳分离,然后转移到PVDF膜上,再用牛血清白蛋白进行封闭,用稀释好的一抗孵育,洗涤,随后用稀释好的二抗孵育,再次洗涤,最后曝光和显色。
1.6 间接免疫荧光实验
将EHP样品均匀涂在粘附性载玻片上,室温过夜;4%多聚甲醛固定后用PBS缓冲液冲洗;1%Triton X-100透明处理(无菌操作)后用PBS缓冲液冲洗;然后用5%牛血清白蛋白封闭;在载玻片上均匀滴加用封闭液稀释100倍的anti-SWP7,轻轻盖上一层封口膜(无气泡),4.0 ℃孵育过夜,对照组载玻片滴加封闭液稀释的免疫前兔血清并以同样方式进行孵育,然后用PBS冲洗;避光条件下向载玻片均匀滴加用PBS稀释100倍的荧光二抗[羊抗兔二抗,Alexa488(GAR-Alexa488)],37 ℃恒温摇床避光孵育1 h后用PBS冲洗;避光条件下向载玻片均匀滴加DAPI染料,室温孵育;轻轻盖上盖玻片,立即于倒置荧光显微镜下观察。
2 结果与分析
2.1 SWP7基因序列的生物信息学分析
通过ExPASy在线软件对SWP7序列的基本特征进行分析,结果显示:SWP7由250个氨基酸组成,分子质量为27.8 kDa,等电点为5.25。SignalP 4.0预测SWP7的N端具有一个疏水峰,信号肽剪切位点位于第21~22个氨基酸之间(图2)。SWP7无GPI锚定位点和跨膜域(图3),通过ProtScale对SWP7疏水性进行分析发现,SWP7的前25个左右的氨基酸疏水性较高(图4),符合信号肽预测的疏水性峰位置。SWP7没有潜在的糖基化位点,但有多个磷酸化位点(图5),主要集中在苏氨酸(Thr)、丝氨酸(Ser)和酪氨酸(Tyr)上。另外,未在SWP7序列中预测到已知功能的结构域。

图2 SWP7的信号肽预测分析Fig.2The prediction analysis of signal peptide in SWP7
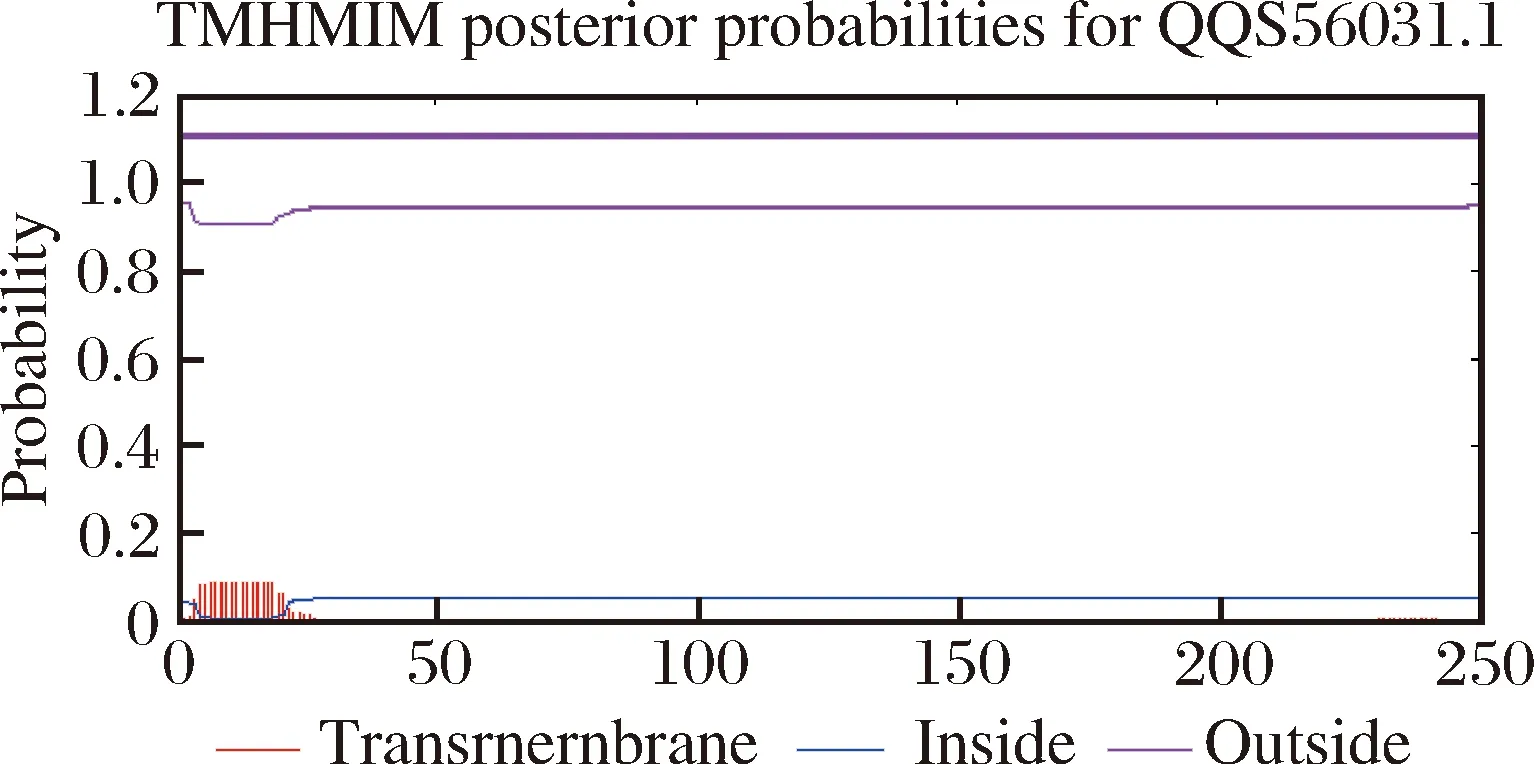
图3 SWP7的跨膜域预测分析Fig.3The prediction analysis of transmembrane domain in SWP7

图4 SWP7的疏水性预测分析Fig.4The hydrophobicity prediction analysis of SWP7
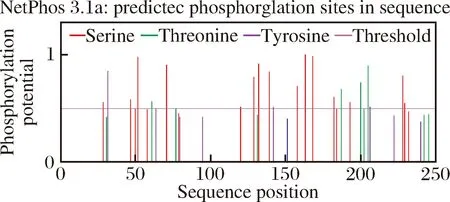
图5 SWP7的磷酸化位点预测分析Fig.5 The prediction analysis of phosphorylation site inSWP7
2.2 SWP7基因克隆
根据从基因组数据库下载的SWP7序列信息,基因合成该序列(689 bp)并连接到pET28a载体上,重组质粒双酶切验证构建pET28a-SWP7阳性克隆(图6-A)。将目的DNA 片段与载体连接,然后转入大肠杆菌,涂布培养后筛选阳性菌落进行PCR鉴定(图6-B)。结果显示扩增所得条带与目的条带长度一致。选取阳性单克隆菌落进行扩培,然后提取质粒,测序比对结果正确,表明已成功构建pET28a-SWP7原核表达载体。
2.3 SWP7蛋白的诱导表达与纯化
将上述阳性单克隆菌落于37 ℃恒温培养4 h至OD600约为0.6左右,加入终浓度为0.5 mmol/L的IPTG诱导pET28a-SWP7表达2 h后,收集蛋白并电泳检测表达情况(图7-A)。电泳结果显示:诱导的重组菌在28 kDa处有明显的高表达条带,而未诱导的重组菌没有该条带。放大培养后,用6xHis标签蛋白纯化试剂盒对SWP7蛋白进行纯化,结果如图7-B,可以看出,SWP7主要以包涵体形式存在于沉淀中,且纯度较高,上清液中目的蛋白很少,不易纯化,接着对SWP7包涵体进行复性和浓缩(图8),最终获得约8.0 mg蛋白。
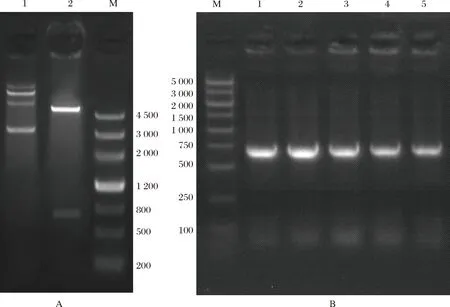
图6 虾肝肠胞虫SWP7原核表达载体的构建(A) pET28a-SWP7重组质粒双酶切验证。1:质粒DNA;2:pET28a-SWP7重组质粒的酶切产物;M:4500bp marker;(B) pET28a-SWP7重组质粒菌落PCR鉴定。M:5000bp marker;1-5:pET28a-SWP7菌检。Fig.6 The prokaryotic expression vector construction of EHP SWP7(A) The enzyme digestion analysis of pET28a-SWP7 recombinant plasmid. 1: Plasmid DNA; 2: The enzyme digested products of pET28a-SWP7 recombinant plasmid; M: 4500bp marker; (B) The bacterial colony PCR identification of pET28a-SWP7 recombinant plasmid. M: 5000bp marker; 1-5: The bacterial colony of pET28a-SWP7 recombinant plasmid.
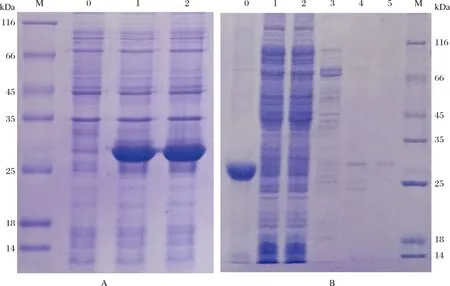
图7 pET28a-SWP7重组蛋白表达及纯化的SDS-PAGE检测(A)pET28a-SWP7诱导表达目的蛋白验证 M:蛋白质marker;0:pET28a-SWP7总菌体裂解液(未诱导);1-2:pET28a-SWP7总菌体裂解液(0.5 mmol/L IPTG 37 ℃,诱导2小时);(B) pET28a-SWP7融合蛋白的纯化 M:蛋白质marker;0:超声沉淀;1:超声上清;2:流出;3:Ni-IDA洗涤;4:Ni-IDA洗脱;5:残留。Fig.7The SDS-PAGE analysis of expression and purification of pET28a-SWP7 recombinant protein(A)The SDS-PAGE electropherogram of induced pET28a-SWP7 recombinant protein expressed in bacterium. M: Protein marker; 0: The uninduced pET28a-SWP7 recombinant protein expressed in total bacterial lysate; 1-2: The induced pET28a-SWP7 recombinant protein expressed in total bacterial lysate (0.5 mmol/L IPTG at 37 ℃ for 2 hours); (B) The purification of pET28a-SWP7 fusion protein; M: Protein marker; 0: The precipitation after ultrasonic treatment; 1: The supernatant after ultrasonic treatment; 2: The effluent; 3: The washing liquid of Ni-IDA; 4: The elution of Ni-IDA; 5: The residuum.
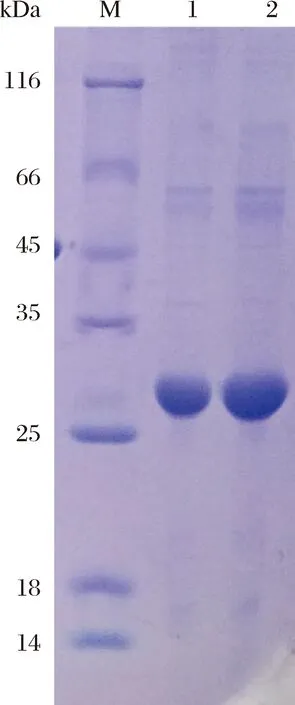
图8 pET28a-SWP7重组表达蛋白复性M:蛋白质 marker;1:pET28a-SWP7重组表达蛋白复性前;2:复性后浓缩。Fig.8 The renaturation of pET28a-SWP7 recombinant proteinM: Protein marker; 1: The induced pET28a-SWP7 recombinant protein before renaturation; 2: The induced pET28a-SWP7 recombinant protein after renaturation and concentration.
2.4 SWP7多克隆抗体的制备和Western Blot分析
用处理过的SWP7蛋白免疫新西兰白兔,8~12周后收集血液,通过间接ELISA法测定抗血清对SWP7的效价大于121 500(表1)。通过Weatern blotting检测抗血清(图9),结果表明:一抗稀释1 000倍,二抗稀释5 000倍时,抗体特异性很好,稀释1 000倍使用时仍可以检测出2.0 ng SWP7蛋白,灵敏度较高;并且SWP7蛋白Western Blot检测条带单一,与理论分子量相当,表明成功制备了SWP7多克隆抗体。
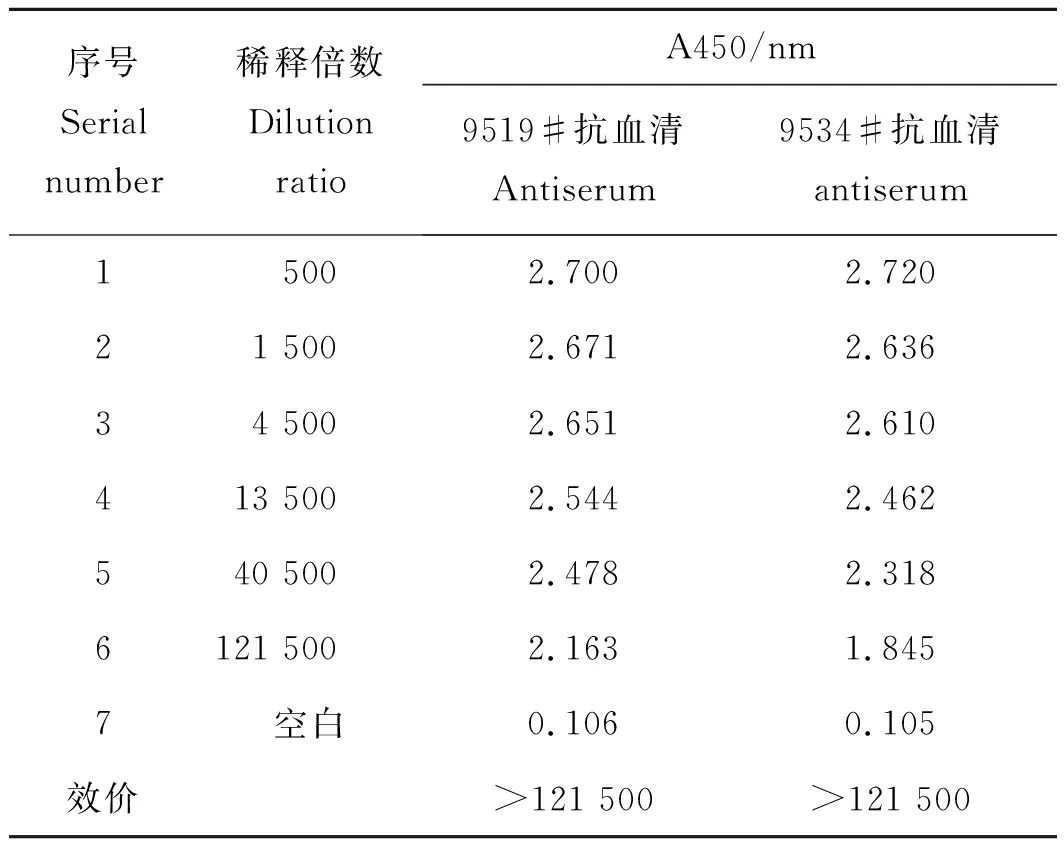
表1 抗血清间接Elisa结果Tab.1 Antiserum indirect Elisa results
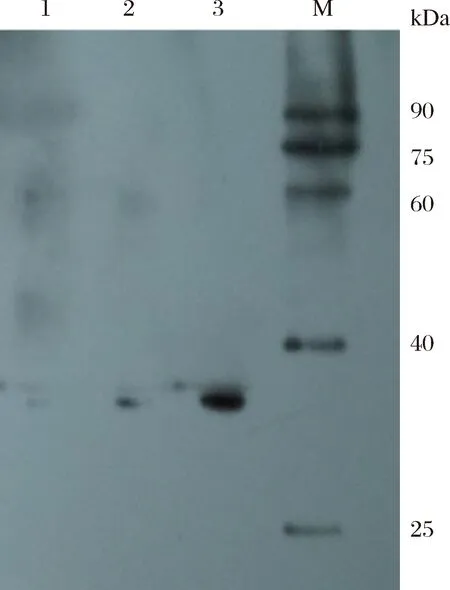
图9 SWP7多克隆抗体免疫印迹分析M:Western blot ECL marker;1:SWP7蛋白1.0 ng;2:SWP7蛋白2.0 ng;3:SWP7蛋白5.0 ng。Fig.9The western blot analysis of anti-SWP7 polyclonal antibodies in rabbitM: Western blot ECL marker; 1: SWP7 of 1.0 ng; 2: SWP7 of 2.0 ng; 3: SWP7 of 5.0 ng.
2.5 间接免疫荧光实验
利用间接免疫荧光实验对虾肝肠胞虫SWP7进行定位分析,结果如图10所示,在倒置荧光显微镜下,观察到SWP7抗体与孢壁识别发出绿色荧光,在明场下显示为椭圆形的成熟孢子,在蓝色荧光通道下,可观察到明亮的蓝色荧光,证明未萌发的EHP孢壁上有SWP7表达。
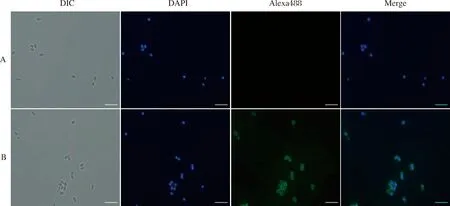
图10 间接免疫荧光实验(IFA)检测虾肝肠胞虫SWP7的定位A:对照组;B:实验组;DIC:明场;DAPI:示核,标记孢子位置;Alexa488:多克隆抗体标记SWP7在孢子中的定位信息;Merge:叠加图;比例尺=5.0 μm.Fig.10 The IFA analysis of the localization of SWP7A: Control group; B: Experimental group; DIC: Bright field; DAPI: Nucleus stained by DAPI, indicating spores location; Alexa488: The anti-SWP7 rabbit polyclonal antibody showed the SWP7 location in spores; Merge: Overlay chart. Scale=5.0 μm.
3 讨论
近年来,微孢子虫的孢壁研究成为国内外学者的热点,但是,由于微孢子虫个体微小,且孢壁结构独特,成分复杂,无形中加大了科研难度。目前已有的孢壁蛋白研究结果相对有限,为了深入研究并了解微孢子虫孢壁形成机制,有必要进一步探索微孢子虫孢壁中的其他蛋白成分,尤其是那些具有特殊结构的孢壁蛋白。本研究采用生物信息学方法对SWP7进行分析,结果显示该蛋白在N末端具有一段信号肽序列,但没有跨膜域和GPI锚定位点,推测该蛋白是一个分泌型蛋白。分泌蛋白合成于胞内,需分泌到胞外发挥作用,已有关于微孢子虫的研究表明,一些分泌蛋白是介导虫体入侵宿主细胞的重要因子[17-18],也可能发挥毒力作用,影响宿主免疫功能。在家蚕微孢子虫(Nosemabombycis)中,部分孢壁蛋白被证明为分泌蛋白,在粘附和侵染过程中发挥着关键作用[17,19]。SWP7未预测到潜在的糖基化位点,但有多个磷酸化位点。蛋白质翻译后会利用糖基化和磷酸化等修饰方式,促进蛋白质折叠的正确性及其构象的稳定性,调节和控制蛋白质的活力和多种生理功能,如信号转导、免疫应答、受体激活等[20]。
本研究制备了SWP7的兔多克隆抗体,这为后续研究该蛋白的功能奠定了基础。为了研究该蛋白在EHP中的表达情况,利用间接免疫荧光技术对其进行了定位。间接免疫荧光结果如图10所示,实验组中在被DAPI标记的成熟孢子周围有环状的绿色信号存在,而明场下呈空泡状的孢子未被DAPI染色,表明该孢子已经发芽,但在该孢子的外壳上仍可见环状的绿色荧光,进一步说明了SWP7确为EHP的孢壁蛋白,且孢子成熟时SWP7已被分泌到孢壁上。另外值得注意的是,已萌发的成熟EHP外壳上仍可获得较强的SWP7荧光信号,推测SWP7可能参与到了萌发过程中。孢壁是微孢子虫沟通外界环境的桥梁,可能参与了侵染时的粘附、信号转导等过程。Pattana等[21]鉴定了EHP的第一个孢壁蛋白(SWP1),在孢子内壁和外壁均有表达,其能够利用N端的3个肝素结合基序(HBMs),在感染过程中主动连接宿主细胞表面的肝素。家蚕微孢子虫中,SWP9作为一种支架蛋白,比SWP7更早地定位于孢子母细胞的孢壁。随后,SWP7分泌到孢壁并与SWP9相互作用,在黏附和感染中共同发挥作用[22]。兔脑炎微孢子虫有一个孢壁蛋白4(SWP4),定位于孢子内壁,其半胱氨酸含量较高,能够利用本身HBMs与宿主细胞膜表面的粘多糖GAG识别作用,参与并推动孢子与宿主细胞的粘附入侵过程[13,23]。成熟孢子的外壁含有大量的孢壁蛋白(SWPs),被认为是与环境和宿主细胞相互作用的第一层,也是最直接的一层[24]。同时,外壁中包含的SWPs是重要的功能蛋白,可能参与信号传递、粘附或酶促相互作用等重要过程[10,24]。
本研究中利用生物信息学方法对SWP7进行分析,结果显示SWP7为一种分泌蛋白,间接免疫荧光结果显示该蛋白定位于EHP孢壁,我们推测SWP7可能参与到虫体的萌发过程,研究结果对于明确EHP孢壁结构组成和进一步研究EHP的入侵机理具有重要意义。
猜你喜欢
环球时报(2022-09-20)2022-09-20
今日农业(2020年24期)2020-12-15
科学(2020年3期)2020-11-26
当代水产(2020年3期)2020-06-15
中国感染控制杂志(2015年7期)2015-12-13
小星星·阅读100分(高年级)(2015年11期)2015-11-28
科学大众(中学)(2015年9期)2015-10-12
小资CHIC!ELEGANCE(2015年15期)2015-09-01
少儿科学周刊·少年版(2015年3期)2015-07-07
医学研究杂志(2015年12期)2015-06-10
