
内蒙古阿拉善左旗乌兰塔塔尔最晚始新世-渐新世剖面的区域年代地层意义
2022-03-03 00:55JoonasWasiljeff张兆群
古脊椎动物学报(中英文) 2022年1期
Joonas Wasiljeff 张兆群
(1 芬兰赫尔辛基大学地球科学与地理学系 赫尔辛基 00014)
(2 中国科学院古脊椎动物与古人类研究所,中国科学院脊椎动物演化与人类起源重点实验室 北京 100044)
(3 中国科学院生物演化与环境卓越创新中心 北京 100044)
(4 中国科学院大学地球与行星科学学院 北京 100049)
1 Introduction
Transition from the Eocene to the Oligocene is a critical interval of the Paleogene period, and has been associated with drastic cooling and aridification events (Zachos et al., 2001; Dupont-Nivet et al., 2007; Hren et al., 2013), faunal and floral turnovers (Sun et al., 2014; Pound and Salzmann, 2017) and changes in seasonality (Eldrett et al., 2009).The dramatic faunal reorganization associated with the Eocene-Oligocene Transition (EOT) had different forms in North America (Prothero and Heaton, 1996), Europe (Hooker et al., 2004) and Asia (Meng and McKenna, 1998).The turnover in Asia was coined the “Mongolian Remodeling” which signified the replacement of perissodactyl dominant faunas adapted to forest habitats of the Eocene with rodent-lagomorph dominant faunas adapted to open grasslands of the Oligocene (Meng and McKenna, 1998; Kraatz and Geisler, 2010).The turnover has been argued to be both synchronous with the EOT and linked to a change in the physical environment (Kraatz and Geisler, 2010; Zhang et al., 2012; Sun et al., 2014).It has been suggested that aridification played a pivotal role in the overhaul (Meng and McKenna, 1998; Sun et al., 2014; Wasiljeff et al., 2020).However, a recent study from Bayanhot Basin, Nei Mongol (Inner Mongolia), provides evidence for a gradual or stepwise faunal turnover, linked to the “Mongolian Remodeling”, occurring already sometime in the latest Eocene (Wasiljeff et al., 2020).While the exact timing of this turnover remains unresolved in the region, it suggests the EOT related faunal modulation in Asia is more complex than previously understood (Wasiljeff et al., 2020), possibly driven by the long-term and stepwise Eocene aridification and affected by biogeographical factors.
Precise regional chronostratigraphic framework is, therefore, the foundation in forming a comprehensive understanding of these faunal events and their relationship to geologic history.The global chronostratigraphic units were mainly defined by marine deposits, e.g.the Eocene-Oligocene Boundary is underpinned by the GSSP at 33.89 Ma in the marine deposits of the Massignano section near Ancona, Italy (Premoli Silva and Jenkins 1993).However, the widely distributed and fossil rich Cenozoic terrestrial deposits in China have required extensive chronostratigraphic classification and correlation.Hence a transition from established land mammal ages to chronostratigraphic stages was adopted by NSC (National Commission of Stratigraphy) in 2002, leaving many stages not well defined without proper upper and lower boundaries.
The early work on the Asian Land Mammal Ages (ALMA) was based on the North American Land Mammal Ages (NALMA) and was pioneered by Romer (1966).This led to further studies in Asian Paleogene chronostratigraphy (Wang et al., 2012).Since China hosts numerous rich mammalian fossil sites relevant to the ALMA, a natural continuation was the interest in creating Chinese Paleogene mammal ages (e.g.Li and Ting, 1983; Tong et al., 1995; Wang, 1997a, b; Wang et al., 2007).Even though some advances have been made in the past decades, the establishment of Chinese Land Mammal Stages has remained difficult mainly due to two major reasons according to Wang et al.(2019): 1) the scarcity of continuous records hampers a reliable calibration and definition of the stages; 2) ambiguities in the subdivisions and correlation in the classic areas, such as Ulantatal, have not been resolved.Wang et al.(2007) divided the Paleogene of China into 11 stages with distinctive faunal features.Recently, this scheme was slightly modified by Wang et al.(2019).In this paper we present a suggestion of the regional stratotype of Ulantatalian and the lower boundary of Tabenbulukian stage based on updated data from the Ulantatal sequence.
2 Retrospection of Baiyinian, Ulantatalian and Tabenbulukian Stages
BaiyinianIn the latest revision of the Chinese Land Mammal Stages, Wang et al.(2019) suggested Baiyinian stage to correspond the Ergilian of Mongolia, which represents the latest Eocene in the ALMA (Speijer et al., 2020; Fig.1).In an earlier scheme of the Chinese Land Mammals Ages, Wang (1997b) introduced Ulangochuian and Houldjinian stages to cover the latest Eocene, whilst the following study (Wang et al., 2007) merged Houldjinian to Ulangochuian and used Ulangochuian to cover both.However, recent evidence from the Erlian Basin (Nei Mongol) suggests a corresponding fauna to the Ergilian of Mongolia which was found in the deposits between the Early Oligocene faunas and the Late Eocene Ulangochuian fauna.Therefore, the Baiyinian land mammal age was proposed equivalent to the Ergilian of ALMA (Wang et al., 2019).According to Wang et al.(2019) the Baiyinian mammals have been reported only from a few sites, with the stage boundaries remaining uncertain.Despite the uncertainties, the faunal assemblage discovered from the basal part of the Xianaogangdai Formation, Erden Obo section in the Erlian Basin, Nei Mongol, has been used as a criterion determining the base of the Baiyinian Age.As for the numerical age for the Xianaogangdai Formation, the basal part of the formation falls to the top of magnetochron C17n.1n, yielding an estimated age of 37.2 Ma for the lower boundary of the Baiyinian (Wang et al., 2019), based on a preliminary paleomagnetic study.
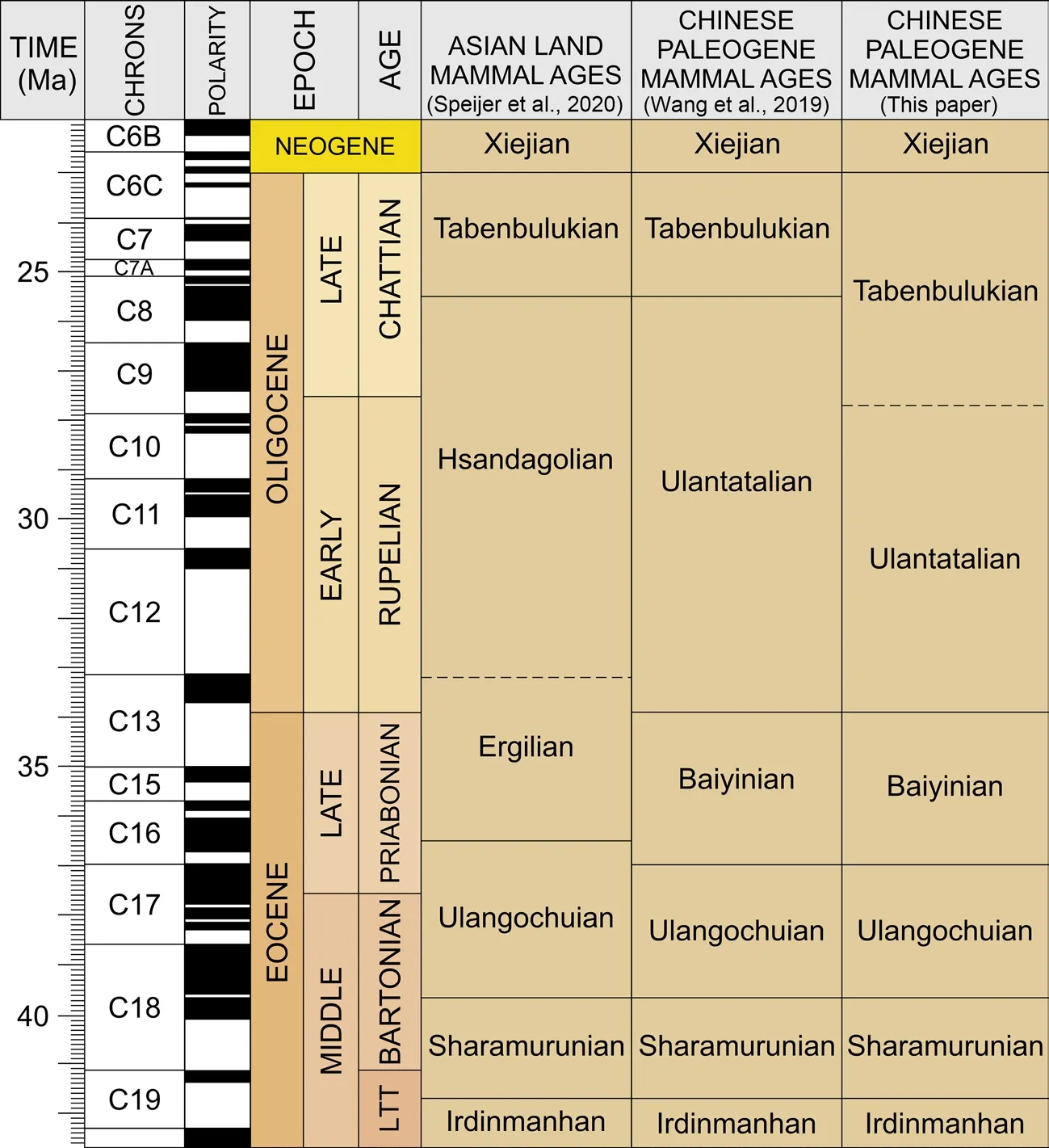
Fig.1 Chinese Paleogene Mammal Ages in relation to Asian Land Mammal Ages Abbreviation: LTT.Lutetian
Fossil mammals restricted to the Baiyinian stage include (c.f.Wang et al., 2019): artiodactylsEntelodon gobiensisandBothriodon chowi, perissodactylsSchizotherium avitum,Embolotherium andrewsi,Zaisanamynodon borisovi,Gigantamynodon giganteus,Urtinotherium intermedium,Parabrontops gobiensis,Ardynia praecox(mongoliensis),Proeggysodon qiuiandGuangnanodon, rodentsHeosminthus primiveris,H.nomogenesis,Sinosminthus inapertus,Allosminthus ernosandA.majusculus.
UlantatalianUlantatal fauna was first named by Huang (1982) for the mammalian fossils collected in the Ulantatal area without detailed stratigraphic records.Tong and Huang (Tertiary Research Group of Chinese National Petroleum Corporation, 1991) suggested the Ulantatalian Land Mammal Age based on this fauna.The nomenclature was then followed by Tong et al.(1995), who further merged the Kekeamuan, named by Wang (1992), into the Ulantatalian Age.These studies correlated the Ulantatalian stage with the latest Stampian/earliest Chattian of Europe.In the following study, the Ulantatalian fauna was considered to cover only the latest early Oligocene, and Wang (1997a) decided to use Hsandagolian (part of the ALMA; Fig.1) as the early Oligocene Land Mammal Age mirroring the proposition by Romer (1966).However, as Hsandagolian may in part overlap with the Tabenbulukian, Wang et al.(2007, 2019) recommended to use Ulantatalian as the early Oligocene land mammal age with its lower base corresponding to the formally fixed base of the Oligocene Series (base of the Rupelian stage), 33.89 Ma (Premoli Silva and Jenkins, 1993; Speijer et al., 2020).
Fossil mammals restricted to the Ulantatalian stage include (c.f.Wang et al., 2019): artiodactylsEumeryx culminisandPraetragulus gobiae, lagomorphsDesmatolagus pusillus,D.youngi, andOrdolagus teilhardi, hyaenodontHyaenodon neimongoliensis, erinaceomorphPalaeoscaptor acridens, rodentsEomys orientalis,Selenomys,Eucricetodon asiaticus,E.meridionalis,Bagacricetodon tongi,Plesiodipus wangae,Witenia yolua,Pseudocricetops matthewi,Anomoemys lohiculus,Litodonomys huangheensis,Euryodontomys exiguus,Coelodontomys asiaticus,Cyclomylus lohensis,Tsaganomys altaicus,Ageitonomys neimongolensis,Tataromys sigmodon,Karakoromys decessus,Paracricetops virgatoincisus,Cricetops dormitor,Propalaeocastor irtyshensis, andBohlinosminthus parvulus, primatesYunnanadapis folivorus,Y.imperator,Laomaki yunnanensis,Gatanthropus micros,Bahinia banyueae, andOligotarsius rarus.
TabenbulukianTabenbulukian Land Mammal Age is based on the Yandantu (Yindirte) fauna of the Tabenbuluk area, Gansu (Li and Ting, 1983; Wang et al., 2008), but the age has remained elusive.The fauna has been since recognized in the Paoniuquan Formation (Wang et al., 2003), however without any reliable dating, and in the lower part of the Xianshuihe Formation in the Lanzhou Basin, Gansu (Qiu et al., 1997), where paleomagnetic records do not confidently constrain the age of the fossil bearing horizons (Yue et al., 2001).Kraatz and Geisler (2010) suggested the Tabenbulukian to be significantly older than previously recognized according to their magnetostratigraphic study on some sections in the Valley of Lakes area, Tsagaan Nuur Basin, Mongolia.Tabenbulukian fossils first appeared from the base of C11r dated to late-early Oligocene (30.6 Ma).According to a recent work of Daxner-Höck et al.(2017) in the Valley of Lakes area, the boundary between Tabenbulukian and Hsandagolian (ALMA) was located in the middle of the Chron C8n.2n with an age of 25.5 Ma, which was then followed by Wang et al.(2019).
Fossil mammals restricted to the Tabenbulukian stage include (c.f.Wang et al., 2019): erinaceomorphsAmphechinus rectus, andA.minimus, lagomorphsSinolagomys major,S.kansuensis, andS.gracilis, rodentsSayimys obliquidens,Yindirtemys grangeri,Y.ambiguus,Y.deflexus,Plesiosminthus asiaecentralis,P.tangingoli,P.parvulus,Heterosminthus lanzhouensis,Tataromys parvus,T.plicidens,T.minor, andDistylomys qianlishanensis, perissodactylsDzungariotherium orgosense, andParaceratherium yagouense.
3 Geological setting
The Ulantatal sequence is a unique fossiliferous area near Bayanhaote, a major city in Alashan Zuoqi, Nei Mongol.The sequence totals over 100 m and can be divided into three sections from stratigraphically the lowermost to the highest: Kekeamu, Shangjing, and Ulantatal main section (Fig.2).The sequence is dominantly composed of laterally and vertically continuous fine-grained deposits, mainly originated from eolian dust deposition (Wasiljeff et al., 2020).The strata are gently inclined to SW and host numerous fossil localities, predominated by rich micromammalian faunas, well laid in stratigraphical superposition (Zhang et al., 2016; Wasiljeff et al., 2020).Currently the only comparable long sequence in the region of Eastern Asia can be found from the Valley of Lakes area, Mongolia.While Oligocene stratigraphy in the Valley of Lakes area is composed of multiple localities and sections, Ulantatal provides a geographically localized and stratigraphically robust continuity, which is often rare in terrestrial sequences.Ulantatal has been surveyed, excavated, and studied sporadically during the past decades (e.g.Huang, 1982; Wang and Wang, 1991; Vianey-Liaud et al., 2006; Gomes Rodrigues et al., 2014), but it was not until 2009, when intensive field research on the area begun (Zhang et al., 2016; Wasiljeff et al., 2020).
Based on earlier research in the area, the Ulantatal sequence was considered to be constrained to the Oligocene (e.g.Huang, 1982; Russell and Zhai, 1987; Wang, 1997a, b; Gomes Rodrigues et al., 2014).Wasiljeff et al.(2020) redefined the preliminary stratigraphical work of Zhang et al.(2016) by creating the first chronostratigraphic framework for the Ulantatal sequence, using paleomagnetic reversal stratigraphy and biostratigraphical correlation.Based on this work, the Ulantatal formation spans a time interval of ca.35 to 27 Ma, from the latest Eocene to late Oligocene.By interpolation, the Eocene-Oligocene boundary (EOB, 33.89 Ma) and therefore, also the base of Ulantatalian stagesensu stricto, can be placed at 15.9-meter level of the Ulantatal sequence.Moreover, this robust chronostratigraphic framework also allows a better understanding of other productive latest Eocene-late Oligocene localities in China through correlation to Ulantatal, such as Saint Jacques, Nei Mongol, or the small mammal bearing Erden Obo Section, Siziwangqi, Nei Mongol, which has been suggested to bracket the EOT (Li et al., 2016; 2017).
4 Bio- and chronostratigraphical features of Ulantatal in relation to Chinese regional Paleogene Stages
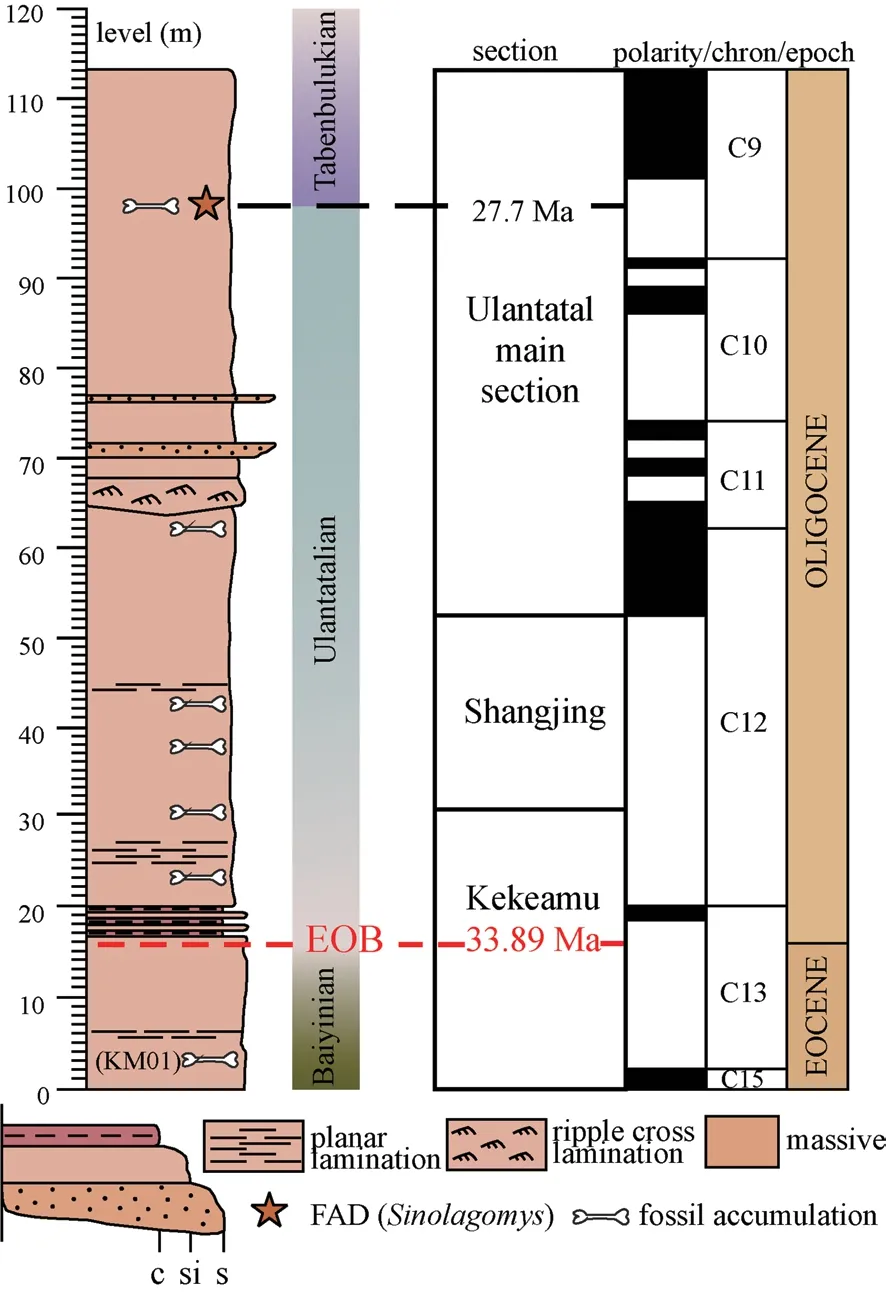
Fig.2 Simplified lithological column of the Ulantatal sequence with its sections and magnetic polarity column (based on Wasiljeff et al., 2020), and regional terrestrial mammal stages Focal fossil localities are indicated in the figure (bone symbol) in addition to First Appearance Datum (FAD) of the lagomorph taxon Sinolagomys (star symbol) Eocene-Oligocene Boundary (EOB) and the FAD (Sinolagomys) with their respective ages are also depicted
To meet the requirements for establishing regional chronostratigraphic units, long continuous sequences rich in fossils should be selected for defining the lower boundary stratotypes.The Ulantatal sequence, a hundred-meter-long continuous strata with profuse mammalian fossils in stratigraphic superposition (Fig.2; Zhang et al., 2016; Wasiljeff et al., 2020) provides a good candidate for defining the lower boundaries of Chinese Oligocene Stages.
Baiyinian and UlantatalianThe Kekeamu section covers the first ca.30 m of the Ulantatal sequence and hosts numerous fossil localities.The first fossil locality of Kekeamu section (KM01) is dated to 34.8 Ma (Fig.2; Wasiljeff et al., 2020), and thus is representative of the latest Eocene.Fossils reported in the locality include: rodentsProsciurussp.,Ageitonomys neimongoliensis,Karakoromyscf.K.decessus,Ardynomyssp.,Allosminthussp.nov.,Heosminthussp.,Parasminthussp.,Shamosminthussp.nov.?,Eucricetodonsp., andEocricetodonsp., erinaceomorphPalaeoscaptorsp.nov., lagomorphDesmatolaguscf.D.vetustus, perissodactylsSchitzotheriumcf.S.avitum, andArdyniacf.A.mongoliensis(Wang and Wang, 1991; Zhang et al., 2016; Wasiljeff et al., 2020).Of them, some are representative fossils of the Baiyinian stage, such asArdyniacf.A.mongoliensis,Allosminthus,Heosminthus, andEocricetodon.The rodents,Allosminthus,HeosminthusandEocricetodon, are shared in the genus level with more derived species with the Qujing fauna from Caijiachong Formation of Yunnan (Wasiljeff et al., 2020).
Based on our recent field investigations on fossil faunas and magnetostratigraphic age determinations it is evident that the fossil assemblage from the lowermost Kekeamu locality (KM01) appears to be transitional between the Ulantatalian and Baiyinian faunas.This either implies thatAllosminthus,Heosminthus, andEocricetodonare inappropriate indicators of the Baiyinian stage orAgeitonomys neimongoliensisandKarakoromys decessusshould not be used in determining the Ulantatalian stage.If the validity of the Baiyinian stage indicators is questioned, it urges extending the range of Ulantatalian in the Chinese Paleogene Regional Stages significantly down from the previous assignment to the base of the Rupelian in the upper part of Chron C13r (33.89 Ma).On the other hand, defining the lower boundaries of Land Mammalian Stages is constrained also e.g., by lithologic markers and paleomagnetic reversals in addition to the biological criteria (Qiu et al., 2013).Hence, the boundaries of the stages need not be consistent with faunal changes.Therefore, for applicability and convenience, the stage would have its lower boundary at the EOB (33.89 Ma) at 15.9 m level of the local section (Figs.2, 3A).
Furthermore, above the EOB the Kekeamu, Shangjing and Ulantatal main sections host rich fossil accumulations representative of the Ulantatalian stage (Gomes Rodrigues et al., 2014; Zhang et al., 2016; Wasiljeff et al., 2020).Fossil mammals restricted to the Ulantatalian stage from the Ulantatal sequence include: artiodactylsEumeryx culminisandHanhaicerus, carnivorePalaeogale sectoria, lagomorphsDesmatolagus gobiensis,D.pusillus, andOrdolagus teilhardi, hyaenodontHyaenodon neimongoliensis, rodentsSelenomys,Pseudocricetops matthewi,Anomoemys lohiculus,Euryodontomys exiguus,Coelodontomys asiaticus,Cyclomylus lohensis,Tataromys sigmodon,Bounomys,Karakoromys decessus,Cricetops dormitor,Propalaeocastor irtyshensis, andBohlinosminthus parvulus.
The profusion of typical Ulantatalian fauna above the EOB in robust temporal control, the continuous sequence (Wasiljeff et al., 2020) and the extensive research undertaken in Ulantatal over the past ten years, are all features that endorse the determination of a regional Unit Stratotype.The lowermost Kekeamu section of the Ulantatal sequence is composed of five largely overlapping subsections that are temporally and lithostratigraphically well constrained.Therefore, we propose Kekeamu as the Chinese regional Unit Stratotype of the Ulantatalian stage.Other sections, Shangjing and Ulantatal main section, in Ulantatal could be auxiliary unit stratotypes.This suggestion is further corroborated by the easy accessibility to the location through the main road from Bayanhaote city, which in turn is only a short distance from Ningxia Hui Autonomous Region’s capital Yinchuan which has an international airport.

Fig.3 Field images from the Ulantatal sequenceA.middle of Kekeamu section (top ca.23 m in the local section, interpolated as 32.97 Ma) with the Eocene-Oligocene Boundary (EOB, 33.89 Ma, 15.9 m local section) and sampling sites for paleomagnetic analyses visible; B.the uppermost Ulantatal main section (top ca.110 m in the composite sequence, overlain by Miocene deposits).The lowest occurrence of Tabenbulukian fauna and the FAD of Sinolagomys can be pinpointed to the lower half of the picture B (ca.27.7 Ma at 98 m level of the composite sequence) Pictures by courtesy of Anu Kaakinen
TabenbulukianWasiljeff et al.(2020) suggested the fossil assemblages from uppermost Ulantatal sequence, namely the topmost part of Ulantatal main section (Fig.3), to resemble the late Oligocene Yandantu fauna from Danghe area, Gansu (Wang et al., 2003, 2008) with the localities yielding typical Tabenbulukian fossils: rodentsTataromys parvus,Yindirtemys grangeri,Parasminthus tangingoli,Eucricetodon jilantaiensis, Tsaganomyidae indet., erinaceomorphPalaeoscaptor acridens, didymoconidaeDidymoconusindet., lagomorphsSinolagomyscf.S.kansuensis, Leporidae indet., and Ochotonidae gen.et sp.nov., and artiodactyl Ruminantia indet.etc.
While the corresponding Tabenbulukian fauna mark the beginning of letter zone C1 in Taatsiin Gol and Taatsiin Tsagaan Nuur region in the Valley of Lakes where the appearance of these fauna was restricted later than 25.6 Ma (Chron C8n.2n) (Daxner-Höck et al., 2017), the first appearance of fossils representative of the Tabenbulukian fauna at ca.98 m level in the composite Ulantatal sequence indicates the lower boundary of the Tabenbulukian could be shifted to an earlier age (Figs.1, 2).Currently, our fossil findings imply the lower boundary falls in Chron C9r and has an interpolated age of ~27.7 Ma, which is coincidently very close to the lower boundary of Chattian, dated to 27.82 Ma (Coccioni et al., 2018).
Hence, this fossil horizon can be marked as a lower boundary of Tabenbukian stage.In addition to the faunal similarity to Tabenbulukian fauna in Gansu, extensive exploration and excavations during our nine field seasons confirmed the First Apperanance Datum (FAD) of the lagomorph taxonSinolagomysfrom this fossil-rich horizon.As the earliest crown group of Ochotonidae,Sinolagomyshas distinct tooth morphology from earlier stem groups, such asDesmatolagus, with rootless teeth, hypostria in upper molars etc.Its widely geographic distribution from Mongolia, Kazakhstan, and northern China also facilitates its biostratigraphic significance.
5 Conclusion
The Ulantatal sequence is till now the most representative record for the Ulantatalian stage, and Kekeamu section can serve as a regional lower boundary unit stratotype section of the stage, while other Ulantatal sections could be auxiliary unit stratotypes.This is supported by the robust temporal framework of Ulantatal sequence, the abundance of fossil fauna characteristic to the Ulantatalian stage, and by the easy accessibility to the locality.The lower boundary of the stage should remain at 33.89 Ma based on bio- and chronostratigraphic results in Ulantatal (Wasiljeff et al., 2020).Moreover, because the Tabenbulukian fossil bearing horizons in Ulantatal are dated to ca.27.7 Ma, we propose the lower boundary of Tabenbulukian stage to be reassigned at Chron C9r, with the lowest occurrence ofSinolagomysas the marker horizon.These features further promote Ulantatal as an important component of the Paleogene of China, which can be broadly utilized in correlation of terrestrial faunas in Asia.
AcknowledgementsThe authors would like to thank the colleagues who have participated in the still ongoing field work in Nei Mongol and acknowledge fruitful discussions with Anu Kaakinen.We would also like to appreciate the insightful comments of the two reviewers Bai Bin and Li Qian.The study was supported by Academy of Finland (Grant No.316799 to Anu Kaakinen) and CAS (XDB26000000, and special fund for fossil excavation and preparation), NSFC (41472003), GeoDoc (JW), and Waldemar von Frenckell’s Foundation (JW).
猜你喜欢
廉政瞭望(2022年11期)2022-07-28
河北地质大学学报(2021年5期)2021-11-08
房地产导刊(2021年8期)2021-10-13
红领巾·萌芽(2020年5期)2020-06-03
青年歌声(2019年2期)2019-02-21
——评乌兰杰的《蒙古族音乐史》
人民音乐(2018年8期)2018-08-21
草原歌声(2018年1期)2018-05-07
时尚北京(2016年6期)2016-05-14
大众考古(2015年5期)2015-06-26
